Transcriptomic Analysis of Streptococcus suis in Response to Ferrous Iron and Cobalt Toxicity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. RNA Extraction
2.3. cDNA Library Preparation and Sequencing
2.4. RNA Sequencing Data Analysis
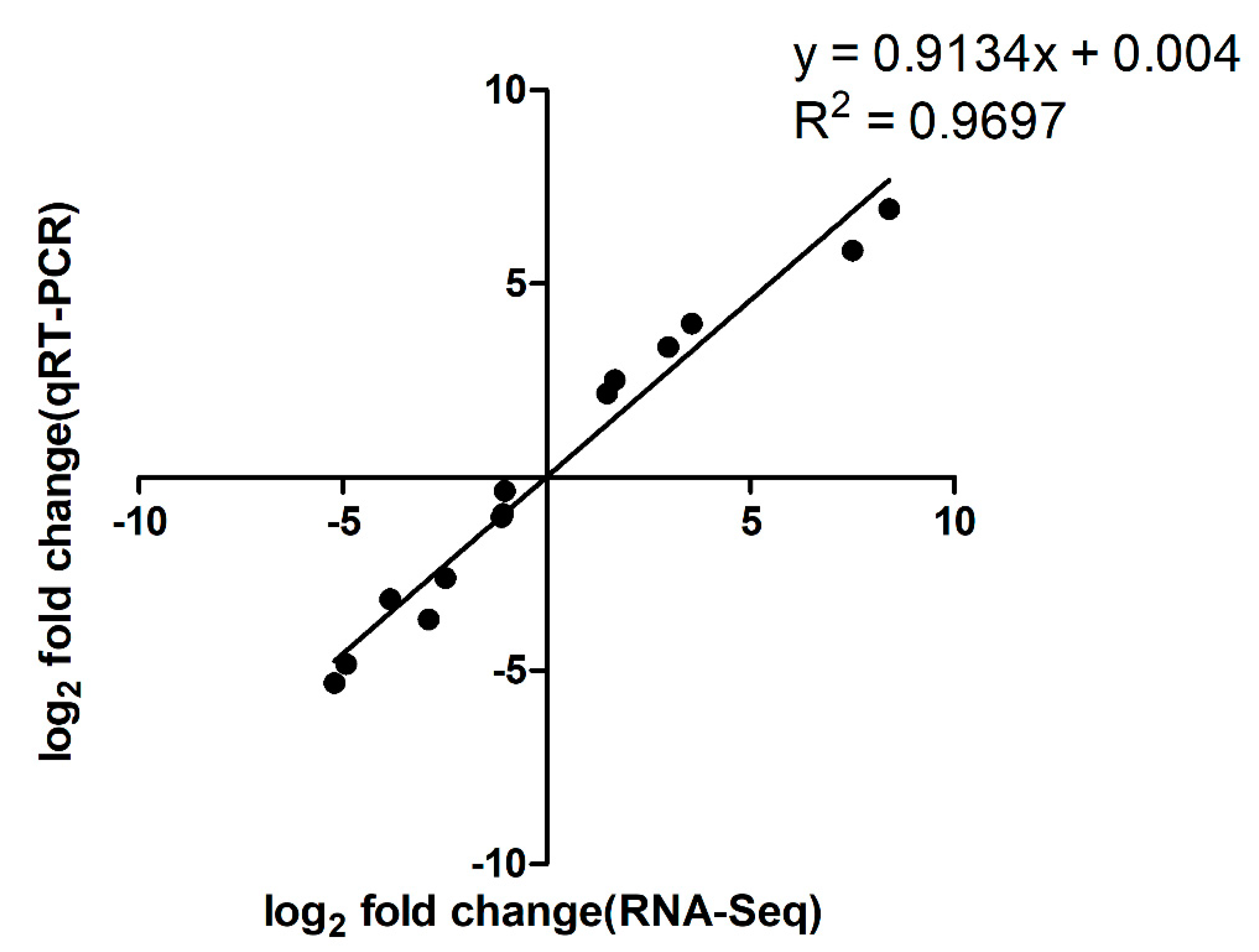
2.5. Gene Expression Analysis by qRT-PCR
3. Results
3.1. RNA Sequencing Information
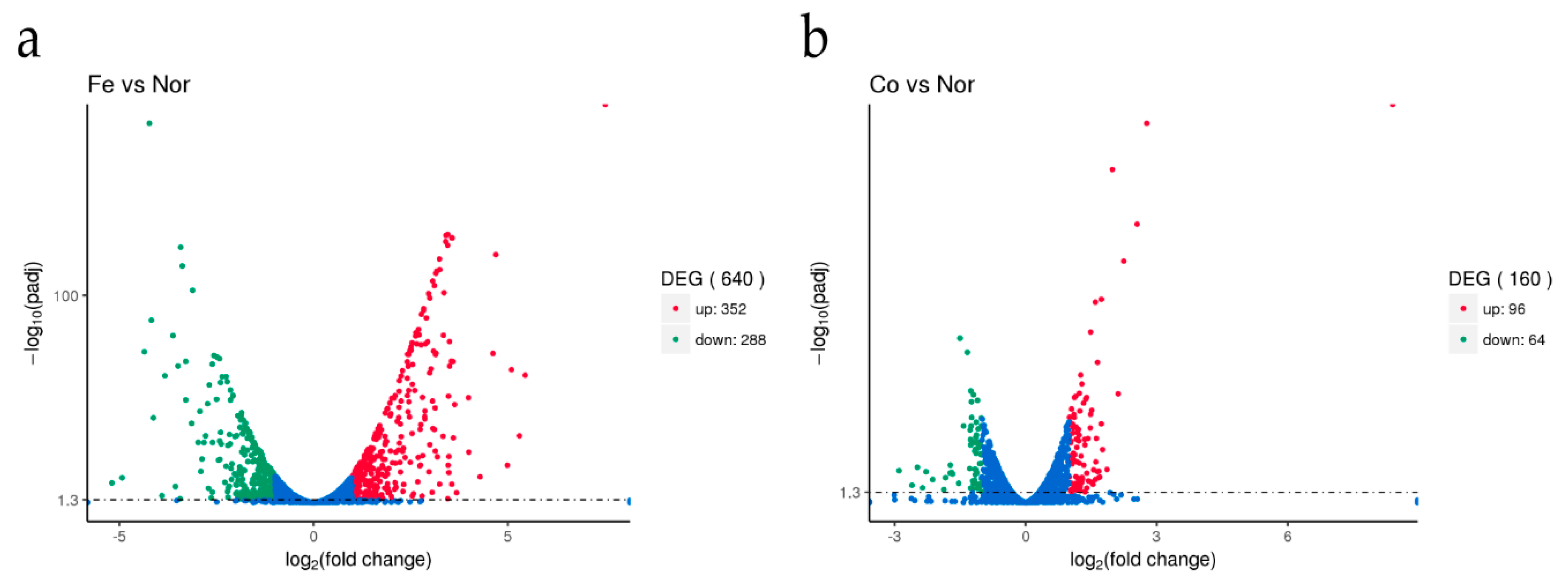
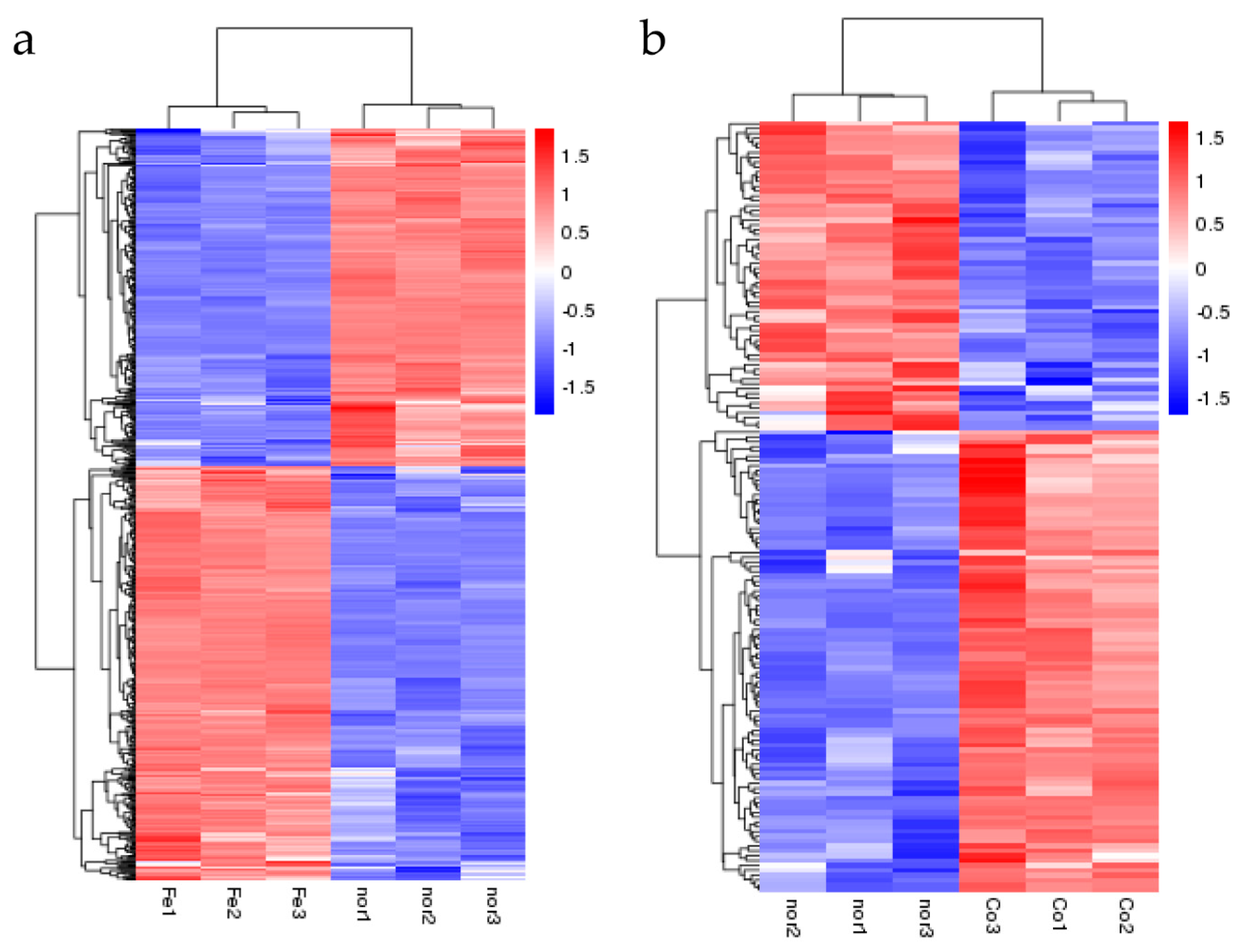
3.2. Changes in S. suis Transcriptome in Response to Ferrous Iron and Cobalt Toxicity
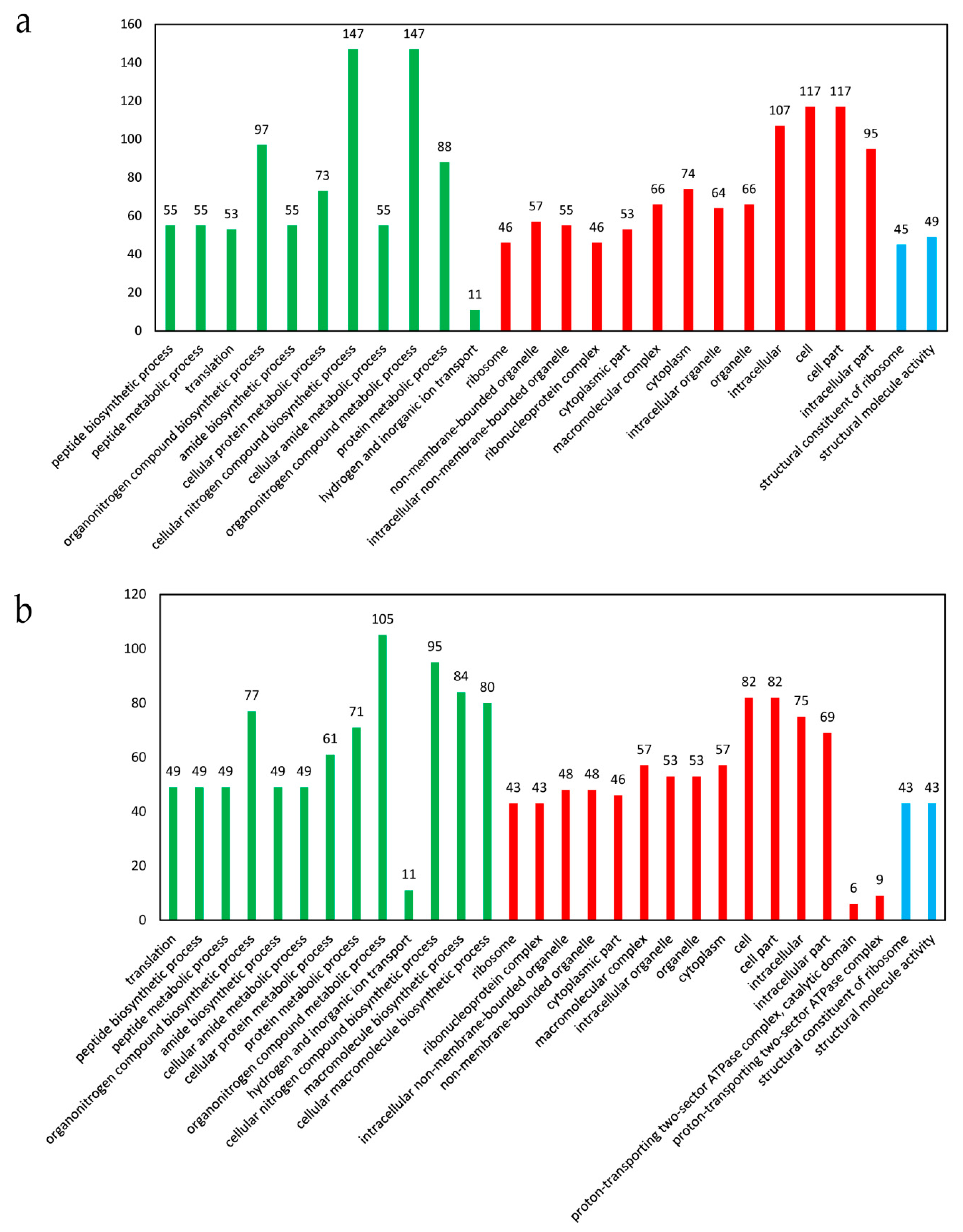
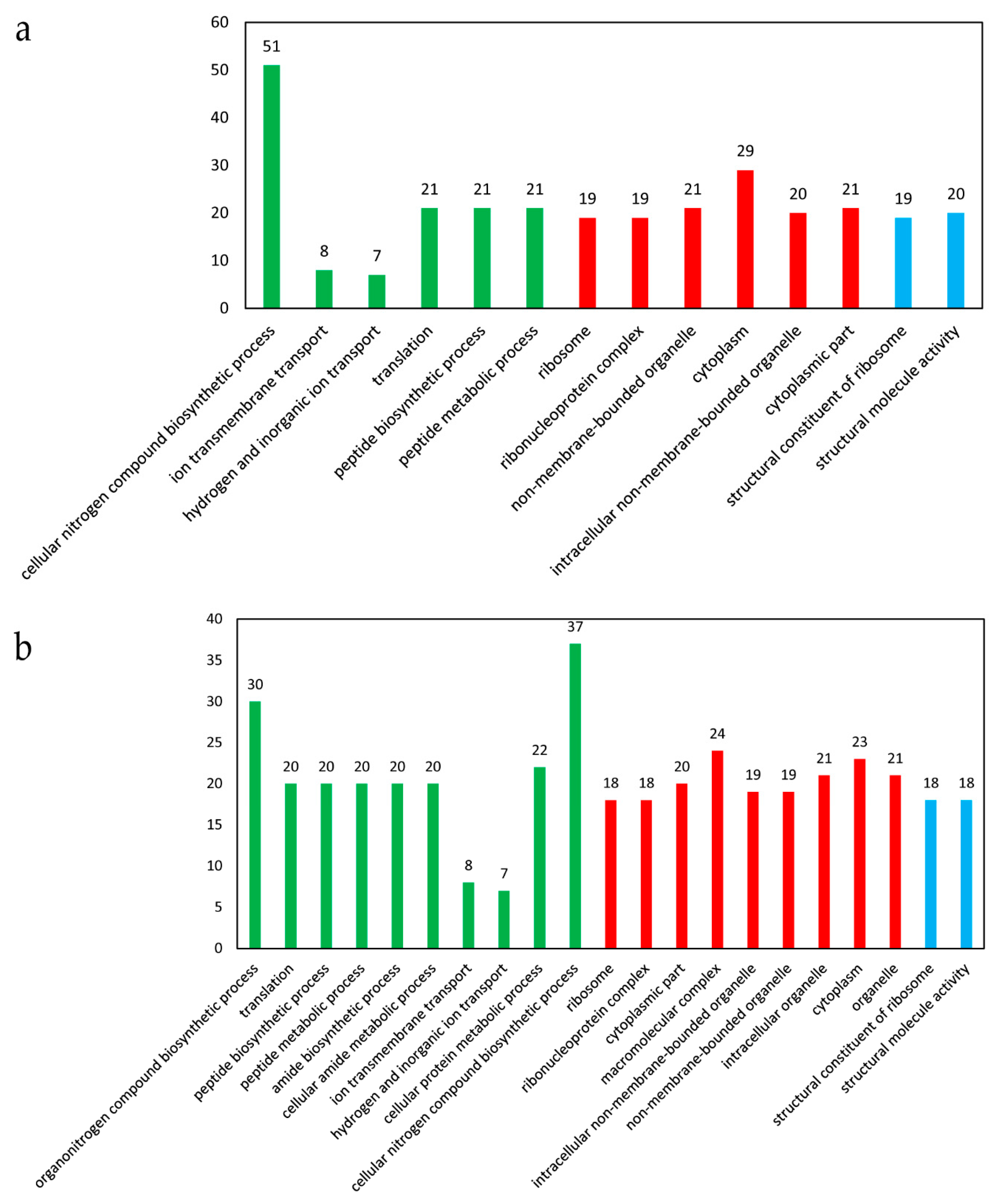
3.3. Gene Ontology (GO) Enrichment Analysis of the DEGs
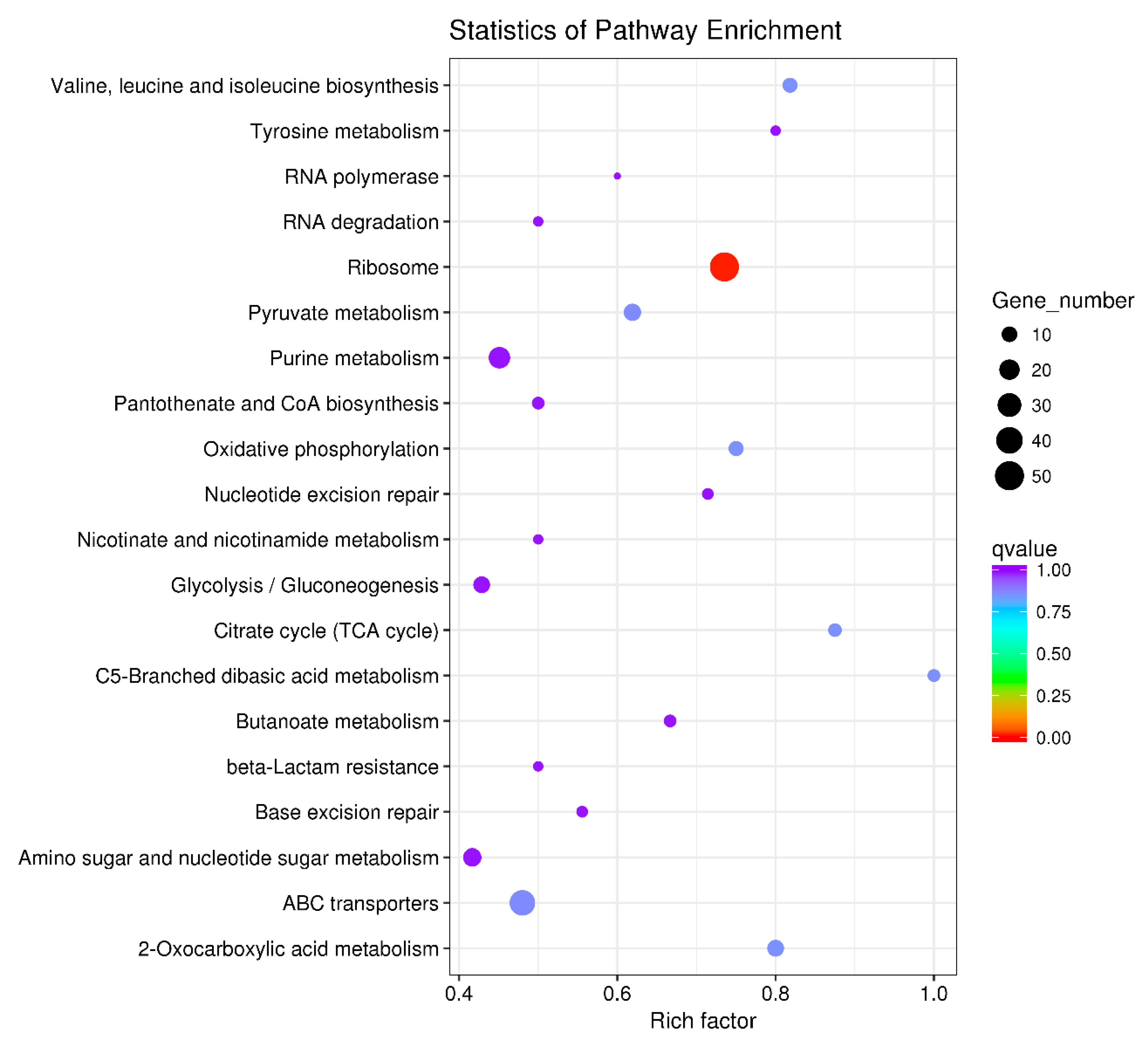
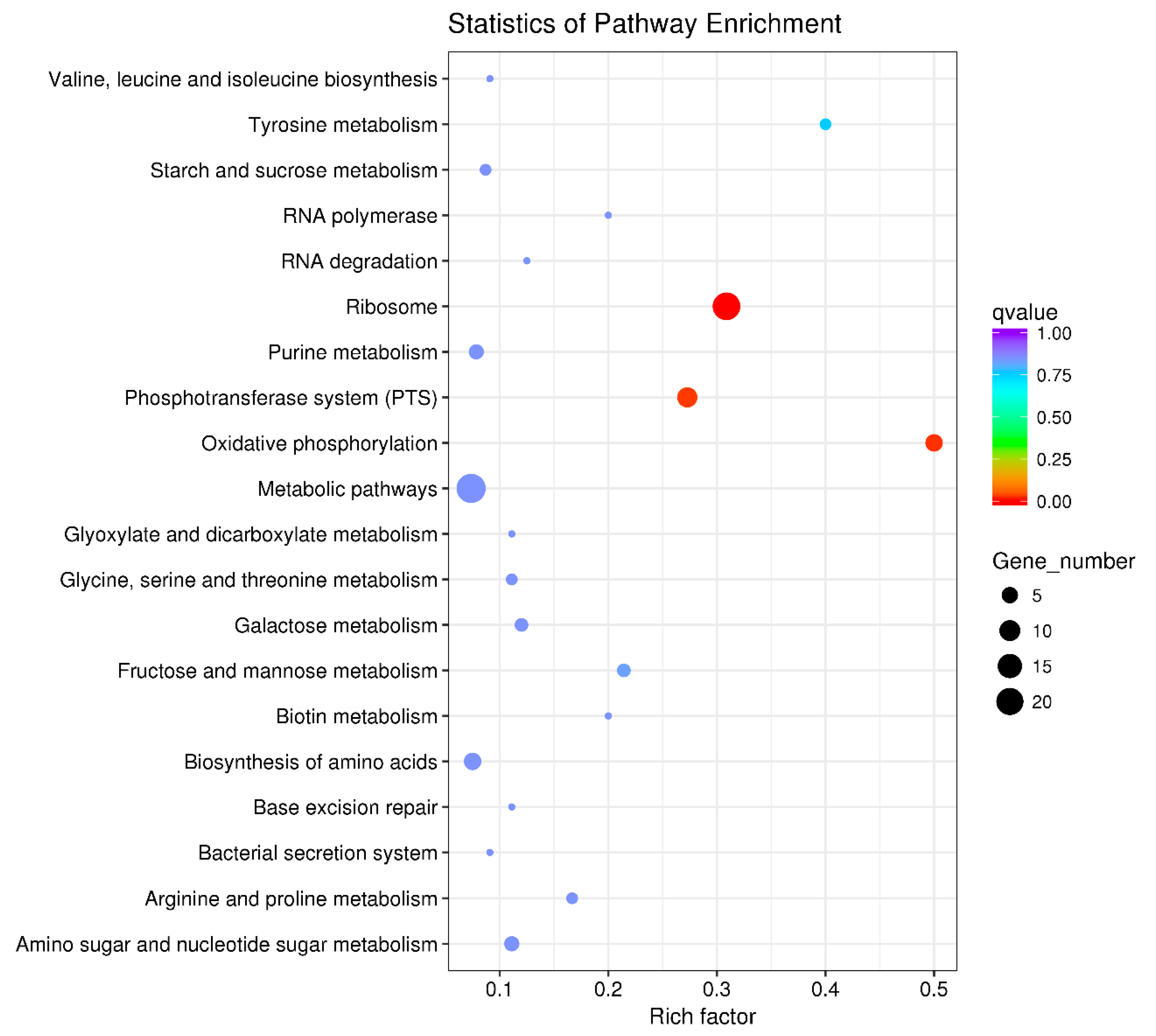
3.4. KEGG (the Kyoto Encyclopedia of Genes and Genomes) Pathway Enrichment Analysis of the DEGs
3.5. Several Genes Involved in Oxidative Stress Tolerance Were Significantly Down-Regulated in the Presence of Ferrous Iron
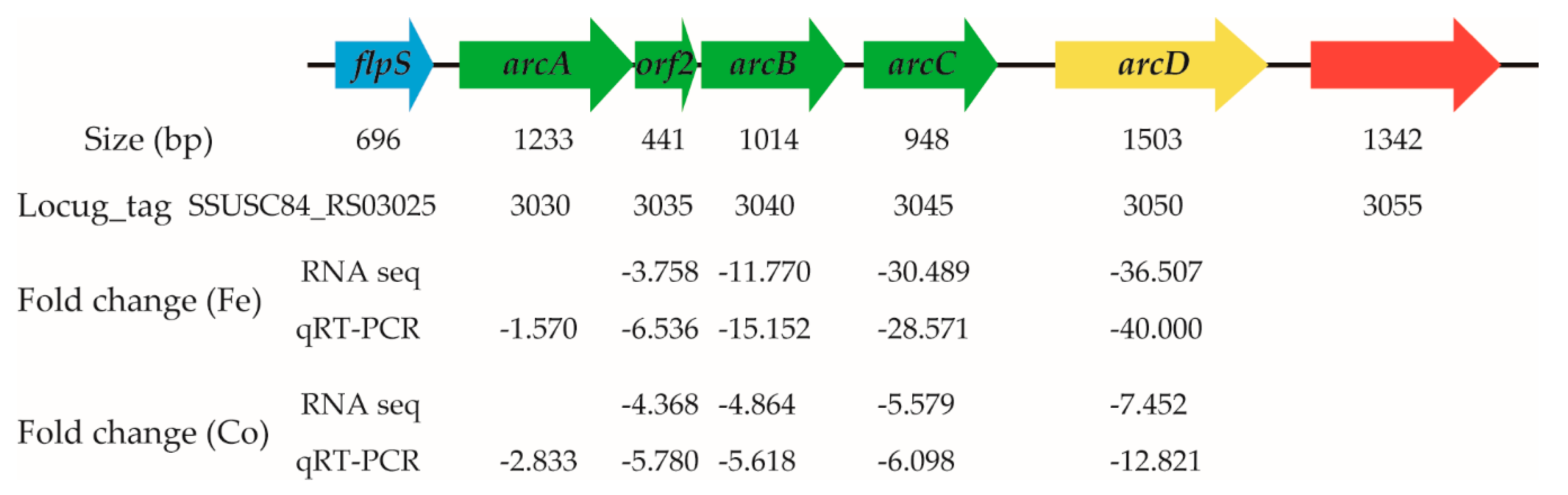
3.6. Treatment with Ferrous Iron and Cobalt Repressed Expression of Several Genes in the Arginine Deiminase System
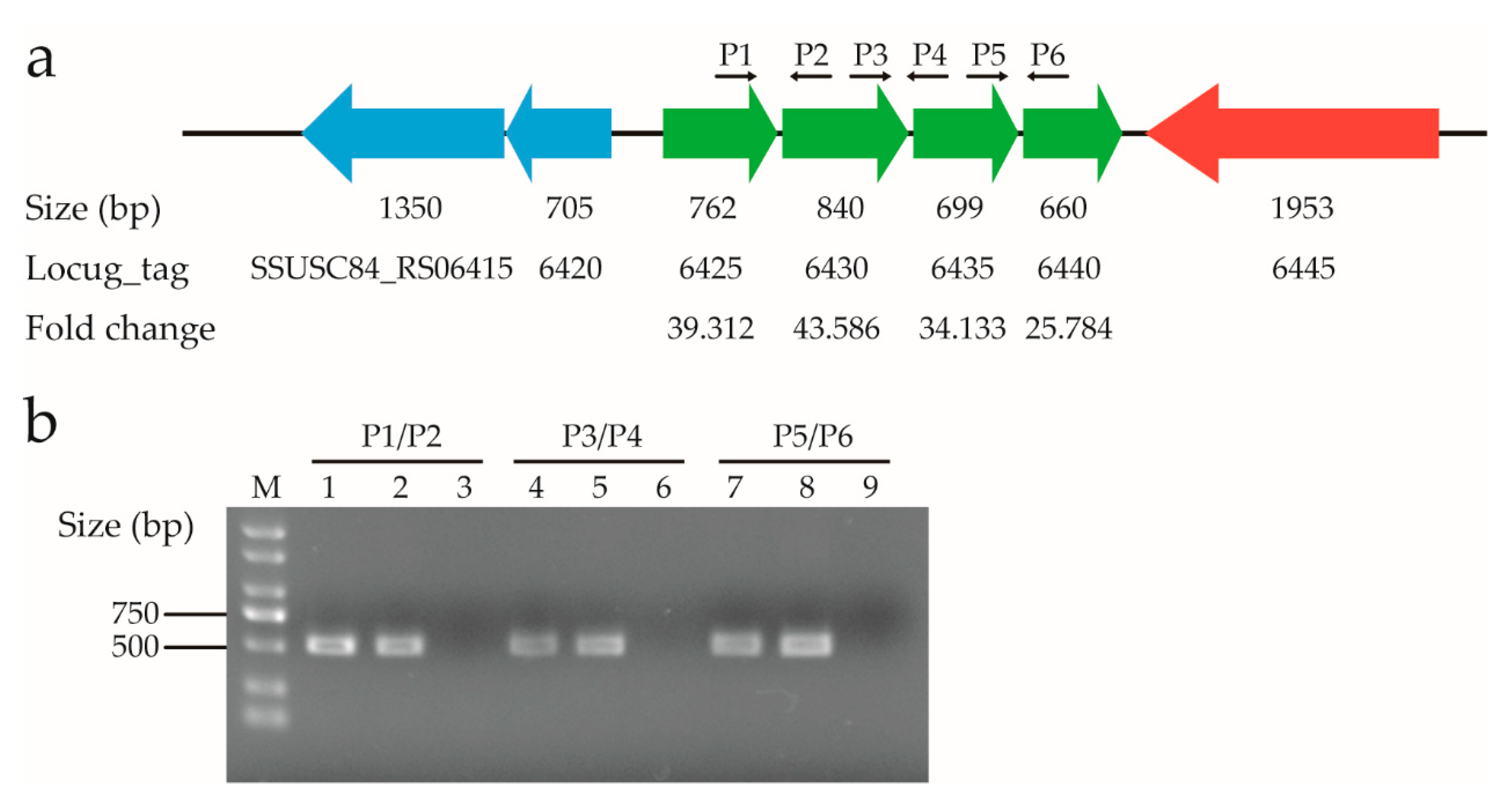
3.7. The Genes in an Amino Acid ABC Transporter Operon Were Significantly Up-Regulated in the Presence of Ferrous Iron
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Segura, M.; Zheng, H.; de Greeff, A.; Gao, G.F.; Grenier, D.; Jiang, Y.Q.; Lu, C.P.; Maskell, D.; Oishi, K.; Okura, M.; et al. Latest developments on Streptococcus suis: An emerging zoonotic pathogen: Part 1. Future Microbiol. 2014, 9, 441–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: Fighting against nonspecific defenses. FEBS Lett. 2016, 590, 3772–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatrongjit, R.; Fittipaldi, N.; Gottschalk, M.; Kerdsin, A. Tools for Molecular Epidemiology of Streptococcus suis. Pathogens 2020, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes. Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef]
- Agoston, Z.; Terhes, G.; Hannauer, P.; Gajdacs, M.; Urban, E. Fatal case of bacteremia caused by Streptococcus suis in a splenectomized man and a review of the European literature. Acta Microbiol. Immunol. Hung. 2020, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Guo, J.; Cheng, C.; Gu, B. Human infection caused by Streptococcus suis serotype 2 in China: Report of two cases and epidemic distribution based on sequence type. BMC Infect. Dis. 2020, 20, 223. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Lee, M.H.; Kim, Y.S.; Choi, J.K.; Yoo, J.H. A Case of Life-threating Streptococcus suis infection presented as septic shock and multiple abscesses. Infect. Chemother. 2018, 50, 274–279. [Google Scholar] [CrossRef]
- Yanase, T.; Morii, D.; Kamio, S.; Nishimura, A.; Fukao, E.; Inose, Y.; Honma, Y.; Kitahara, N.; Yokozawa, T.; Chang, B.; et al. The first report of human meningitis and pyogenic ventriculitis caused by Streptococcus suis: A case report. J. Infect. Chemother. 2018, 24, 669–673. [Google Scholar] [CrossRef]
- Zhang, B.; Ku, X.; Yu, X.; Sun, Q.; Wu, H.; Chen, F.; Zhang, X.; Guo, L.; Tang, X.; He, Q. Prevalence and antimicrobial susceptibilities of bacterial pathogens in Chinese pig farms from 2013 to 2017. Sci. Rep. 2019, 9, 9908. [Google Scholar] [CrossRef] [PubMed]
- Begg, S.L. The role of metal ions in the virulence and viability of bacterial pathogens. Biochem. Soc. Trans. 2019, 47, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.D.; Bobrov, A.G.; Fetherston, J.D. The role of transition metal transporters for iron, zinc, manganese, and copper in the pathogenesis of Yersinia pestis. Metallomics 2015, 7, 965–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honsa, E.S.; Johnson, M.D.L.; Rosch, J.W. The roles of transition metals in the physiology and pathogenesis of Streptococcus pneumoniae. Front. Cell Infect. Microbiol. 2013, 3, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.G.; Ong, C.Y.; Walker, M.J.; Djoko, K.Y.; McEwan, A.G. Transition metal homeostasis in Streptococcus pyogenes and Streptococcus pneumoniae. Adv. Microb. Physiol. 2017, 70, 123–191. [Google Scholar]
- Turner, A.G.; Ong, C.L.Y.; Gillen, C.M.; Davies, M.R.; West, N.P.; McEwan, A.G.; Walker, M.J. Manganese homeostasis in Group A Streptococcus is critical for resistance to oxidative stress and virulence. mBio 2015, 6, e00278-15. [Google Scholar] [CrossRef] [Green Version]
- Grunenwald, C.M.; Choby, J.E.; Juttukonda, L.J.; Beavers, W.N.; Weiss, A.; Torres, V.J.; Skaar, E.P. Manganese detoxification by MntE is critical for resistance to oxidative stress and virulence of Staphylococcus aureus. mBio 2019, 10, e02915-18. [Google Scholar] [CrossRef] [Green Version]
- Wolschendorf, F.; Ackart, D.; Shrestha, T.B.; Hascall-Dove, L.; Nolan, S.; Lamichhane, G.; Wang, Y.; Bossmann, S.H.; Basaraba, R.J.; Niederweis, M. Copper resistance is essential for virulence of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2011, 108, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.J.; Li, M.; Zhang, H.M.; Zheng, B.W.; Han, H.M.; Wang, C.J.; Yan, J.H.; Tang, J.Q.; Gao, G.F. Functional definition and global regulation of Zur, a zinc uptake regulator in a Streptococcus suis serotype 2 strain causing streptococcal toxic shock syndrome. J. Bacteriol. 2008, 190, 7567–7578. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.L.; Zheng, C.K.; Cao, M.M.; Zeng, T.; Zhao, X.G.; Shi, G.L.; Chen, H.C.; Bei, W.C. The manganese efflux system MntE contributes to the virulence of Streptococcus suis serotype 2. Microb. Pathog. 2017, 110, 23–30. [Google Scholar] [CrossRef]
- Zheng, C.K.; Jia, M.D.; Lu, T.Y.; Gao, M.M.; Li, L.Z. CopA protects Streptococcus suis against copper toxicity. Int. J. Mol. Sci. 2019, 20, 2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vats, N.; Lee, S.F. Characterization of a copper-transport operon, copYAZ, from Streptococcus mutans. Microbiology 2001, 147, 653–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrakul, K.; Loo, C.Y.; Hughes, C.V.; Ganeshkumar, N. Role of a Streptococcus gordonii copper-transport operon, copYAZ, in biofilm detachment. Oral Microbiol. Immunol. 2004, 19, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Shafeeq, S.; Yesilkaya, H.; Kloosterman, T.G.; Narayanan, G.; Wandel, M.; Andrew, P.W.; Kuipers, O.P.; Morrissey, J.A. The cop operon is required for copper homeostasis and contributes to virulence in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 1255–1270. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.K.; Jia, M.D.; Gao, M.M.; Lu, T.Y.; Li, L.Z.; Zhou, P.P. PmtA functions as a ferrous iron and cobalt efflux pump in Streptococcus suis. Emerg. Microbes Infect. 2019, 8, 1254–1264. [Google Scholar] [CrossRef] [Green Version]
- Teng, L.; Dong, X.; Zhou, Y.; Li, Z.; Deng, L.; Chen, H.; Wang, X.; Li, J. Draft genome sequence of hypervirulent and vaccine candidate Streptococcus suis strain SC19. Genome Announc. 2017, 5, e01484-16. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Mao, X.Z.; Huang, J.J.; Ding, Y.; Wu, J.M.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L.P. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(−ΔΔC) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Svec, D.; Tichopad, A.; Novosadova, V.; Pfaffl, M.W.; Kubista, M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Ren, S.; Xu, J.; Zhao, X.; Shi, G.; Wu, J.; Li, J.; Chen, H.; Bei, W. Contribution of NADH oxidase to oxidative stress tolerance and virulence of Streptococcus suis serotype 2. Virulence 2017, 8, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Hajaj, B.; Yesilkaya, H.; Benisty, R.; David, M.; Andrew, P.W.; Porat, N. Thiol peroxidase is an important component of Streptococcus pneumoniae in oxygenated environments. Infect. Immun. 2012, 80, 4333–4343. [Google Scholar] [CrossRef] [Green Version]
- Hajaj, B.; Yesilkaya, H.; Shafeeq, S.; Zhi, X.Y.; Benisty, R.; Tchalah, S.; Kuipers, O.P.; Porat, N. codY Regulates Thiol peroxidase expression as part of the Pneumococcal defense mechanism against H2O2 stress. Front. Cell Infect. Microbiol. 2017, 7, 210. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.L.; Zhang, X.Y.; Wu, W.; Lu, Z.Y.; Fang, W.H. Inactivation of the sodA gene of Streptococcus suis type 2 encoding superoxide dismutase leads to reduced virulence to mice. Vet. Microbiol. 2012, 158, 360–366. [Google Scholar] [CrossRef]
- Fang, L.H.; Shen, H.X.; Tang, Y.L.; Fang, W.H. Superoxide dismutase of Streptococcus suis serotype 2 plays a role in anti-autophagic response by scavenging reactive oxygen species in infected macrophages. Vet. Microbiol. 2015, 176, 328–336. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Wu, F.L.; Pang, H.Y.; Tang, J.F.; Cai, S.H.; Jian, J.C. Superoxide dismutase B (sodB), an important virulence factor of Vibrio alginolyticus, contributes to antioxidative stress and its potential application for live attenuated vaccine. Fish. Shellfish Immunol. 2019, 89, 354–360. [Google Scholar] [CrossRef]
- Heindorf, M.; Kadari, M.; Heider, C.; Skiebe, E.; Wilharm, G. Impact of Acinetobacter baumannii superoxide dismutase on motility, virulence, oxidative stress resistance and susceptibility to antibiotics. PLoS ONE 2014, 9, e101033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Wang, Y.; Ni, Y.; Zhou, J.; Han, L.; Yu, Z.; Mao, A.; Wang, D.; Fan, H.; He, K. The Redox-Sensing Regulator Rex contributes to the virulence and oxidative stress Response of Streptococcus suis serotype 2. Front. Cell Infect. Microbiol. 2018, 8, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Yang, X.; Zhang, P.; Ma, Z.; Lin, H.; Fan, H. The arginine deiminase system facilitates environmental adaptability of Streptococcus equi ssp. zooepidemicus through pH adjustment. Res. Microbiol. 2016, 167, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hitzmann, A.; Bergmann, S.; Rohde, M.; Chhatwal, G.S.; Fulde, M. Identification and characterization of the arginine deiminase system of Streptococcus canis. Vet. Microbiol. 2013, 162, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Gruening, P.; Fulde, M.; Valentin-Weigand, P.; Goethe, R. Structure, regulation, and putative function of the arginine deiminase system of Streptococcus suis. J. Bacteriol. 2006, 188, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Schulz, C.; Gierok, P.; Petruschka, L.; Lalk, M.; Mader, U.; Hammerschmidt, S. Regulation of the arginine deiminase system by ArgR2 interferes with arginine metabolism and fitness of Streptococcus pneumoniae. mBio 2014, 5, e01858-14. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.G.; Ong, C.L.Y.; Djoko, K.Y.; West, N.P.; Davies, M.R.; McEwan, A.G.; Walker, M.J. The PerR-regulated P1B-4-Type ATPase (PmtA) acts as a ferrous iron efflux pump in Streptococcus pyogenes. Infect. Immun. 2017, 85, e00140-17. [Google Scholar] [CrossRef] [Green Version]
- VanderWal, A.R.; Makthal, N.; Pinochet-Barros, A.; Helmann, J.D.; Olsen, R.J.; Kumaraswami, M. Iron efflux by PmtA is critical for oxidative stress resistance and contributes significantly to Group A Streptococcus virulence. Infect. Immun. 2017, 85, e00091-17. [Google Scholar] [CrossRef] [Green Version]
- Pi, H.; Helmann, J.D. Ferrous iron efflux systems in bacteria. Metallomics 2017, 9, 840–851. [Google Scholar] [CrossRef]
- Li, W.; Liu, L.; Chen, H.; Zhou, R. Identification of Streptococcus suis genes preferentially expressed under iron starvation by selective capture of transcribed sequences. FEMS Microbiol. Lett. 2009, 292, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Lopez, G.; Latorre, M.; Reyes-Jara, A.; Cambiazo, V.; Gonzalez, M. Transcriptomic response of Enterococcus faecalis to iron excess. Biometals 2012, 25, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.M.; Diaz-Mejia, J.J.; Maldonado-Rodriguez, R.; Amabile-Cuevas, C.F. Differential activities of the SoxR protein of Escherichia coli: SoxS is not required for gene activation under iron deprivation. FEMS Microbiol. Lett. 2001, 201, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Masse, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwugo, C.C.; Gaddy, J.A.; Zimbler, D.L.; Actis, L.A. Deciphering the iron response in Acinetobacter baumannii: A proteomics approach. J. Proteom. 2011, 74, 44–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michta, E.; Ding, W.; Zhu, S.; Blin, K.; Ruan, H.; Wang, R.; Wohlleben, W.; Mast, Y. Proteomic approach to reveal the regulatory function of aconitase AcnA in oxidative stress response in the antibiotic producer Streptomyces viridochromogenes Tu494. PLoS ONE 2014, 9, e87905. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.M.; Coenye, T. Low iron-induced small RNA BrrF regulates central metabolism and oxidative stress responses in Burkholderia cenocepacia. PLoS ONE 2020, 15, e0236405. [Google Scholar] [CrossRef]
- Stirnberg, M.; Gütschow, M. Matriptase-2, a regulatory protease of iron homeostasis: Possible substrates, cleavage sites and inhibitors. Curr. Pharm. Des. 2013, 19, 1052–1061. [Google Scholar]
- Wang, C.Y.; Meynard, D.; Lin, H.Y. The role of TMPRSS6/matriptase-2 in iron regulation and anemia. Front. Pharmacol. 2014, 5, 114. [Google Scholar] [CrossRef] [Green Version]
- Gnana-Prakasam, J.P.; Baldowski, R.B.; Ananth, S.; Martin, P.M.; Smith, S.B.; Ganapathy, V. Retinal expression of the serine protease matriptase-2 (Tmprss6) and its role in retinal iron homeostasis. Mol. Vis. 2014, 20, 561–574. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′–3′) | Size (bp) |
|---|---|---|---|
| SSUSC84_RS00185 | Q0185F | CTTGAAGGGATGGCTGCTGA | 123 |
| Q0185R | CCTCACTAAAACTGATCCCGGA | ||
| SSUSC84_RS00550 | Q0550F | TCGTACGATTGAACAAGCCCA | 125 |
| Q0550R | TAATACGACCGCTGAGACGCT | ||
| SSUSC84_RS01570 | Q1570F | CGATGTTGTCCGCAATGTCAC | 118 |
| Q1570R | AACCATCGCTTCTCCTTGTGC | ||
| SSUSC84_RS03030 | Q3030F | GAAGCTAACATTCGTGGCCG | 112 |
| Q3030R | GAAGCTAACATTCGTGGCCG | ||
| SSUSC84_RS03035 | Q3035F | TCCTTGAGCGTGGTATTGCA | 105 |
| Q3035R | GTCCCGTTGAAACAGGCTCT | ||
| SSUSC84_RS03040 | Q3040F | TGGTTTGACAGATGCATGGC | 112 |
| Q3040R | ACGGCCATCACCACAGTAAA | ||
| SSUSC84_RS03045 | Q3045R | AGTCGTTGCTTCGCCAAAAC | 123 |
| Q3045F | TCTGCTTCTTGGACAACTGGAAT | ||
| SSUSC84_RS03050 | Q3050F | TTCTTCCCGCTCCTTGTTCC | 105 |
| Q3050R | AGTAGAAGCCAAACAGCCGATTT | ||
| SSUSC84_RS06475 | Q6475F | TCTTCCGGAACCTTGATGCC | 130 |
| Q6475R | ACGAGGAAGGCTACGCTCTAGC | ||
| SSUSC84_RS07245 | Q7245F | AAGCCCAACCTGAACCGAAA | 106 |
| Q7245R | TCAGCAGAATTGGCAGCAGA | ||
| 16s RNA | Q16S1 | TAGTCCACGCCGTAAACGATG | 159 |
| Q16S2 | TAAACCACATGCTCCACCGC |
| Group | Sample | Raw Reads | Clean Reads | Q20(%) | Q30(%) | Mapped Reads | Mapping Ratio (%) |
|---|---|---|---|---|---|---|---|
| Fe | Fe1 | 11760266 | 11516070 | 97.91 | 93.85 | 11364301 | 98.68% |
| Fe2 | 12974760 | 12717952 | 98.12 | 94.33 | 12568136 | 98.82% | |
| Fe3 | 11724878 | 11434962 | 98.02 | 94.07 | 11300186 | 98.82% | |
| Co | Co1 | 9204800 | 8993140 | 98.01 | 94.11 | 8870502 | 98.64% |
| Co2 | 12938278 | 12668354 | 98.09 | 94.29 | 12507324 | 98.73% | |
| Co3 | 11516386 | 11277240 | 97.92 | 93.82 | 11127475 | 98.67% | |
| Nor | Nor1 | 11972384 | 11701188 | 97.96 | 93.97 | 11512748 | 98.39% |
| Nor2 | 11584884 | 11384656 | 98.08 | 94.28 | 11203130 | 98.41% | |
| Nor3 | 9594030 | 9393308 | 98.26 | 94.73 | 9259745 | 98.58% |
| Gene | Product | Ferrous Iron Treatment | Cobalt Treatment | ||
|---|---|---|---|---|---|
| Fold Change | Adjusted p-Value | Fold Change | Adjusted p-Value | ||
| SSUSC84_RS03505 | NADH oxidase | −18.734 | 7.50 × 10−184 | −1.641 | 1.63 × 10−4 |
| SSUSC84_RS06530 | thiol peroxidase | −18.065 | 8.93 × 10−89 | −1.690 | 1.43 × 10−5 |
| SSUSC84_RS07245 | superoxide dismutase (SodA) | −14.206 | 6.09 × 10−62 | −2.044 | 2.31 × 10−8 |
| SSUSC84_RS05100 | transcriptional repressor Rex | −12.289 | 2.52 × 10−81 | −2.837 | 1.42 × 10−21 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, M.; Wei, M.; Zhang, Y.; Zheng, C. Transcriptomic Analysis of Streptococcus suis in Response to Ferrous Iron and Cobalt Toxicity. Genes 2020, 11, 1035. https://doi.org/10.3390/genes11091035
Jia M, Wei M, Zhang Y, Zheng C. Transcriptomic Analysis of Streptococcus suis in Response to Ferrous Iron and Cobalt Toxicity. Genes. 2020; 11(9):1035. https://doi.org/10.3390/genes11091035
Chicago/Turabian StyleJia, Mengdie, Man Wei, Yunzeng Zhang, and Chengkun Zheng. 2020. "Transcriptomic Analysis of Streptococcus suis in Response to Ferrous Iron and Cobalt Toxicity" Genes 11, no. 9: 1035. https://doi.org/10.3390/genes11091035
APA StyleJia, M., Wei, M., Zhang, Y., & Zheng, C. (2020). Transcriptomic Analysis of Streptococcus suis in Response to Ferrous Iron and Cobalt Toxicity. Genes, 11(9), 1035. https://doi.org/10.3390/genes11091035