
PRKG2 Splice Site Variant in Dogo Argentino Dogs with Disproportionate Dwarfism

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
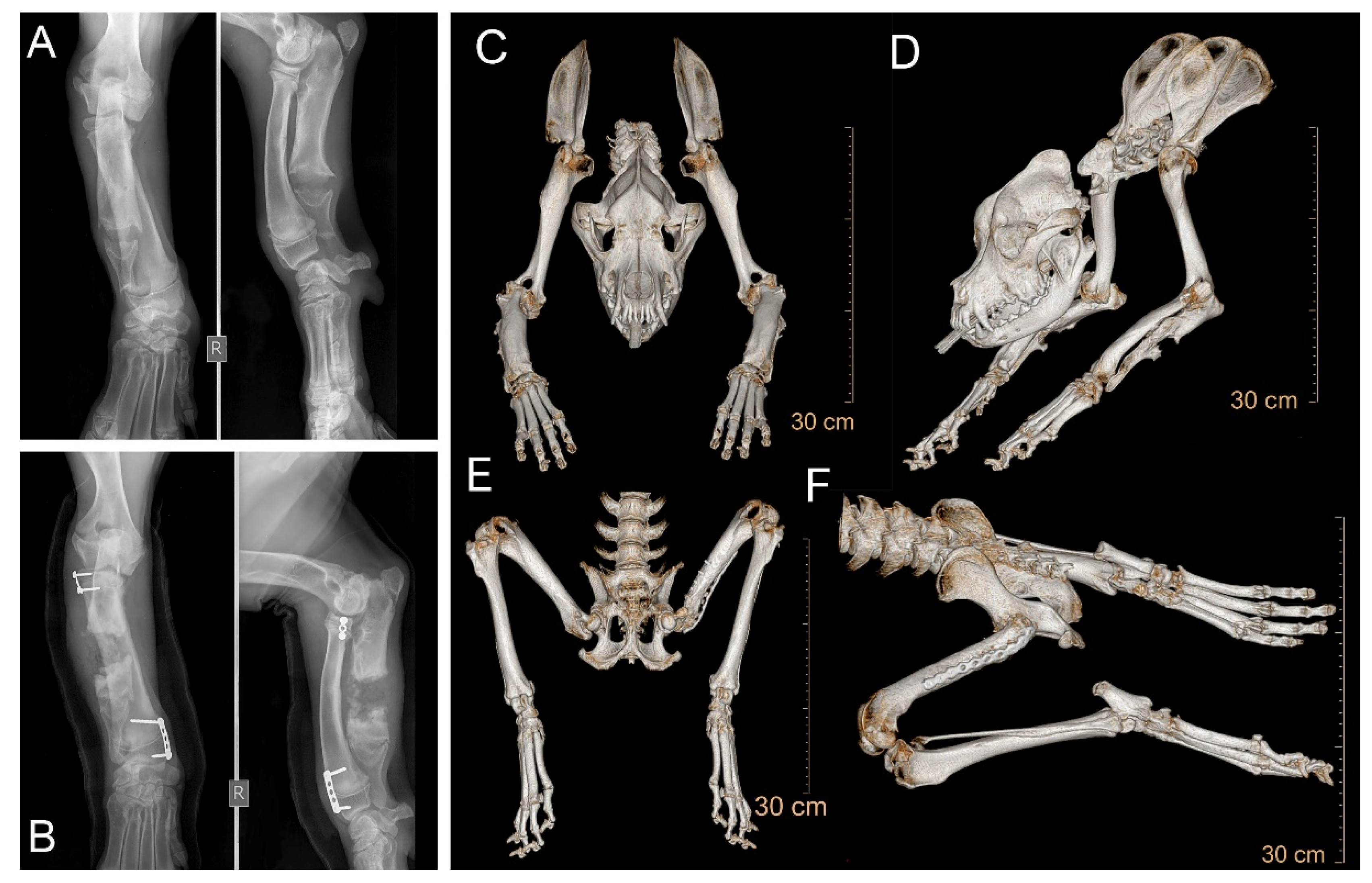
2.1. Diagnostic Imaging
2.2. DNA Isolation
2.3. Linkage and Homozygosity Mapping
2.4. Whole Genome Sequencing of an Affected Dog
2.5. Variant Calling
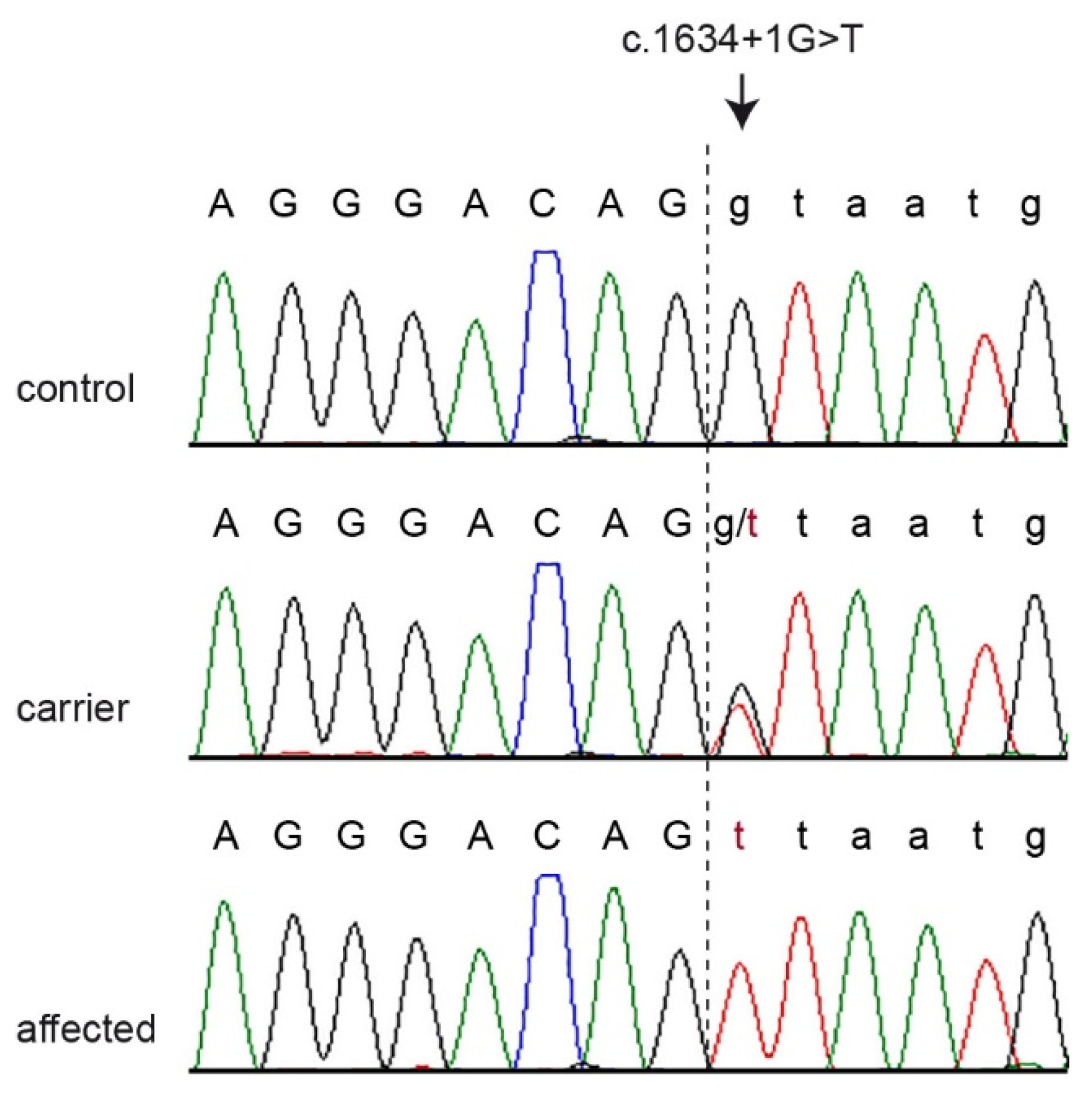
2.6. Sanger Sequencing
3. Results
3.1. Clinical Investigations and Phenotype Description
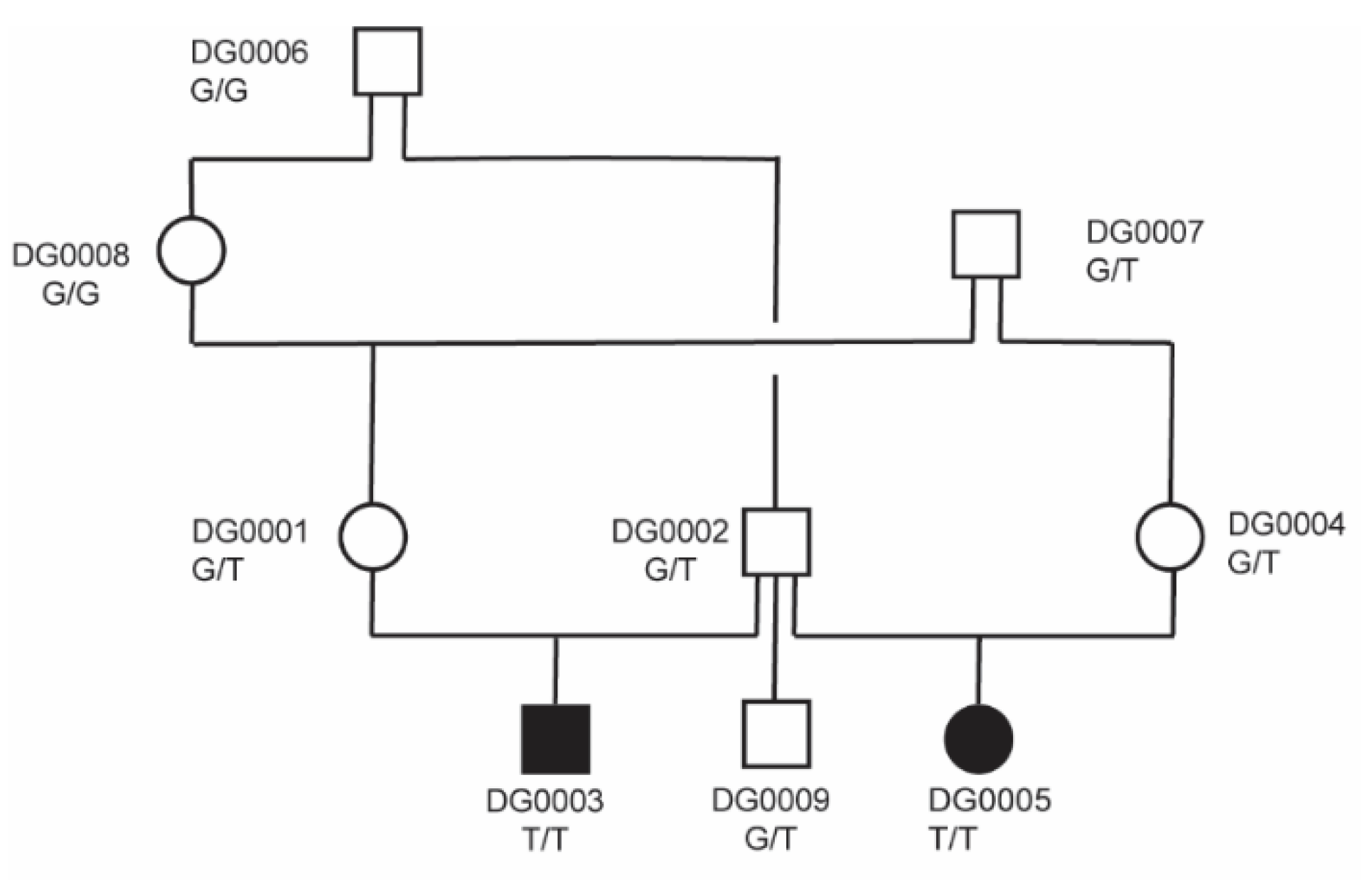
3.2. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krakow, D. Skeletal dysplasias. Clin. Perinatol. 2015, 42, 301–319. [Google Scholar] [CrossRef] [Green Version]
- Mortier, G.R.; Cohn, D.H.; Cormier-Daire, V.; Hall, C.; Krakow, D.; Mundlos, S.; Nishimura, G.; Robertson, S.; Sangiorgi, L.; Savarirayan, R.; et al. Nosology and classification of genetic skeletal disorders: 2019 revision. Am. J. Med. Genet. A 2019, 179, 2393–2419. [Google Scholar] [CrossRef]
- Sewell, M.D.; Chahal, A.; Al-Hadithy, N.; Blunn, G.W.; Molloy, S.; Hashemi-Nejad, A. Genetic skeletal dysplasias: A guide to diagnosis and management. J. Back Musculoskelet. Rehabil. 2015, 28, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Saber, A.Y. Dwarfism. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK563282/ (accessed on 16 October 2020).
- Deng, J.Z.; Hao, L.L.; Li, M.T.; Lang, S.; Zeng, Y.Z.; Liu, S.C.; Zhang, Y.L. Growth hormone and receptor gene mutations in Chinese Banna miniature pig. Anim. Cells Syst. 2011, 15, 310–314. [Google Scholar] [CrossRef]
- Rafati, N.; Andersson, L.S.; Mikko, S.; Feng, C.; Raudsepp, T.; Pettersson, J.; Janecka, J.; Wattle, O.; Ameur, A.; Thyreen, G.; et al. Large deletions at the SHOX locus in the pseudoautosomal region are associated with skeletal atavism in Shetland Ponies. G3 (Bethesda) 2016, 6, 2213–2223. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, J.A.; Tammen, I.; Windsor, P.A.; Bateman, J.F.; Savarirayan, R.; Nicholas, F.W.; Raadsma, H.W. Bulldog dwarfism in Dexter cattle is caused by mutations in ACAN. Mamm. Genome 2007, 18, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Bemji, M.N.; Isa, A.M.; Ibeagha-Awemu, E.M.; Wheto, M. Polymorphisms of caprine GnRHR gene and their association with litter size in West African Dwarf goats. Mol. Biol. Rep. 2018, 45, 63–69. [Google Scholar] [CrossRef]
- Boegheim, I.J.M.; Leegwater, P.A.J.; van Lith, H.A.; Back, W. Current insights into the molecular genetic basis of dwarfism in livestock. Vet. J. 2017, 224, 64–75. [Google Scholar] [CrossRef]
- Bannasch, D.L.; Baes, C.F.; Leeb, T. Genetic variants affecting skeletal morphology in domestic dogs. Trends Genet. 2020, 36, 598–609. [Google Scholar] [CrossRef]
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 retrogene on CFA12 is responsible for chondrodystrophy and intervertebral disc disease in dogs. Proc. Natl. Acad. Sci. USA 2017, 114, 11476–11481. [Google Scholar] [CrossRef] [Green Version]
- Kyöstilä, K.; Lappalainen, A.K.; Lohi, H. Canine chondrodysplasia caused by a truncating mutation in collagen-binding integrin alpha subunit 10. PLoS ONE 2013, 8, e75621. [Google Scholar] [CrossRef] [Green Version]
- Frischknecht, M.; Niehof-Oellers, H.; Jagannathan, V.; Owczarek-Lipska, M.; Drögemüller, C.; Dietschi, E.; Dolf, G.; Tellhelm, B.; Lang, J.; Tiira, K.; et al. A COL11A2 mutation in Labrador retrievers with mild disproportionate dwarfism. PLoS ONE 2013, 8, e60149. [Google Scholar] [CrossRef] [Green Version]
- Neff, M.W.; Beck, J.S.; Koeman, J.M.; Boguslawski, E.; Kefene, L.; Borgman, A.; Ruhe, A.L. Deletion of the sulfate transporter SLC13A1 is associated with an osteochondrodysplasia in the Miniature Poodle breed. PLoS ONE 2012, 7, e51917. [Google Scholar] [CrossRef] [PubMed]
- Willet, C.E.; Makara, M.; Reppas, G.; Tsoukalas, G.; Malik, R.; Haase, B.; Wade, C.M. Canine disorder mirrors human disease: Exonic deletion in HES7 causes autosomal recessive spondylocostal dysostosis in Miniature Schnauzer Dogs. PLoS ONE 2015, 10, e0117055. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abecasis, G.R.; Cherny, S.S.; Cookson, W.O.; Cardon, L.R. Merlin—Rapid analysis of dense genetic maps using sparse gene flow trees. Nat. Genet. 2002, 30, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, V.; Drögemüller, C.; Leeb, T. Dog Biomedical Variant Database Consortium (DBVDC). A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Dogo Argentino-Breeders and Kennels-EuroBreeder.com. Available online: https://www.eurobreeder.com/breeds/dogo_argentino.html (accessed on 18 May 2021).
- Orstavik, S.; Solberg, R.; Taskén, K.; Nordahl, M.; Altherr, M.R.; Hansson, V.; Jahnsen, T.; Sandberg, M. Molecular cloning, cDNA structure, and chromosomal localization of the human type II cGMP-dependent protein kinase. Biochem. Biophys. Res. Commun. 1996, 220, 759–765. [Google Scholar] [CrossRef]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, K.; Uehara, S.; Nagahama, M.; Tsuji, A. Transcriptional regulation of cGMP-dependent protein kinase II (cGK-II) in chondrocytes. Biosci. Biotechnol. Biochem. 2010, 74, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, H.; Chaboissier, M.-C.; Martin, J.F.; Schedl, A.; de Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koltes, J.E.; Kumar, D.; Kataria, R.S.; Cooper, V.; Reecy, J.M. Transcriptional profiling of PRKG2-null growth plate identifies putative down-stream targets of PRKG2. BMC Res. Notes 2015, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Chikuda, H.; Kugimiya, F.; Hoshi, K.; Ikeda, T.; Ogasawara, T.; Shimoaka, T.; Kawano, H.; Kamekura, S.; Tsuchida, A.; Yokoi, N.; et al. Cyclic GMP-dependent protein kinase II is a molecular switch from proliferation to hypertrophic differentiation of chondrocytes. Genes Dev. 2004, 18, 2418–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-González, F.; Wadhwa, S.; Rodriguez-Zabala, M.; Kumar, S.; Aza-Carmona, M.; Sentchordi-Montané, L.; Alonso, M.; Ahmad, I.; Zahra, S.; Kumar, D.; et al. Biallelic cGMP-dependent type II protein kinase gene (PRKG2) variants cause a novel acromesomelic dysplasia. J. Med. Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Koltes, J.E.; Mishra, B.P.; Kumar, D.; Kataria, R.S.; Totir, L.R.; Fernando, R.L.; Cobbold, R.; Steffen, D.; Coppieters, W.; Georges, M.; et al. A nonsense mutation in cGMP-dependent type II protein kinase (PRKG2) causes dwarfism in American Angus cattle. Proc. Natl. Acad. Sci. USA 2009, 106, 19250–19255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, A.; Aszódi, A.; Seidler, U.; Ruth, P.; Hofmann, F.; Fässler, R. Intestinal secretory defects and dwarfism in mice lacking cGMP-dependent protein kinase II. Science 1996, 274, 2082–2086. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, T.; Ogawa, Y.; Chusho, H.; Yasoda, A.; Tamura, N.; Komatsu, Y.; Pfeifer, A.; Hofmann, F.; Nakao, K. Cyclic GMP-dependent protein kinase II plays a critical role in C-type natriuretic peptide-mediated endochondral ossification. Endocrinology 2002, 143, 3604–3610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragicevich, C.J.; Jones, J.C.; Bridges, W.; Dunn, H. Computed tomographic measures of funnel-shaped lumbar vertebral canal and articular process dysplasia malformations differ between German Shepherd and Belgian Malinois military working dogs. Front. Vet. Sci. 2020, 7, 275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Filtering Step | Variants |
|---|---|
| Homozygous variants in whole genome | 2,625,704 |
| Private homozygous variants (absent from 795 control genomes) | 2007 |
| Private homozygous variants in 125 Mb critical intervals | 196 |
| Protein-changing private variants in critical intervals | 3 |
| Chr. | Position | Ref. | Alt. | Gene | HGVS-c | HGVS-p |
|---|---|---|---|---|---|---|
| 12 | 1,411,804 | C | A | NELFE | c.7G>T | p.Val3Leu |
| 23 | 50,457,119 | G | C | LEKR1 | c.206G>C | p.Arg69Thr |
| 32 | 5,299,068 | C | A | PRKG2 | c.1634+1G>T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudd Garces, G.; Turba, M.E.; Muracchini, M.; Diana, A.; Jagannathan, V.; Gentilini, F.; Leeb, T. PRKG2 Splice Site Variant in Dogo Argentino Dogs with Disproportionate Dwarfism. Genes 2021, 12, 1489. https://doi.org/10.3390/genes12101489
Rudd Garces G, Turba ME, Muracchini M, Diana A, Jagannathan V, Gentilini F, Leeb T. PRKG2 Splice Site Variant in Dogo Argentino Dogs with Disproportionate Dwarfism. Genes. 2021; 12(10):1489. https://doi.org/10.3390/genes12101489
Chicago/Turabian StyleRudd Garces, Gabriela, Maria Elena Turba, Myriam Muracchini, Alessia Diana, Vidhya Jagannathan, Fabio Gentilini, and Tosso Leeb. 2021. "PRKG2 Splice Site Variant in Dogo Argentino Dogs with Disproportionate Dwarfism" Genes 12, no. 10: 1489. https://doi.org/10.3390/genes12101489
APA StyleRudd Garces, G., Turba, M. E., Muracchini, M., Diana, A., Jagannathan, V., Gentilini, F., & Leeb, T. (2021). PRKG2 Splice Site Variant in Dogo Argentino Dogs with Disproportionate Dwarfism. Genes, 12(10), 1489. https://doi.org/10.3390/genes12101489