1. Introduction
Monoclonal antibodies (mAbs) are the most effective class of biopharmaceuticals for the treatment of diseases [
1,
2]. In fact, the market for therapeutic mAbs has significantly increased over the past few decades [
3]. In addition to therapeutic mAbs currently in use, numerous mAbs are being developed and tested in clinical trials [
4]. Therefore, the demand for their production is predicted to continuously increase going forward. Presently, most therapeutic mAbs are produced as recombinant proteins using mammalian cultured cells [
5]. However, this approach requires expensive production facilities and complex production control. The high manufacturing costs of therapeutic mAbs creates significant economic and social burdens; therefore, the development of a low-cost alternative technology is imperative for the continued use and development of this class of biopharmaceuticals. Alternative approaches include the development of bioreactors using transgenic plants, insects, and mammals [
5,
6,
7]. Such transgenic organisms do not require expensive facilities and can produce foreign proteins in their products including the plant body, cocoon, and milk, at low cost [
8,
9]. Therefore, they have great potential as alternative methods of mAb production that reduce associated costs and increase efficiency.
Transgenic chickens, which produce functional mAbs in their eggs, provide a useful bioreactor for cost-effective mAb manufacture [
10]. Furthermore, a transgenic chicken bioreactor has several advantages over other transgenic organisms, including short generation time, facile flock expansion, human-like glycosylation profiles, low housing costs, and simple containment requirements [
11]. In 2005, the production of human mAbs in egg whites, using transgenic chimera chickens, was first reported [
12]. In this study, chicken embryonic stem (ES) cells were transfected with DNA vectors designed to specifically express human mAbs in chicken oviduct magnum, and chimera chickens were established via transplantation of the ES cells to recipient embryos. The resulting chimera hens produced 34–148 µg/mL of mAbs in their egg whites; however, transgenic offspring were not obtained from this study. Recently, heritable transgenic chickens capable of producing mAbs in their egg whites were generated using chicken primordial germ cells (PGCs) [
13]. In this study, a gene construct, designed to expresses the mAb in chicken oviduct magnum, was randomly integrated into the genome of the chickens. Functional biosimilar mAbs were produced in the egg white of the transgenic hens; however, expression levels were limited to 2–18 µg/mL. The low production efficiency of ectopic proteins in transgenic chickens can be attributed to gene silencing or the so-called positional effect of transgenes [
14,
15]. Therefore, it is critical that the transgene integrates into the host chicken genome in such a way that avoids translational suppression [
10].
The use of lentiviral vectors is a promising method to avoid silencing of transgenes in a transgenic chicken bioreactor [
16,
17]. However, the packaging limit of the vector could be an obstacle in the introduction of large-size transgenes, which allows the abundant expression of mAbs in the chicken oviduct magnum.
Recently, we developed transgenic chickens that efficiently produced a foreign protein in their egg whites [
18]. In this study, the gene encoding human interferon β (IFN-β) was integrated into a major egg white gene, the ovalbumin gene (
OVA) locus, of PGCs using a CRISPR/Cas9 system (gene knock-in; KI). The KI chickens were then established using the modified PGCs and were found to abundantly produce recombinant human IFN-β in their egg whites (1.9–4.4 mg/mL). Thus, we anticipated abundant mAb production in egg whites using the same approach to generate KI chickens via insertion of mAb genes at the
OVA locus.
The purpose of this study was to examine whether mAbs could efficiently be produced in the egg white of a KI chicken. We integrated the genes encoding humanized anti-HER2 antibody at the initiation site of the OVA locus of PGCs to establish KI hens. We analyzed the mAbs in the eggs laid by the KI hens and compared the binding affinity and selectivity with those of trastuzumab, a commercially available therapeutic anti-HER2 antibody.
2. Materials and Methods
2.1. Animal Experiments
All animal experiments were conducted according to protocols approved by the Institutional Animal Care and Use Committees of the National Institute of Advanced Industrial Science and Technology Tsukuba, Japan (protocol number 2016-115), the Institute of Livestock and Grassland Science (NILGS), National Agriculture and Food Research Organization (NARO), Tsukuba, Japan (protocol number 1611B056), and Cosmo Bio Co. Ltd., Tokyo, Japan, (protocol number PMC201703). Knock-in (KI) and wildtype (WT) chickens were maintained and bred at the animal farm facilities of NARO-NILGS and Cosmo Bio Co. Ltd.
2.2. Plasmid Construction
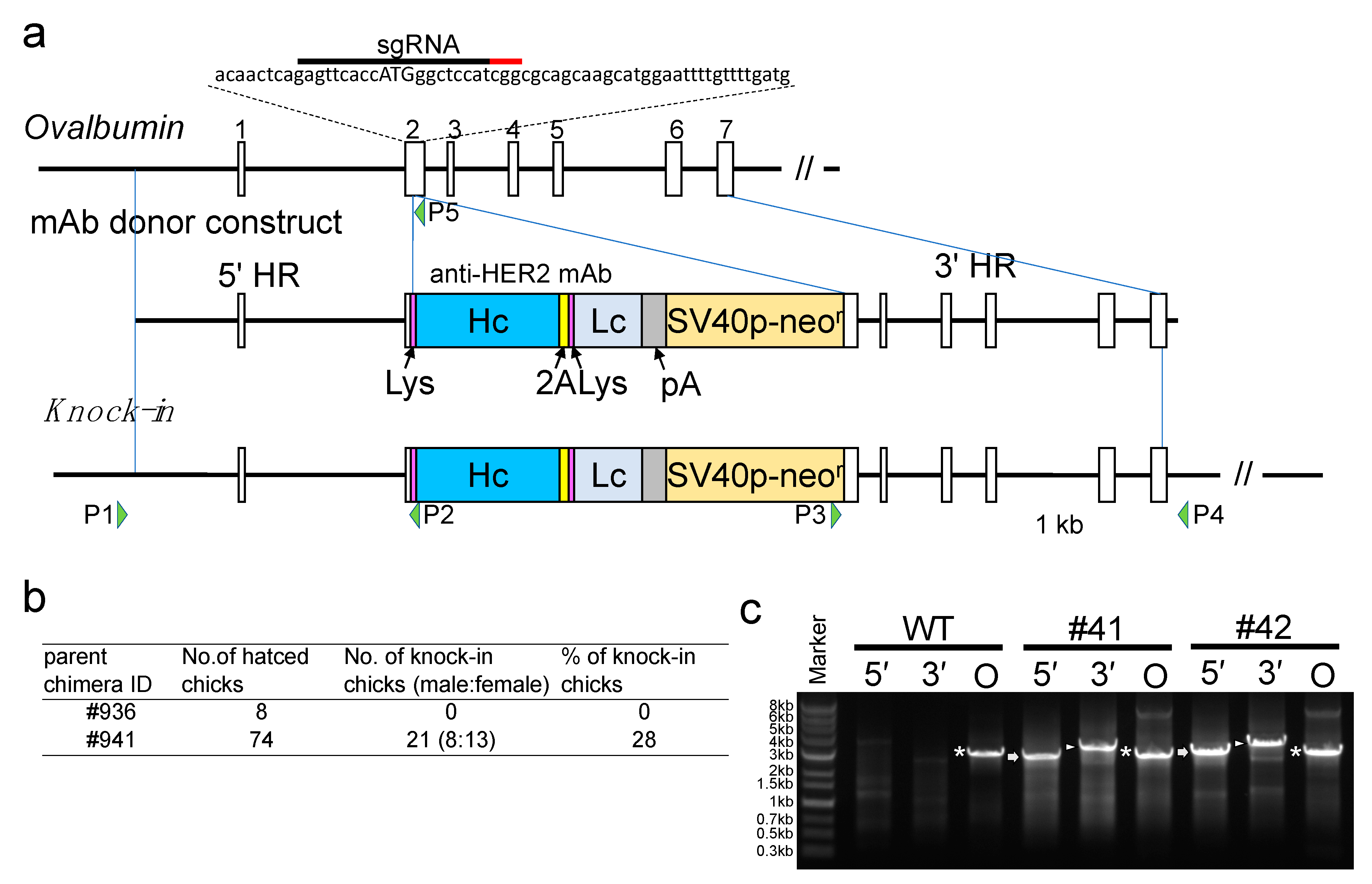
The plasmid expressing hCas9 and single-guide RNA (sgRNA) targeted to OVA (px330-Neo-OVATg2) were generated as described elsewhere [
19]. In the current experiment, the puromycin resistance gene was inserted instead of the neomycin resistance gene in the plasmid (px330-Puro-OVATg2). The donor construct for the mAbs was generated by ligating PCR fragments consisting of 2.8 kb of
OVA DNA (upstream of the ATG initiation codon) as the 5′ homology arm, complementary DNA (cDNA) encoding the heavy and light chains of the anti-HER2 mAb with the lysozyme signal sequence at their respective 5′-ends and linked through the sequence encoding the furin-2A peptide, the bovine growth hormone polyadenylation (bGH-pA) sequence, neomycin resistance gene sequence, and 3.2 kb of the
OVA DNA sequence as the 3′ homology arm (
Figure S1, Supplementary Materials). The donor construct was cloned into the
SalI and
BamHI sites of the pBluescript II SK(+) vector plasmid (Takara Bio, Mountain View, CA, USA) and named the pBS-mAb donor.
2.3. Generation of Knock-In Hens
The KI chickens were generated using essentially the same method as described previously [
16]. Briefly, px330-Puro-OVATg2 and pBS-mAb donor were cotransfected into cultured chicken primordial germ cells (PGCs) and selected with 0.5 mg/mL neomycin for 5 days, following 2 days of culture growth with 1 µg/mL puromycin. The selected cells were proliferated and transplanted into the bloodstream of the recipient chicken embryo to generate germline chimera chickens. A chimeric rooster was crossed with a wildtype hen, and mAb KI offspring were identified by PCR genotyping using MightyAmp DNA Polymerase Ver. 2 (Takara Bio, Kusatsu, Japan). Thermocycling conditions were as follows: 98 °C for 2 min, 35 cycles of 98 °C for 10 s, 60 °C for 3 min, and 72 °C for 30 s. Sequences of primers used in this study were as follows: P1: 5′–ACCTGTGGTGTAGACATCCAGCA–3′, P2: 5′–CCCCAGAGCAGCCAGGGGCAGGAAGCAAAG–3′, P3: 5′–GCACTCTGTCGATACCCCACCGA–3′, P4: 5′–CAACTTCTAGGGCCATACCTGCT–3′, P5: 5′–AAAATTCCATGCTTGCTGCACCGAT–3′. Eggs from two 10 month old KI hens (#41 and #42) were used in this study.
2.4. Preparation of WT and KI Egg White
A disposable medical dropper was used to collect KI and WT egg white samples. The egg whites were homogenized through sonication using an ultrasonic homogenizer (VP050N, TAITEC, Koshigaya, Japan). A portion of each sample was suspended in phosphate-buffered saline (PBS) and appropriately diluted.
2.5. Antibodies
Trastuzumab (Chugai, Tokyo, Japan) was purchased and diluted in PBS. Anti-human immunoglobulin G (hIgG) Fc (I-124, Jackson ImmunoResearch, West Grove, PA, USA), anti-hIgG heavy and light chains (hIgG (H + L), AB_2337577, Jackson ImmunoResearch), and Alexa Fluor goat anti-mouse IgG (H + L) antibodies (Alexa Fluor, A-11029, Thermo Fisher Scientific, Waltham, MA, USA) were also purchased.
2.6. SDS-PAGE and CBB Staining
Egg white samples from WT and mAb KI chickens were suspended in PBS at a ratio of 1:10 for SDS-PAGE. Samples were mixed with an equal volume of 2× Laemmli SDS-PAGE buffer (0.125 M Tris-HCl, pH 6.8, 10% (w/v) sucrose, 4% (w/v) SDS, 10% (v/v) 2-mercaptoethanol, and 0.01% bromophenol blue) and 10 µL of each was subjected to SDS-PAGE (5–20% w/v acrylamide; Oriental Instruments Ltd., Sagamihara, Japan). Samples were prepared for nonreducing SDS-PAGE analysis using 2-mercaptoethanol-free SDS-PAGE sample buffer. The gels were stained with Coomassie Brilliant Blue (CBB) R-250 solution (Nacalai, Kyoto, Japan). Precision Plus Protein Standard (BIO-RAD, Hercules, CA, USA) was used as a protein molecular weight marker.
2.7. Immunoblot Analysis
For immunoblot analysis, WT and mAb KI egg white samples were suspended in PBS at a ratio of 1:100 and dissolved in an equal volume of 2× Laemmli SDS-PAGE buffer, with or without 2-mercaptoethanol. Samples (10 µL each) were separated by SDS-PAGE, and then transferred onto a polyvinylidene difluoride membrane (Immobilon-P; Millipore, Bedford, MA, USA) via electroblotting. After blocking with 1% bovine serum albumin in 10 mM Tris-HCl containing 150 mM NaCl and 0.1% Tween-20 (TBS-T), the membrane was subjected to immunoblotting with antibodies against hIgG Fc or hIgG (H + L) at a 6000-fold dilution. Proteins were visualized with horseradish peroxidase-conjugated anti-mouse (for hIgG Fc) (Jackson ImmunoResearch) and anti-rabbit IgG (for hIgG (H + L)) (Jackson ImmunoResearch) using an enhanced chemiluminescence system (ImmunoStar reagent; Wako, Osaka, Japan). The immunoreactive bands were analyzed with ImageQuant LAS 500 (GE Healthcare, Wauwatosa, WI, USA) and the relative intensity of the bands was determined using the ImageJ software (National Institutes of Health, Bethesda, MD, USA).
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
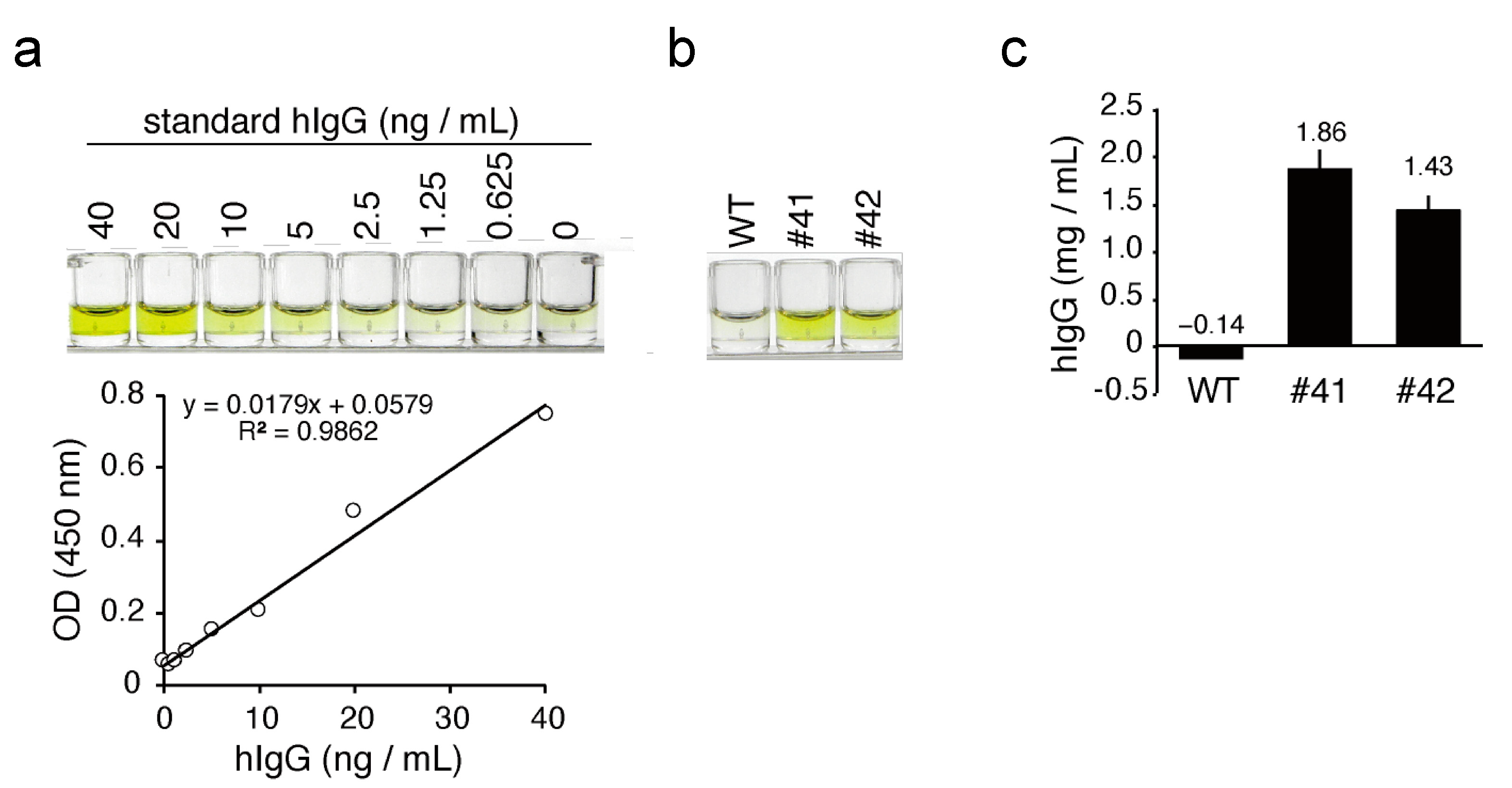
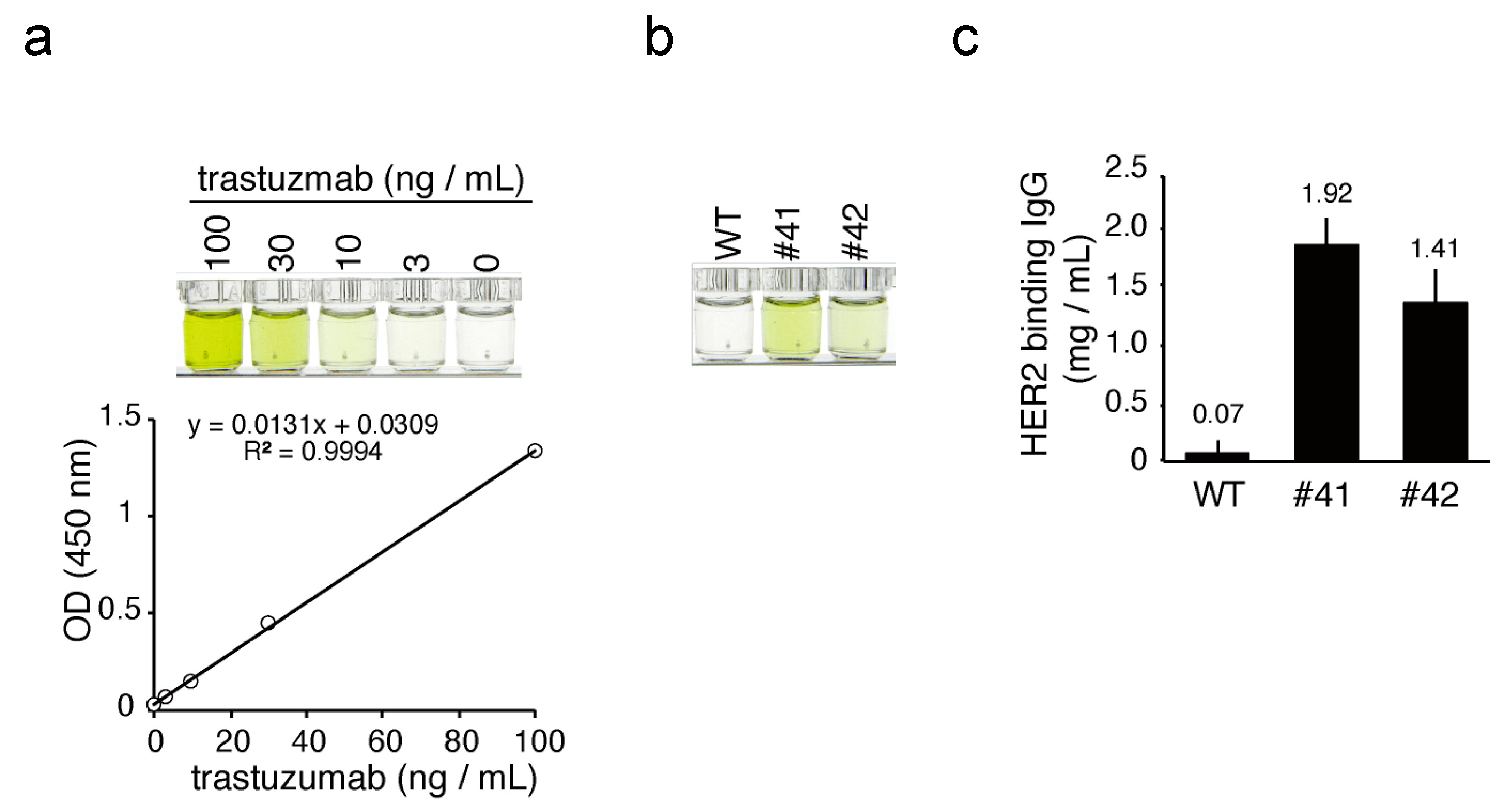
The concentration of hIgG in KI egg white was determined using human immunoglobulin G (IgG) AssayMax ELISA kit from AssayPro (St. Charles, MO, USA), according to the protocols provided by the manufacturer. The HER2 antigen binding activity of mAbs in KI egg white was evaluated using a Herceptin ELISA kit (α diagnostic International San Antonio, TX, USA), with commercial trastuzumab as a reference. The color generated in each ELISA was quantified using a microplate reader (iMark, BIO-RAD).
2.9. Immunofluorescence
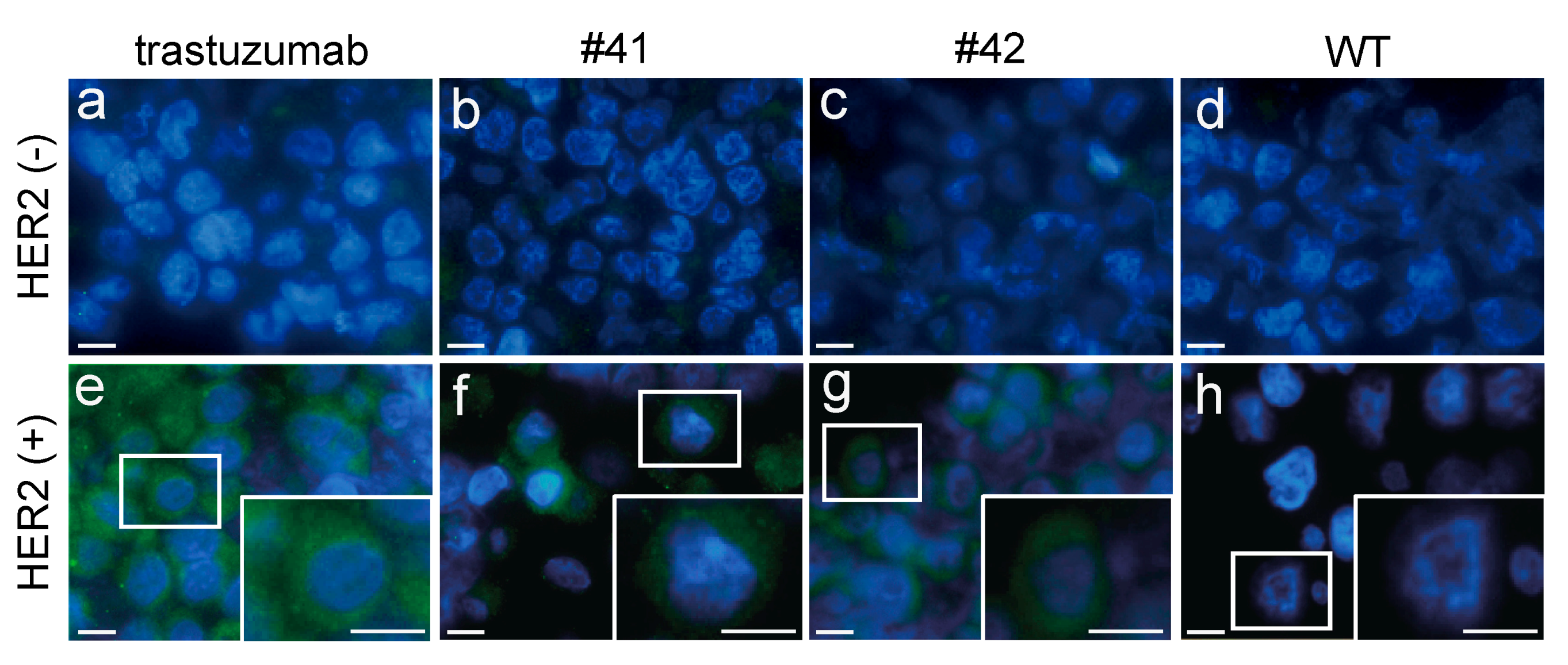
Glass slides mounted with 4–5 μm thick slices of formalin-fixed paraffin-embedded human HER2-positive and -negative cells (POSICON-slide HER2 IHC, Pathology Institute, Toyama, Japan) were purchased and used for immunofluorescence. The sections on the glass slides were deparaffinized and rehydrated sequentially with xylene, ethanol, and water. The sections were treated with blocking solution (Blocking ONE histo, Nacalai) for 30 min, and then incubated with 1 µg/mL anti-HER2 antibodies (diluted KI egg white or trastuzumab) for 30 min at room temperature. After a secondary antibody reaction with mouse anti-hIgG Fc (1:1000, I-124) for 30 min, sections were visualized with Alexa Fluor 488 goat anti-mouse IgG (H + L) antibody (1:1000). The sections were mounted using Fluoroshield with DAPI (H-1200, ImmunoBioScience Corp, Mukilteo, WA, USA), and images were obtained using a fluorescence microscope (BZ-800; Keyence, Osaka, Japan).
4. Discussion
In this study, we efficiently produced fully assembled anti-HER2 mAbs in egg whites by integrating the mAb gene at the initiation site of the OVA locus. We further demonstrated that anti-HER2 mAbs deposited in the KI egg white showed similar HER2-binding properties in comparison to the commercially available therapeutic antibody, trastuzumab.
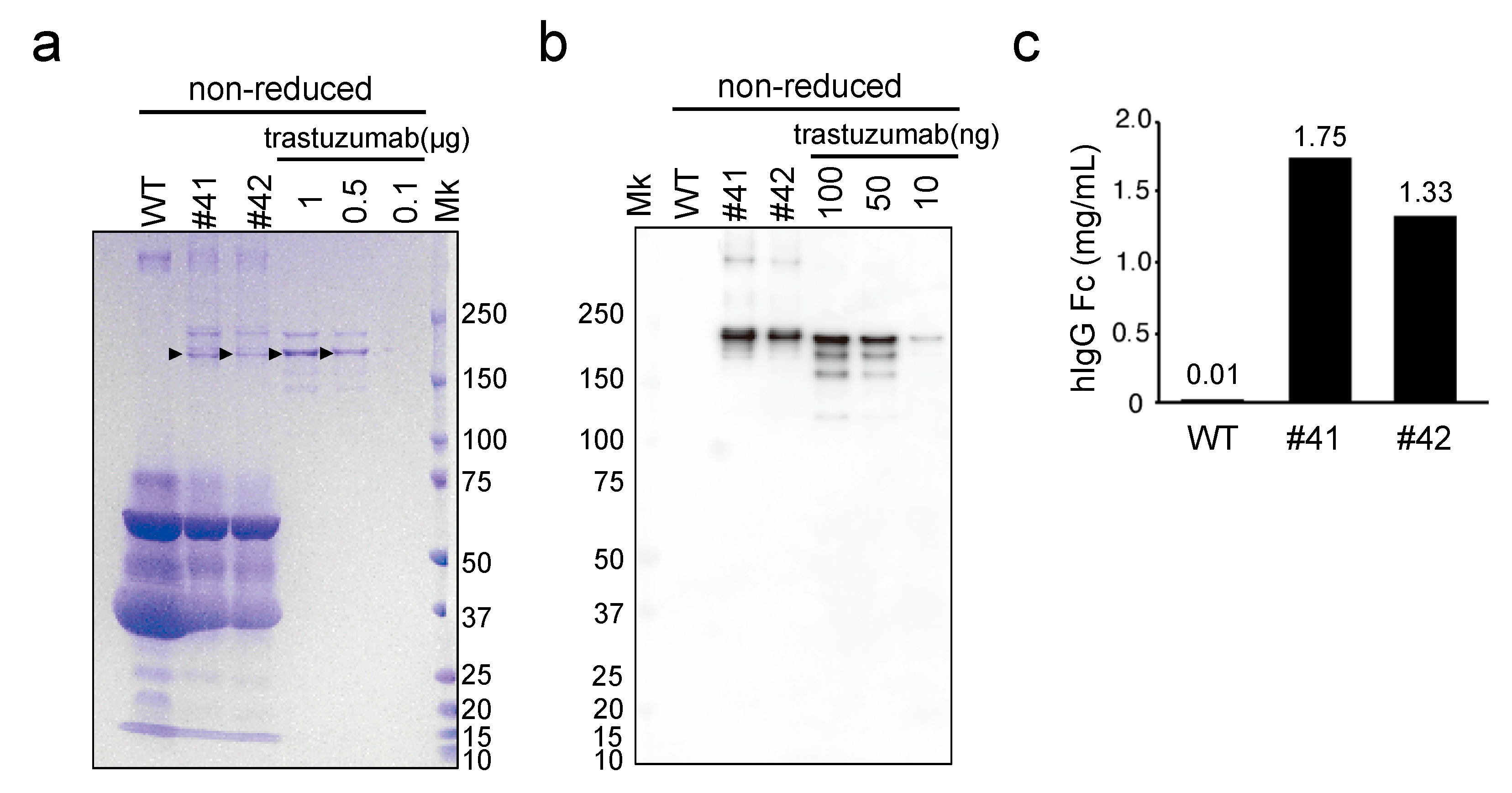
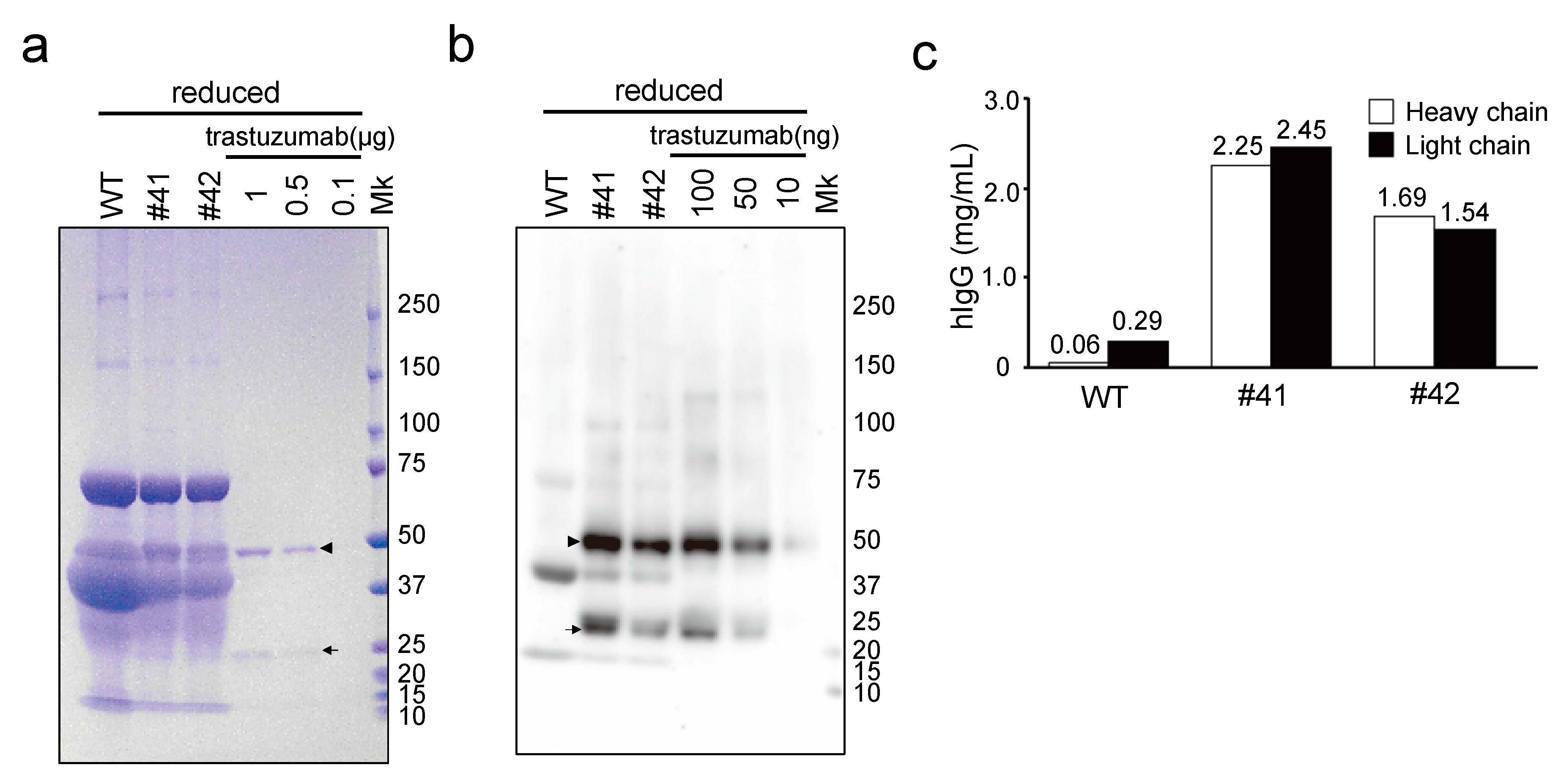
The concentration of mAbs deposited in the egg whites of KI chickens was in the 1.4–1.9 mg/mL range (
Figure 4c), which is 80- to 1000-fold higher than the previously reported mAb expression levels in similar transgenic chickens employing 3.5 kb of the
OVA promoter, to drive expression, with random gene integration [
13]. Similarly, in our previous study, the concentration of human IFN-β deposited in the egg whites of KI chickens was 1.9–4.4 mg/mL [
18], which is 15- to 1200-fold higher than other reports using transgenic chickens for human IFN-β production with 2.8 kb of the
OVA promoter, to drive expression and random integration of
IFN-β [
16]. In both our present and our previous KI chicken studies, the foreign genes were integrated into the initiation site of
OVA. This approach resulted in abundant levels of foreign proteins deposited into the KI egg whites, in comparison to transgenic systems employing random gene integration. Therefore, it is conceivable that transgene insertion at the
OVA locus is a reliable method to establish an efficient transgenic chicken bioreactor.
The antigen binding of anti-HER2 mAbs was analyzed directly using diluted KI egg whites, without purification (
Figure 5c). Nevertheless, the anti-HER2 mAbs exhibited almost equal antigen binding affinity to that of trastuzumab, a therapeutic anti-HER2 antibody. This result indicates that fully assembled mAbs were efficiently synthesized in KI chicken magnum, and stably deposited into the egg whites. Thus, it is anticipated that the chicken bioreactor system developed in this study is well suited for mAb production and has great potential as an alternative method for commercial manufacturing. In addition, the development of a more cost-effective approach to mAb production is highly desirable. Although production costs are dependent on many variables, including production scale and methods, we estimate the cost of 1 g of mAb to be >40 USD, using both cell culture and plant bioreactor systems [
21,
22,
23]. According to our current study, a KI egg with 30 mL of egg white would contain 42–57 mg of mAbs. Because commercial egg production costs are approximately
$1 per dozen eggs [
24], 1 g of mAbs could be produced for 1.5–2.0 USD using our chicken bioreactor system. It is important to note that this is a rough estimate, as it does not consider several parameters, including costs associated with the development of KI chickens, their genotyping, and the egg-laying period of KI hens. However, these factors are limited, and we believe that our chicken bioreactor system represents a cost-effective upstream process for therapeutic mAb production. For commercial manufacturing of therapeutic mAbs, the costs incurred in the downstream process (i.e., mAb purification) and approvals (or drug certification) by regulatory bodies should also be considered. However, the cost advantage of the chicken bioreactor system vis-à-vis the other mAb production systems has not been evaluated. It was surmised that the costs for downstream processing of mAbs produced using the chicken bioreactor should be equivalent to those for mAbs produced using the mammalian cell culture methods [
17]. Although the costs for regulatory approvals are not in the public domain, recently, a therapeutic recombinant protein produced using a chicken bioreactor was approved by the FDA, suggesting that these processes are not unrealistically expensive [
25].
It is estimated that the typical production scale of commercial therapeutic mAbs is on the order of hundreds of kg/year or less [
26]. To meet this demand using the chicken bioreactor system, tens of thousands of KI hens will be required annually. This is feasible considering the high fertility of chickens. Furthermore, it is expected that one transgenic rooster could be used to breed over a hundred thousand transgenic offspring in a single year [
11]. In addition, a chicken flock is easily scalable; thus, the system can be adapted to suit variable levels of production demands. Therefore, the chicken bioreactor system is a suitable approach for the manufacture of commercial therapeutic mAbs.
In conclusion, a chicken bioreactor system, based on mAb gene KI at the OVA locus, can efficiently produce mAbs with adequate antigen binding capacity and has the potential to be a viable alternative production system for commercial mAbs. Furthermore, the approach is cost-effective and easily scalable. Although further studies are required to establish downstream processes that will be required to enable medical applications, the production of therapeutic mAbs using KI chicken bioreactors is a promising cost-effective technology for the production of therapeutic mAbs.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





