Identification of microRNA-Associated-ceRNA Networks Regulating Crop Milk Production in Pigeon (Columba livia)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Sample Collection
2.3. RNA Isolation and Quality Assessment
2.4. miRNA Library Construction and Sequencing
2.5. Quality Control, Mapping and Acquisition of Sequencing Data
2.6. Identification of miRNA, lncRNA and mRNA
2.7. Differentially Expressed Analysis of RNAs
2.8. miRNA Target Prediction
2.9. Functional Enrichment
2.10. Protein-Protein Interaction Analyze
2.11. Construction of ceRNA Network
2.12. Quantitative Real-Time PCR
2.13. Data Accessibility
3. Results
3.1. Identification of Pigeon Small RNAs
3.2. miRNA Target Prediction and Analyze
3.3. PPI Network
3.4. miRNA-mRNA-lncRNA/circRNA Interaction Network
3.5. Functional Annotation of the mRNAs in ceRNA Network
3.6. Construct Fatty Acid Biosynthesis Pathway Function Network
3.7. The Exploration of ceRNA Regulates Axes
3.8. Validation of Differentially Expressed RNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
Abbreviations
| MREs | miRNA response elements |
| GO | Gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
| ceRNA | Competing endogenous RNA |
| HPRT1 | Hypoxanthine phosphoribosyltransferase 1 |
| DEGs | differentially expressed genes |
| PPI | Protein protein interaction |
| EPRS | Glutamyl-prolyl tRNA synthetase |
| SREBF1 | Sterol regulatory element-binding protein 1 |
| ACACA | Acetyl-CoA carboxylase α |
| PPP1CA | Protein phosphatase type 1 α |
| CREBRF | CREB3 regulatory factor |
| ELOVL4 | ELOVL fatty acid elongase 4 |
| ELOVL5 | ELOVL fatty acid elongase 5 |
| ELOVL6 | ELOVL fatty acid elongase 6 |
| ELOVL7 | ELOVL fatty acid elongase 7 |
| HGF | Hepatocyte growth factor |
| LAMC2 | Laminin subunit γ 2 |
| ITGB8 | Integrin subunit β 8 |
| PDGFRB | Platelet derived growth factor receptor β |
| ADRB1 | Adrenoceptor β 1 |
| PIK3CD | Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta |
| CDH1 | E-cadherin |
| DOT1L | DOT1 like histone lysine methyltransferase |
| ESRP2 | Epithelial splicing regulatory protein 2 |
| TTC28 | Tetratricopeptide repeat domain 28 |
| IGF2 | Insulin-like growth factor 2 |
| CDH15 | M-cadherin |
| MAGUK | Membrane associated guanylate kinases |
| ADRB3 | Adrenoceptor β 3 |
| DHCR7 | 7-dehydrocholesterol reductase |
| IHH | Indian Hedgehog Regulates |
| GRHL2 | Grainy head like transcription factor 2 |
| EMT | Epithelial mesenchymal transition |
| MET | Mesenchymal epithelial transition |
| KIF26B | Kinesin family member 26B |
| PUFA | polyunsaturated fatty acid |
| VLC-FA | very long-chain fatty acids |
References
- Anderson, T.R.; Pitts, D.S.; Nicoll, C.S. Prolactin’s mitogenic action on the pigeon crop-sac mucosal epithelium involves direct and indirect mechanisms. Gen. Comp. Endocrinol. 1984, 54, 236–246. [Google Scholar] [CrossRef]
- Griminger, P. Physiology and Behaviour of the Pigeon; Academic Press: London, UK, 1983. [Google Scholar]
- Horseman, N.D.; Buntin, J.D. Regulation of pigeon cropmilk secretion and parental behaviors by prolactin. Annu. Rev. Nutr. 1995, 15, 213–238. [Google Scholar] [CrossRef]
- Davies, W.L. The composition of the crop milk of pigeons. Biochem. J. 1939, 33, 898–901. [Google Scholar] [CrossRef] [Green Version]
- Bharathi, L.; Shenoy, K.B.; Mojamdar, M.; Hegde, S.N. In vitro growth-stimulatory property of pigeon milk. Biochem. Cell Biol. 1993, 71, 303–307. [Google Scholar] [CrossRef]
- Desmeth, M.V.J. Lipid composition of pigeon cropmilk—I. Total lipids and lipid classes. Comp. Biochem. Physiol. Part B 1980, 1, 129–133. [Google Scholar] [CrossRef]
- Shetty, S.; Hegde, S.N. Stimulatory effect of pigeon milk growth factor on protein and nucleic acid synthesis in neonate mice. Biol. Neonate 1992, 62, 409–415. [Google Scholar] [CrossRef]
- Hegde, S.N. A note on the comparative weight increase in the nidicolous pigeon squab and the nidifugous domestic chick during the first four postnatal weeks. Indian Zool 1970, 1, 1–6. [Google Scholar]
- Hegde, S.N. The composition of pigeon milk and its effects on growth in chicks. Ind. J. Exp. Biol. 1973, 11, 238–239. [Google Scholar]
- Shetty, S.; Hegde, S.N. Pigeon milk: A new source of growth factor. Experientia 1993, 49, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Bharathi, L.; Shenoy, K.B.; Hegde, S.N. In vivo and in vitro growth-stimulatory effects of pigeon milk. Comp. Biochem. Physiol. Comp. Physiol. 1994, 108, 451–459. [Google Scholar] [CrossRef]
- König, H.E.; Korbel, R.; Liebich, H. Avian Anatomy; 5M Publishing Ltd., 8 Smithy Wood Drive: Sheffield, UK, 2016. [Google Scholar]
- Hu, X.C.; Gao, C.Q.; Wang, X.H.; Yan, H.C.; Chen, Z.S.; Wang, X.Q. Crop milk protein is synthesised following activation of the IRS1/Akt/TOR signalling pathway in the domestic pigeon (Columba livia). Br. Poult. Sci. 2016, 57, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Fu, Z.; Jiang, S.G.; Wang, X.Q.; Yan, H.C.; Gao, C.Q. Targeted disruption of TORC1 retards young squab growth by inhibiting the synthesis of crop milk protein in breeding pigeon (Columba livia). Poult. Sci. 2019, 99, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.Y.; Fu, Z.; Pan, N.X.; Yan, H.C.; Wang, X.Q.; Gao, C.Q. Leucine promotes the growth of squabs by increasing crop milk protein synthesis through the TOR signaling pathway in the domestic pigeon (Columba livia). Poult. Sci. 2019, 98, 5514–5524. [Google Scholar] [CrossRef]
- Xie, P.; Wang, X.P.; Bu, Z.; Zou, X.T. Differential expression of fatty acid transporters and fatty acid synthesis-related genes in crop tissues of male and female pigeons (Columba livia domestica) during incubation and chick rearing. Br. Poult. Sci. 2017, 58, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.J.; Crowley, T.M.; Haring, V.R.; Wilson, S.L.; Harper, J.A.; Payne, J.S.; Green, D.; Monaghan, P.; Stanley, D.; Donald, J.A.; et al. Transcriptome analysis of pigeon milk production—Role of cornification and triglyceride synthesis genes. BMC Genom. 2013, 14, 169. [Google Scholar] [CrossRef] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Thomson, D.W.; Dinger, M.E. Endogenous microRNA sponges: Evidence and controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.; Lieberman, J.; Lal, A. Desperately seeking microRNA targets. Nat. Struct. Mol. Biol. 2010, 17, 1169–1174. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, J.S.; Shen, Y.F.; Mao, H.G.; Xu, N.Y. MicroRNA expression profiling of lactating mammary gland in divergent phenotype swine breeds. Int. J. Mol. Sci. 2015, 16, 1448–1465. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Moisá, S.; Khan, M.J.; Wang, J.; Bu, D.; Loor, J.J. MicroRNA expression patterns in the bovine mammary gland are affected by stage of lactation. J. Dairy Sci. 2012, 95, 6529–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avril-Sassen, S.; Goldstein, L.D.; Stingl, J.; Blenkiron, C.; Le Quesne, J.; Spiteri, I.; Karagavriilidou, K.; Watson, C.J.; Tavaré, S.; Miska, E.A.; et al. Characterisation of microRNA expression in post-natal mouse mammary gland development. BMC Genom. 2009, 10, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucar, A.; Vafaizadeh, V.; Jarry, H.; Fiedler, J.; Klemmt, P.A.; Thum, T.; Groner, B.; Chowdhury, K. miR-212 and miR-132 are required for epithelial stromal interactions necessary for mouse mammary gland development. Nat. Genet. 2010, 42, 1101–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.; Yoon, K.S.; Cho, K.W.; Kim, K.S.; Jung, H.S. Expression of miR-206 during the initiation of mammary gland development. Cell Tissue Res. 2013, 353, 425–433. [Google Scholar] [CrossRef]
- Manjarín, R.; Steibel, J.P.; Kirkwood, R.N.; Taylor, N.P.; Trottier, N.L. Transcript abundance of hormone receptors, mammalian target of rapamycin pathway-related kinases, insulin-like growth factor I, and milk proteins in porcine mammary tissue. J. Anim. Sci. 2012, 90, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Ni, A.; Ge, P.; Li, Y.; Shi, L.; Wang, P.; Fan, J.; Isa, A.M.; Sun, Y.; Chen, J. Analysis of Long Non-Coding RNAs and mRNAs Associated with Lactation in the Crop of Pigeons (Columba livia). Genes 2020, 11, 201. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory efficient alignment of short DNA sequences to the human genome. Genome. Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome. Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Gillespie, M.J.; Haring, V.R.; McColl, K.A.; Monaghan, P.; Donald, J.A.; Nicholas, K.R.; Moore, R.J.; Crowley, T.M. Histological and global gene expression analysis of the ‘lactating’ pigeon crop. BMC Genom. 2011, 12, 452. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zheng, Y.; Li, X.; Gao, Q.; Feng, T.; Zhang, P.; Liao, M.; Tian, X.E.; Lu, H.; Zeng, W. Profiling of miRNAs in porcine Sertoli cells. J. Anim. Sci. Biotechnol. 2020, 11, 85. [Google Scholar] [CrossRef]
- Aryal, B.; Singh, A.K.; Rotllan, N.; Price, N.; Fernández-Hernando, C. MicroRNAs and lipid metabolism. Curr. Opin. Lipidol. 2017, 28, 273–280. [Google Scholar] [CrossRef]
- Goody, D.; Pfeifer, A. MicroRNAs in brown and beige fat. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 29–36. [Google Scholar] [CrossRef]
- Litwer, G. Die Histologischen Veränderungen der Kropfwandung beiTauben, zur Zeit der Bebrütung und Ausfütterung ihrer Jungen. Z. Zellforsch. Mikrosk. Anat. 1926, 3, 695–722. [Google Scholar] [CrossRef]
- Weber, W. Zur Histologie und Cytologie der Kropfmilchbildung derTaube. Z. Zellforsch. Mikrosk. Anat. 1962, 56, 247–276. [Google Scholar] [CrossRef]
- Qi, J.; Yang, Z.; Zhang, Y.; Lu, B.; Yin, Y.; Liu, K.; Xue, W.; Qu, C.; Li, W. miR-20b-5p, TGFBR2, and E2F1 Form a Regulatory Loop to Participate in Epithelial to Mesenchymal Transition in Prostate Cancer. Front. Oncol. 2020, 9, 1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ansari, M.M.; Aboussekhra, A. miR-146b-5p mediates p16-dependent repression of IL-6 and suppresses paracrine procarcinogenic effects of breast stromal fibroblasts. Oncotarget 2015, 6, 30006–30016. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, S.W.; Han, J.S.; Shin, S.P.; Lee, S.I.; Park, T.S. Functional analyses of miRNA-146b-5p during myogenic proliferation and differentiation in chicken myoblasts. BMC Mol. Cell Biol. 2020, 21, 40. [Google Scholar] [CrossRef]
- Cai, M.; Shao, W.; Yu, H.; Hong, Y.; Shi, L. Paeonol Inhibits Cell Proliferation, Migration and Invasion and Induces Apoptosis in Hepatocellular Carcinoma by Regulating miR-21-5p/KLF6 Axis. Cancer Manag. Res. 2020, 12, 5931–5943. [Google Scholar] [CrossRef]
- Yang, Z.; Liao, B.; Xiang, X.; Ke, S. miR-21-5p promotes cell proliferation and G1/S transition in melanoma by targeting CDKN2C. FEBS Open Bio 2020, 10, 752–760. [Google Scholar] [CrossRef]
- Yan, L.; Ma, J.; Wang, Y.; Zan, J.; Wang, Z.; Zhu, Y.; Zhu, Y.; Ling, L.; Cao, L.; Liu, X.; et al. miR-21-5p induces cell proliferation by targeting TGFBI in non-small cell lung cancer cells. Exp. Ther. Med. 2018, 16, 4655–4663. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Zhu, K.; Wang, Y.; Yu, H.; Guo, J. Overexpression of miR-21-5p promotes proliferation and invasion of colon adenocarcinoma cells through targeting CHL1. Mol. Med. 2018, 24, 36. [Google Scholar] [CrossRef] [Green Version]
- Yi, F.; Shang, Y.; Li, B.; Dai, S.; Wu, W.; Cheng, L.; Wang, X. MicroRNA-193-5p modulates angiogenesis through IGF2 in type 2 diabetic cardiomyopathy. Biochem. Biophys. Res. Commun. 2017, 491, 876–882. [Google Scholar] [CrossRef]
- Luo, Y.; Ji, H.; Cao, Y.; Ding, X.; Li, M.; Song, H.; Li, S.; WaTableng, C.; Wu, H.; Meng, J.; et al. miR-26b-5p/TCF-4 Controls the Adipogenic Differentiation of Human Adipose-derived Mesenchymal Stem Cells. Cell Transplant. 2020, 29, 963689720934418. [Google Scholar] [CrossRef] [PubMed]
- Arif, A.; Terenzi, F.; Potdar, A.A.; Jia, J.; Sacks, J.; China, A.; Halawani, D.; Vasu, K.; Li, X.; Brown, J.M.; et al. EPRS is a critical mTORC1-S6K1 effector that influences adiposity in mice. Nature 2017, 542, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ding, D.; Huang, Q.; Liu, Q.; Lu, H.; Lu, Y.; Chi, Y.; Sun, X.; Ye, G.; Zhu, H.; et al. Downregulation of miR-192 causes hepatic steatosis and lipid accumulation by inducing SREBF1: Novel mechanism for bisphenol A-triggered non-alcoholic fatty liver disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 869–882. [Google Scholar] [CrossRef]
- Bruner, H.C.; Derksen, P. Loss of E-Cadherin-Dependent Cell-Cell Adhesion and the Development and Progression of Cancer. Cold Spring Harb. Perspect. Biol. 2018, 10, a029330. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.; Schlue, K.T.; Kniffin, A.F.; Mayer, C.R.; Duke, A.A.; Narayanan, V.; Arsenovic, P.T.; Bathula, K.; Danielsson, B.E.; Dumbali, S.P.; et al. Spatial Proliferation of Epithelial Cells Is Regulated by E-Cadherin Force. Biophys. J. 2018, 115, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, S.; Hultin, S.; Subramani, A.; Petropoulos, S.; Zhang, Y.; Cao, X.; Mpindi, J.; Kalloniemi, O.; Johansson, S.; Majumdar, A.; et al. The E-cadherin/AmotL2 complex organizes actin filaments required for epithelial hexagonal packing and blastocyst hatching. Sci. Rep. 2017, 7, 9540. [Google Scholar] [CrossRef]
- Shigetomi, K.; Ikenouchi, J. Cell Adhesion Structures in Epithelial Cells Are Formed in Dynamic and Cooperative Ways. Bioessays 2019, 41, e1800227. [Google Scholar] [CrossRef]
- Shigetomi, K.; Ono, Y.; Inai, T.; Ikenouchi, J. Adherens junctions influence tight junction formation via changes in membrane lipid composition. J. Cell Biol. 2018, 217, 2373–2381. [Google Scholar] [CrossRef] [Green Version]
- Dailey, M.O. Expression of T lymphocyte adhesion molecules: Regulation during antigen-induced T cell activation and differentiation. Crit. Rev. Immunol. 1998, 18, 153–184. [Google Scholar] [CrossRef]
- Federica, S.; Francesco, N.; Giovanna, D.M.; Carmela, S.M.; Gennaro, C.; Carmela, T.; Bianca, M. Identification of novel single nucleotide polymorphisms in promoter III of the acetyl-CoA carboxylase-{alpha} gene in goats affecting milk production traits. J. Hered. 2009, 100, 386–389. [Google Scholar] [CrossRef]
- Moioli, B.; Scatà, M.C.; De Matteis, G.; Annicchiarico, G.; Catillo, G.; Napolitano, F. The ACACA gene is a potential candidate gene for fat content in sheep milk. Anim. Genet. 2013, 44, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.C.; Clegg, R.A.; Travers, M.T.; Vernon, R.G. Lipid metabolism in the lactating mammary gland. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1997, 1347, 101–126. [Google Scholar] [CrossRef]
- Cao, W.; Liu, Z.; Guo, F.; Yu, J.; Li, H.; Yin, X. Adipocyte ADRB3 Down-Regulated in Chinese Overweight Individuals Adipocyte ADRB3 in Overweight. Obes. Facts 2018, 11, 524–533. [Google Scholar] [CrossRef]
- Lee, Y.H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In vivo identification of bipotential adipocyte progenitors recruited by β3-adrenoceptor activation and high-fat feeding. Cell Metab. 2012, 15, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Bordicchia, M.; Pocognoli, A.; D’Anzeo, M.; Siquini, W.; Minardi, D.; Muzzonigro, G.; Dessì-Fulgheri, P.; Sarzani, R. Nebivolol induces, via β3 adrenergic receptor, lipolysis, uncoupling protein 1, and reduction of lipid droplet size in human adipocytes. J. Hypertens 2014, 32, 389–396. [Google Scholar] [CrossRef]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Franquet, E.E.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab. 2015, 21, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Miklas, J.W.; Clark, E.; Levy, S.; Detraux, D.; Leonard, A.; Beussman, K.; Showalter, M.R.; Smith, A.T.; Hofsteen, P.; Yang, X.; et al. TFPa/HADHA is required for fatty acid beta-oxidation and cardiolipin re-modeling in human cardiomyocytes. Nat. Commun. 2019, 10, 4671. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Carrio, J.; Alperi-López, M.; Naves-Díaz, M.; Dusso, A.; López, P.; Ballina-García, F.J.; Cannata-Andía, J.B.; Suárez, A. Vitamin D Receptor Polymorphism and DHCR7 Contribute to the Abnormal Interplay Between Vitamin D and Lipid Profile in Rheumatoid Arthritis. Sci. Rep. 2019, 9, 2546. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Guillou, H.; Zadravec, D.; Martin, P.G.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef]
- Cameron, D.J.; Tong, Z.; Yang, Z.; Kaminoh, J.; Kamiyah, S.; Chen, H.; Zeng, J.; Chen, Y.; Luo, L.; Zhang, K. Essential role of Elovl4 in very long chain fatty acid synthesis, skin permeability barrier function, and neonatal survival. Int. J. Biol. Sci. 2007, 3, 111–119. [Google Scholar] [CrossRef]
- Li, W.; Sandhoff, R.; Kono, M.; Zerfas, P.; Hoffmann, V.; Ding, B.C.; Proia, R.L.; Deng, C.X. Depletion of ceramides with very long chain fatty acids causes defective skin permeability barrier function, and neonatal lethality in ELOVL4 deficient mice. Int. J. Biol. Sci. 2007, 3, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaka, T.; Shimano, H. Elovl6: A new player in fatty acid metabolism and insulin sensitivity. J. Mol. Med. 2009, 87, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Kosinski, C.; Stange, D.E.; Xu, C.; Chan, A.S.; Ho, C.; Yuen, S.T.; Mifflin, R.C.; Powell, D.W.; Clevers, H.; Leung, S.Y.; et al. Indian hedgehog regulates intestinal stem cell fate through epithelial-mesenchymal interactions during development. Gastroenterology 2010, 139, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Boglev, Y.; Wilanowski, T.; Caddy, J.; Parekh, V.; Auden, A.; Darido, C.; Hislop, N.R.; Cangkrama, M.; Ting, S.B.; Jane, S.M. The unique and cooperative roles of the Grainy head-like transcription factors in epidermal development reflect unexpected target gene specificity. Dev. Biol. 2011, 349, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Chung, V.Y.; Tan, T.Z.; Tan, M.; Wong, M.K.; Kuay, K.T.; Yang, Z.; Ye, J.; Muller, J.; Koh, C.M.; Guccione, E.; et al. GRHL2-miR-200-ZEB1 maintains the epithelial status of ovarian cancer through transcriptional regulation and histone modification. Sci. Rep. 2016, 6, 19943. [Google Scholar] [CrossRef] [Green Version]
- Warzecha, C.C.; Sato, T.K.; Nabet, B.; Hogenesch, J.B.; Carstens, R.P. ESRP1 and ESRP2 are epithelial cell-type-specific regulators of FGFR2 splicing. Mol. Cell 2009, 33, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Bebee, T.W.; Park, J.W.; Sheridan, K.I.; Warzecha, C.C.; Cieply, B.W.; Rohacek, A.M.; Xing, Y.; Carstens, R.P. The splicing regulators Esrp1 and Esrp2 direct an epithelial splicing program essential for mammalian development. eLife 2015, 4, e08954. [Google Scholar] [CrossRef]
- Sun, X.J.; Li, Y.L.; Wang, L.G.; Liu, L.Q.; Ma, H.; Hou, W.H.; Yu, J.M. Mastl overexpression is associated with epithelial to mesenchymal transition and predicts a poor clinical outcome in gastric cancer. Oncol. Lett. 2017, 14, 7283–7287. [Google Scholar] [CrossRef]
- Simeone, P.; Trerotola, M.; Franck, J.; Cardon, T.; Marchisio, M.; Fournier, I.; Salzet, M.; Maffia, M.; Vergara, D. The multiverse nature of epithelial to mesenchymal transition. Semin. Cancer Biol. 2019, 58, 1–10. [Google Scholar] [CrossRef]
- Antony, J.; Thiery, J.P.; Huang, R.Y. Epithelial-to-mesenchymal transition: Lessons from development, insights into cancer and the potential of EMT-subtype based therapeutic intervention. Phys. Biol. 2019, 16, 041004. [Google Scholar] [CrossRef]
- Uchiyama, Y.; Sakaguchi, M.; Terabayashi, T.; Inenaga, T.; Inoue, S.; Kobayashi, C.; Oshima, N.; Kiyonari, H.; Nakagata, N.; Sato, Y.; et al. Kif26b, a kinesin family gene, regulates adhesion of the embryonic kidney mesenchyme. Proc. Natl. Acad. Sci. USA 2010, 107, 9240–9245. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zou, J.; Guan, Y.; Zhang, Y.; Zhang, W.; Zhou, X.; Xiong, C.; Tolbert, E.; Zhao, T.C.; Bayliss, G.; et al. Blocking the histone lysine 79 methyltransferase DOT1L alleviates renal fibrosis through inhibition of renal fibroblast activation and epithelial-mesenchymal transition. FASEB J. 2019, 33, 11941–11958. [Google Scholar] [CrossRef] [Green Version]
- Tangye, S.G.; Bier, J.; Lau, A.; Nguyen, T.; Uzel, G.; Deenick, E.K. Immune Dysregulation and Disease Pathogenesis due to Activating Mutations in PIK3CD-the Goldilocks’ Effect. J. Clin. Immunol. 2019, 39, 148–158. [Google Scholar] [CrossRef]
- Bier, J.; Rao, G.; Payne, K.; Brigden, H.; French, E.; Pelham, S.J.; Lau, A.; Lenthall, H.; Edwards, E.; Smart, J.M.; et al. Activating mutations in PIK3CD disrupt the differentiation and function of human and murine CD4(+) T cells. J. Allergy Clin. Immunol. 2019, 144, 236–253. [Google Scholar] [CrossRef] [Green Version]
- Kiyota, K.; Yoshiura, K.I.; Houbara, R.; Miyahara, H.; Korematsu, S.; Ihara, K. Auto-immune disorders in a child with PIK3CD variant and 22q13 deletion. Eur. J. Med. Genet. 2018, 61, 631–633. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
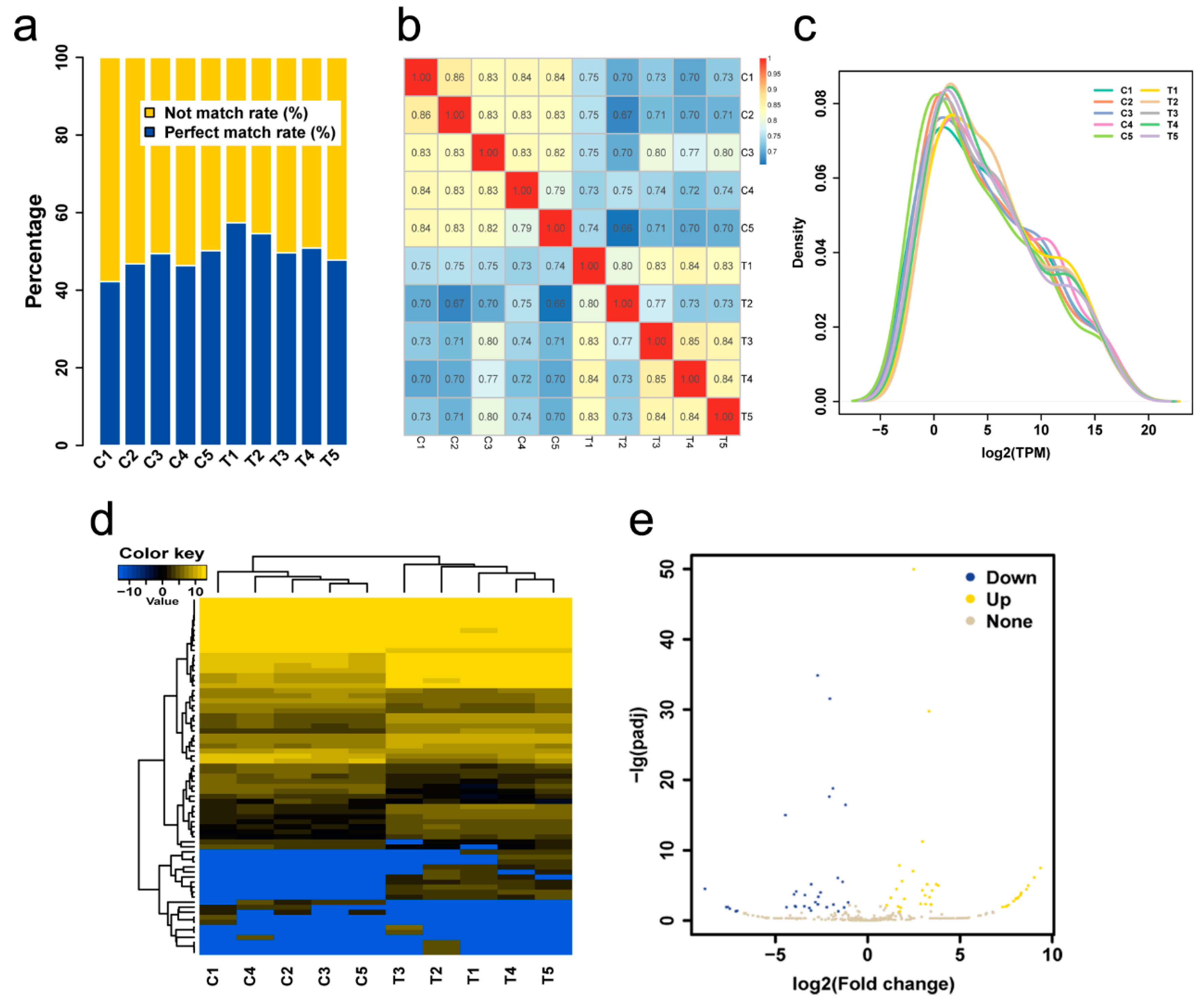
| Sample | Total Reads | Match Reads | Match Rate, % | Not Match Rate, % |
|---|---|---|---|---|
| C1 | 23,186,308 | 9,780,560 | 42.18 | 57.82 |
| C2 | 24,761,407 | 11,578,750 | 46.76 | 53.24 |
| C3 | 21,598,417 | 10,675,774 | 49.43 | 50.57 |
| C4 | 20,696,077 | 9,580,916 | 46.29 | 53.71 |
| C5 | 25,248,936 | 12,668,115 | 50.17 | 49.83 |
| T1 | 21,729,292 | 12,465,242 | 57.37 | 42.63 |
| T2 | 21,760,177 | 11,875,079 | 54.57 | 45.43 |
| T3 | 30,614,359 | 15,187,405 | 49.61 | 50.39 |
| T4 | 32,883,572 | 16,714,329 | 50.83 | 49.17 |
| T5 | 31,975,781 | 15,266,339 | 47.74 | 52.26 |
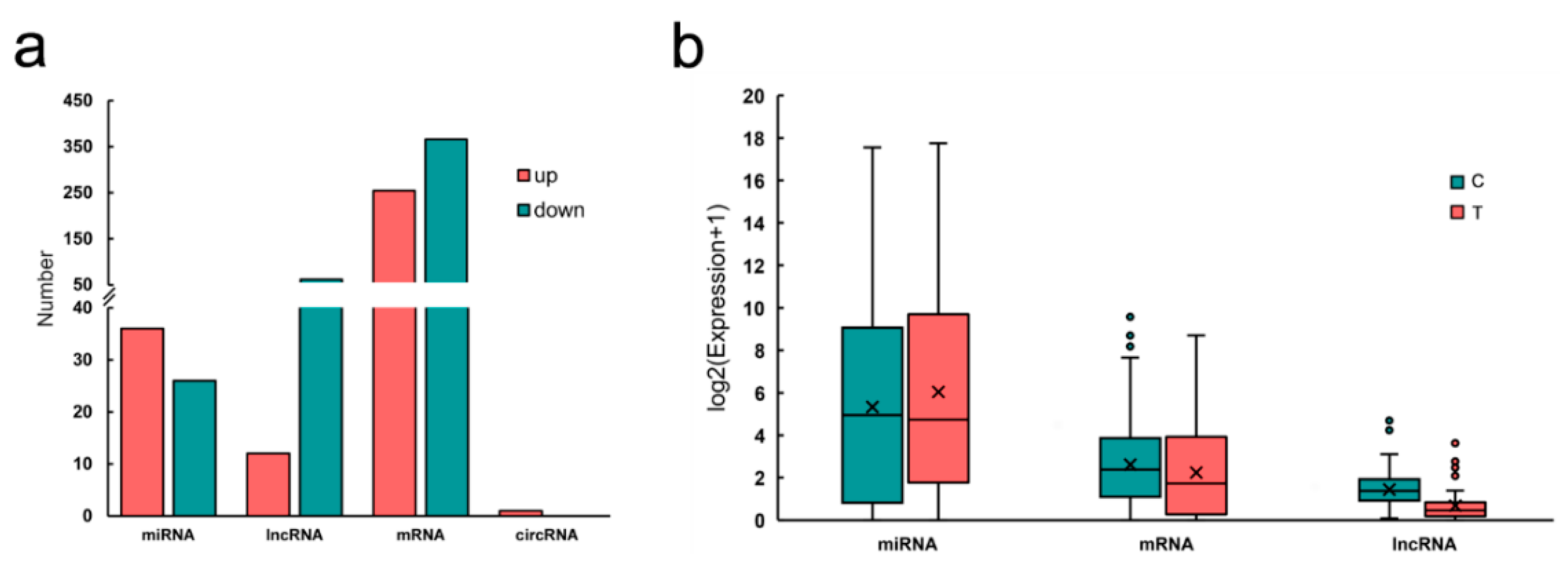
| Name | Log2 (FC) | p-Value | Label |
|---|---|---|---|
| cli-miR-7a-5p | 3.32 | 1.85 × 10−32 | up |
| cli-miR-20b-5p | 2.97 | 1.29 × 10−13 | up |
| cli-miR-21-5p | 2.49 | 2.86 × 10−53 | up |
| cli-miR-146b-5p | 1.72 | 3.66 × 10−10 | up |
| cli-miR-26-5p | −1.20 | 6.57 × 10−19 | down |
| cli-miR-99-5p | −1.88 | 2.08 × 10−21 | down |
| cli-miR-10a-5p | −2.05 | 2.14 × 10−34 | down |
| cli-miR-100-5p | −2.08 | 3.72 × 10−20 | down |
| cli-miR-125-5p | −2.71 | 7.37 × 10−38 | down |
| cli-miR-145-5p | −4.45 | 2.01 × 10−17 | down |
| Name | Betweenness Centrality | Closeness Centrality | Degree | Class |
|---|---|---|---|---|
| cli-miR-193-5p | 0.99 | 0.70 | 389 | miRNA |
| cli-miR-92-2-5p | 0.18 | 0.33 | 53 | miRNA |
| cli-miR-7b-5p | 0.93 | 0.77 | 14 | miRNA |
| cli-miR-214-5p | 0.05 | 0.30 | 13 | miRNA |
| cli-miR-106-5p | 0.06 | 0.30 | 10 | miRNA |
| Novel_189 | 0.09 | 0.58 | 22 | miRNA |
| Novel_188 | 0.09 | 0.58 | 22 | miRNA |
| Novel_187 | 0.09 | 0.58 | 22 | miRNA |
| cli-let-7c-5p | 1.00 | 1.00 | 21 | miRNA |
| TTC28 | 0.01 | 0.23 | 3 | mRNA |
| CREBRF | 0.03 | 0.42 | 2 | mRNA |
| SCML4 | 0.03 | 0.42 | 2 | mRNA |
| RAPGEF4 | 0.00 | 0.41 | 2 | mRNA |
| STRA6 | 0.02 | 0.45 | 2 | mRNA |
| LOC110358245 | 0.13 | 0.60 | 8 | mRNA |
| PLN | 0.02 | 0.50 | 7 | mRNA |
| UGCG | 0.02 | 0.50 | 7 | mRNA |
| DOT1L | 0.00 | 0.51 | 1 | mRNA |
| MSTRG.71575 | 0.01 | 0.23 | 3 | lncRNA |
| MSTRG.151245 | 0.01 | 0.23 | 3 | lncRNA |
| MSTRG.2421 | 0.01 | 0.23 | 3 | lncRNA |
| LOC110360772 | 0.00 | 0.16 | 2 | lncRNA |
| MSTRG.81107 | 0.14 | 0.53 | 2 | lncRNA |
| MSTRG.1547 | 0.14 | 0.53 | 2 | lncRNA |
| circ_0003020 | 0.00 | 0.51 | 1 | circRNA |
| miRNA | Label | mRNA | Label | lncRNA | Label |
|---|---|---|---|---|---|
| cli-miR-193-5p | up | CREBRF | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | ADRB1 | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | IHH | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | ADRB3 | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | CDH15 | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | PIK3CD | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | ITGB8 | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | HGF | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | PDGFRB | down | MSTRG.65211 | down |
| cli-miR-193-5p | up | CREBRF | down | LOC110355588 | down |
| cli-miR-193-5p | up | ADRB1 | down | LOC110355588 | down |
| cli-miR-193-5p | up | ADRB3 | down | LOC110355588 | down |
| cli-miR-193-5p | up | CDH15 | down | LOC110355588 | down |
| cli-miR-193-5p | up | PIK3CD | down | LOC110355588 | down |
| cli-miR-193-5p | up | ITGB8 | down | LOC110355588 | down |
| cli-miR-193-5p | up | HGF | down | LOC110355588 | down |
| cli-miR-193-5p | up | PDGFRB | down | LOC110355588 | down |
| cli-miR-193-5p | up | IHH | down | LOC110355588 | down |
| cli-miR-106-5p | up | CREBRF | down | MSTRG.71575 | down |
| cli-miR-106-5p | up | CREBRF | down | MSTRG.2421 | down |
| cli-miR-106-5p | up | CREBRF | down | MSTRG.151245 | down |
| cli-miR-338b-5p | down | RAPGEF4 | up | MSTRG.20439 | down |
| cli-miR-460b-5p | down | GRHL2 | up | MSTRG.132954 | down |
| cli-let-7c-5p | down | DOT1L | up | circ-0003020 | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, P.; Ma, H.; Li, Y.; Ni, A.; Isa, A.M.; Wang, P.; Bian, S.; Shi, L.; Zong, Y.; Wang, Y.; et al. Identification of microRNA-Associated-ceRNA Networks Regulating Crop Milk Production in Pigeon (Columba livia). Genes 2021, 12, 39. https://doi.org/10.3390/genes12010039
Ge P, Ma H, Li Y, Ni A, Isa AM, Wang P, Bian S, Shi L, Zong Y, Wang Y, et al. Identification of microRNA-Associated-ceRNA Networks Regulating Crop Milk Production in Pigeon (Columba livia). Genes. 2021; 12(1):39. https://doi.org/10.3390/genes12010039
Chicago/Turabian StyleGe, Pingzhuang, Hui Ma, Yunlei Li, Aixin Ni, Adamu Mani Isa, Panlin Wang, Shixiong Bian, Lei Shi, Yunhe Zong, Yuanmei Wang, and et al. 2021. "Identification of microRNA-Associated-ceRNA Networks Regulating Crop Milk Production in Pigeon (Columba livia)" Genes 12, no. 1: 39. https://doi.org/10.3390/genes12010039
APA StyleGe, P., Ma, H., Li, Y., Ni, A., Isa, A. M., Wang, P., Bian, S., Shi, L., Zong, Y., Wang, Y., Jiang, L., Hagos, H., Yuan, J., Sun, Y., & Chen, J. (2021). Identification of microRNA-Associated-ceRNA Networks Regulating Crop Milk Production in Pigeon (Columba livia). Genes, 12(1), 39. https://doi.org/10.3390/genes12010039