
A Single Base Insertion in F9 Causing Hemophilia B in a Family of Newfoundland–Parti Standard Poodle Hybrid Dogs

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dogs and Samples
2.2. Molecular Genetic Analysis
2.3. Genotyping
2.4. Family Study
3. Results
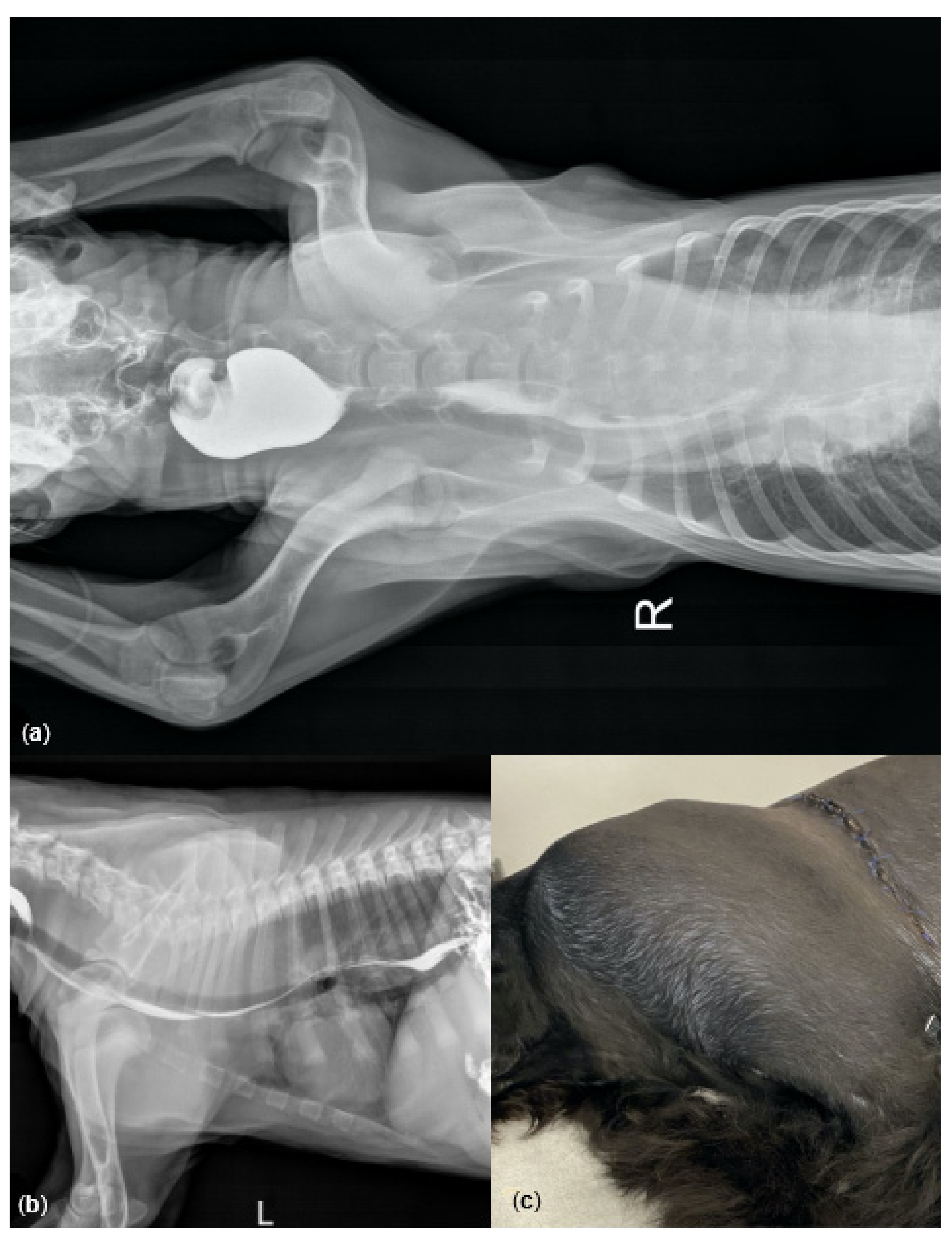
3.1. Case Report
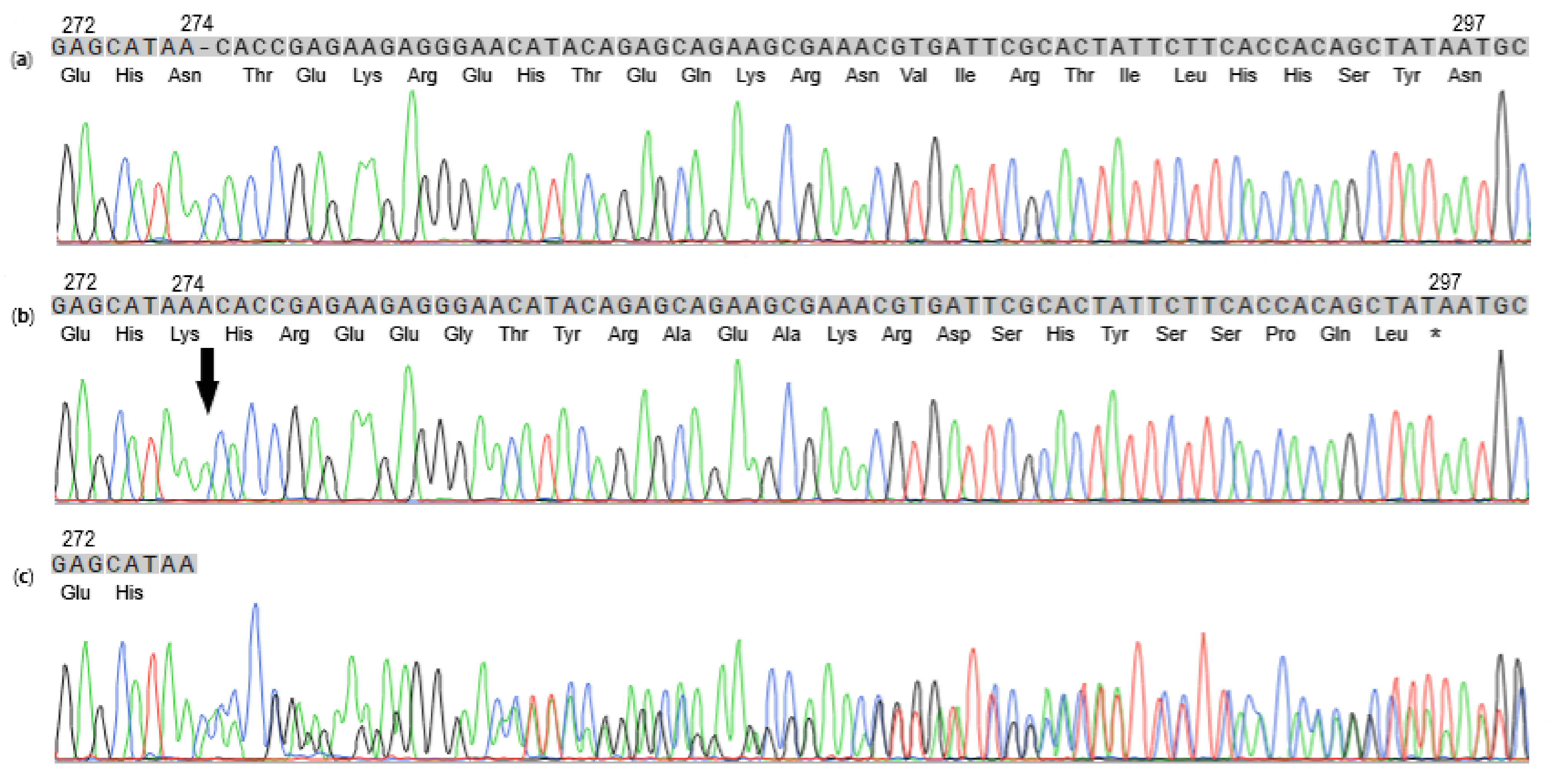
3.2. Molecular Genetic Analysis
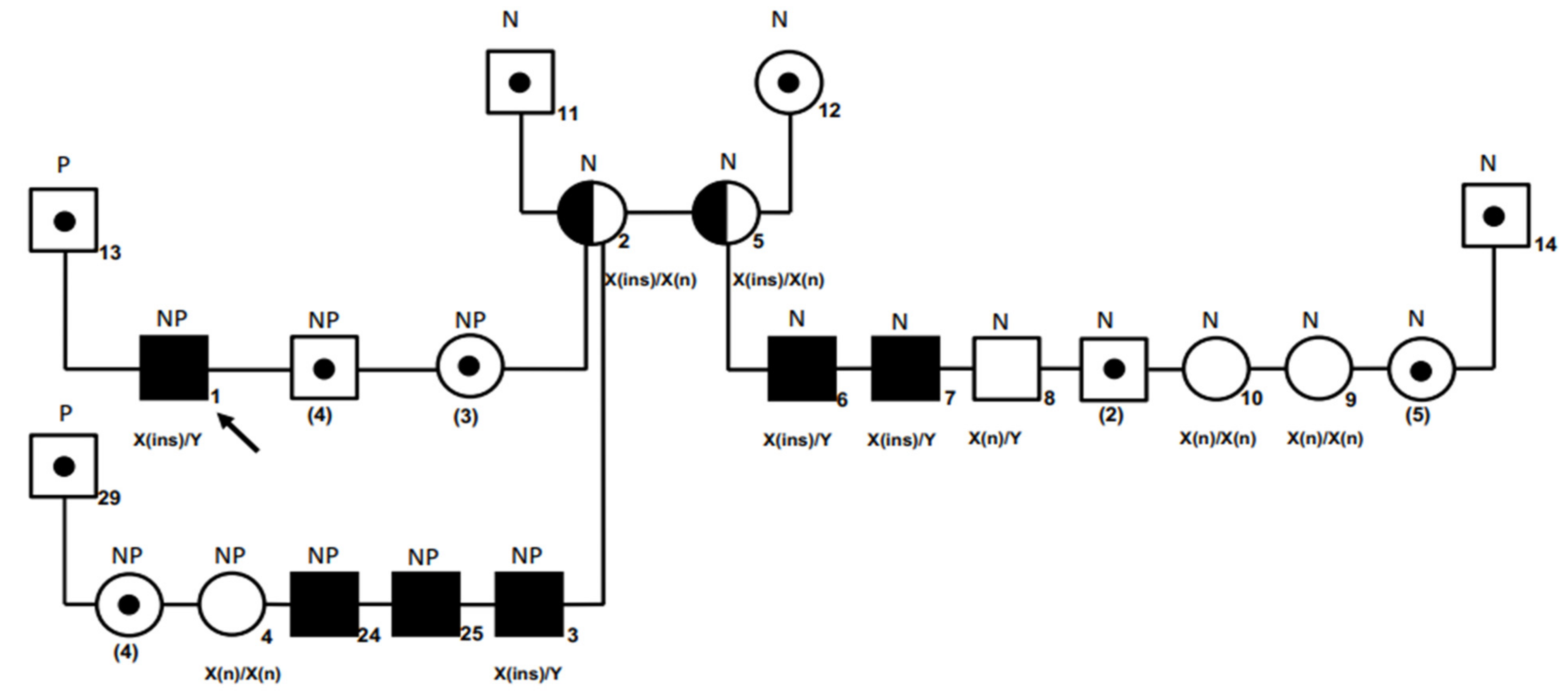
3.3. Family and Breed Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolton-Maggs, P.H.B.; Pasi, K.J. Haemophilias A and B. Lancet Lond. Engl. 2003, 361, 1801–1809. [Google Scholar] [CrossRef]
- Lozier, J.N.; Nichols, T.C. Animal Models of Hemophilia and Related Bleeding Disorders. Semin. Hematol. 2013, 50, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, M. A Review of Canine Inherited Bleeding Disorders: Biochemical and Molecular Strategies for Disease Characterization and Carrier Detection. J. Hered. 1999, 90, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Mischke, R. Hämophilie A und B beim Hund. Tierärztl. Prax. Ausg. K Kleintiere Heimtiere 2012, 40, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Slappendel, R.J. Congenital (Hereditary) Clotting Disorders in the Dog. Vet. Q. 1998, 20 (Suppl. 1), S41–S43. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Wardrop, K.J. Schalm´s Veterinary Hematology, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 661–667, Chapter 86; ISBN 978-0-8138-0896-3. [Google Scholar]
- Evans, J.P.; Brinkhous, K.M.; Brayer, G.D.; Reisner, H.M.; High, K.A. Canine Hemophilia B Resulting from a Point Mutation with Unusual Consequences. Proc. Natl. Acad. Sci. USA 1989, 86, 10095–10099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauser, A.E.; Whitlark, J.; Whitney, K.M.; Lothorp, C.D.J. A Deletion Mutation Causes Hemophilia B in Lhasa Apso Dogs. Blood 1996, 88, 3451–3455. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.B.; Gu, W. Complete Deletion of Factor IX Gene and Inhibition of Factor IX Activity in a Labrador Retriever with Hemophilia B. J. Am. Vet. Med. Assoc. 1997, 211, 1418–1421. [Google Scholar]
- Gu, W.; Brooks, M.; Catalfamo, J.; Ray, J.; Ray, K. Two Distinct Mutations Cause Severe Hemophilia B in Two Unrelated Canine Pedigrees. Thromb. Haemost. 1999, 82, 1270–1275. [Google Scholar] [CrossRef]
- Brooks, M.B.; Gu, W.; Barnas, J.L.; Ray, J.; Ray, K. A Line 1 Insertion in the Factor IX Gene Segregates with Mild Hemophilia B in Dogs. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2003, 14, 788–795. [Google Scholar] [CrossRef]
- Mischke, R.; Kühnlein, P.; Kehl, A.; Langbein-Detsch, I.; Steudle, F.; Schmid, A.; Dandekar, T.; Czwalinna, A.; Müller, E. G244E in the Canine Factor IX Gene Leads to Severe Haemophilia B in Rhodesian Ridgebacks. Vet. J. Lond. Engl. 1997 2011, 187, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Brenig, B.; Steingräber, L.; Shan, S.; Xu, F.; Hirschfeld, M.; Andag, R.; Spengeler, M.; Dietschi, E.; Mischke, R.; Leeb, T. Christmas Disease in a Hovawart Family Resembling Human Hemophilia B Leyden Is Caused by a Single Nucleotide Deletion in a Highly Conserved Transcription Factor Binding Site of the F9 Gene Promoter. Haematologica 2019, 104, 2307–2313. [Google Scholar] [CrossRef] [Green Version]
- Online Mendelian Inheritance in Animals, OMIA. Sydney School of Veterinary Science. Available online: Https://Omia.Org/ (accessed on 21 August 2021).
- O’Leary, N.A.; Wright, M.W.; Brister, R.J.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference Sequence (RefSeq) Database at NCBI: Current Status, Taxonomic Expansion, and Functional Annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- SeqScanner; Applied Biosystems/Thermo Fisher Scientific. Available online: https://www.thermofisher.cn/cn/zh/home/life-science/sequencing/sanger-sequencing/sanger-dna-sequencing/sanger-sequencing-data-analysis.html (accessed on 20 September 2021).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jagannathan, V.; Drögemüller, C.; Leeb, T. Dog Biomedical Variant Database Consortium (DBVC) A Comprehensive Biomedical Variant Catalogue Based on Whole Genome Sequences of 582 Dogs and Eight Wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Delphine, B.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Wisdom PanelTM|World’s Leading DNA Test Service for Dogs and Cats. Available online: https://www.wisdompanel.com/en-us/ (accessed on 23 August 2021).
- Vetnostic Laboratories®. Available online: https://www.vetnostic.com/ (accessed on 23 August 2021).
- Russell, K.E.; Olsen, E.H.N.; Raymer, R.A.; Merricks, E.P.; Bellinger, D.A.; Read, M.S.; Rup, B.J.; Keith Jr, J.C.; McCarthy, K.P.; Schaub, R.G.; et al. Reduced Bleeding Events with Subcutaneous Administration of Recombinant Human Factor IX in Immune-Tolerant Hemophilia B Dogs. Blood 2003, 102, 4393–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Dunnen, J.T.; Dalgleish, R.; Maglott, D.R.; Hart, R.K.; Greenblatt, M.S.; McGowan-Jordan, J.; Roux, A.-F.; Smith, T.; Antonarakis, S.E.; Taschner, P.E.M. HGVS Recommendations for the Description of Sequence Variants: 2016 Update. Hum. Mutat. 2016, 37, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.E.; Bajaj, S.P. Structure–Function Relationships in Factor IX and Factor IXa. Trends Cardiovasc. Med. 2003, 13, 39–45. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, Y.K. When a Ribosome Encounters a Premature Termination Codon. BMB Rep. 2013, 46, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Wulff, K.; Schröder, W.; Wehnert, M.; Herrmann, F.H. Twenty-Five Novel Mutations of the Factor IX Gene in Haemophilia B. Hum. Mutat. 1995, 6, 346–348. [Google Scholar] [CrossRef]
- Rallapalli, P.M.; Kemball-Cook, G.; Tuddenham, E.G.; Gomez, K.; Perkins, S.J. An Interactive Mutation Database for Human Coagulation Factor IX Provides Novel Insights into the Phenotypes and Genetics of Hemophilia B. J. Thromb. Haemost. JTH 2013, 11, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- McVey, J.H.; Rallapalli, P.M.; Kemball-Cook, G.; Hampshire, D.J.; Giansily-Blaizot, M.; Gomez, K.; Perkins, S.J.; Ludlam, C.A. The European Association for Haemophilia and Allied Disorders (EAHAD) Coagulation Factor Variant Databases: Important Resources for Haemostasis Clinicians and Researchers. Haemophilia 2020, 26, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Nazeef, M.; Sheehan, J.P. New Developments in the Management of Moderate-to-Severe Hemophilia B. J. Blood Med. 2016, 7, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, T.C.; Hough, C.; Agerso, H.; Ezban, M.; Lillicrap, D. Canine Models of Inherited Bleeding Disorders in the Development of Coagulation Assays, Novel Protein Replacement and Gene Therapies. J. Thromb. Haemost. 2016, 14, 894–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchini, M.; Mannucci, P.M. The History of Hemophilia. Semin. Thromb. Hemost. 2014, 40, 571–576. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Day | Time of Day | PCV (%)/ TP (g/dL) | aPTT/ PT (s) * | Comments |
|---|---|---|---|---|
| 1 | 17 | 30/ND | 240/11 | Admission, dyspneic |
| 2 | 08 | 13/5.0 | >300/11 | Progressive dyspnea, ventilated |
| 10 | ND/ND | 162/ND | Post pRBC transfusions ** | |
| 14 | 18/5.0 | 133/ND | Post thoracic surgery and FFP transfusion | |
| 23 | ND/ND | 135/ND | Post additional FFP transfusion | |
| 3 | 07 | 20/6.0 | ND/ND | Clinically stable |
| 4 | 14 | 24/5.0 | 126/11 | Clinically stable |
| 5 | 10 | 35/7.0 | 124/ND | Prior to FFP transfusion ** |
| 22 | 34/8.0 | 129/ND | Post FFP transfusion | |
| 6 | 10 | ND/ND | 105/ND | Post FFP transfusion |
| 10 | 08 | 41/8.0 | >300/ND | Severe chest wall hematoma |
| 11 | 10 | 20/6.0 | ND/ND | Euthanasia |
| Reference Ranges | 30-45/5.5-7.2 | 72-102/11-17 | ||
| Hemostatic Parameters | Unit | Results | Reference Range | |
|---|---|---|---|---|
| Day 2 | Day 10 | |||
| PT | s | 11.9 | ND | 11.0–15.5 |
| aPTT | s | 25.0 | ND | 8.5–15.5 |
| TCT | s | 4.5 | ND | 5.0–9.0 |
| FVII | % | 123 | 100 | 50–150 |
| FVIII | % | 104 | 51 | 50–200 |
| FIX | % | 11 | 3 | 50–150 |
| FX | % | 166 | 106 | 80–175 |
| FXI | % | 77 | 56 | 60–150 |
| FXII | % | 84 | 64 | 60–150 |
| vWF | % | 141 | ND | 70–180 |
| FIX Inhibitor | BU/mL | ND | 0.15 | 0–1.00 |
| Breed a | Position in Genome | cDNA & Amino Acid Position | Type of Mutation | FIX Activity (%) b | aPTT (s) c | Clinical Signs | Occurrence |
|---|---|---|---|---|---|---|---|
| Cairn Terrier [7,22] | g.109532018G>A Exon 8 | c.1253G>A p.Gly418Glu * | missense | 0 | 70–90 | Hemarthrosis, hematoma, CNS bleeding, mediastinal & cavity bleeding | Research Colony |
| Lhasa Apso [8] | g.109521356_109521361delinsT Intron 5 | c.548_553delinsT p.Arg183LeufsTer3 | deletion, insertion | <1 | 48/60 | Hemorrhage, not specified | Single cases |
| Labrador Retriever [9] | complete deletion of factor IX gene | - | deletion | 1 | >60 | Hematoma, hemarthrosis, prolonged bleeding from minor wounds | Family |
| Pit Bull Terrier [10] | deletion of entire 5´region extending to exon 6 | - | deletion | <1 | 86–120 | Hemarthrosis, intramuscular hematoma, epistaxis | Single case |
| Airedale Terrier [10] | g.109532012_13ins(5 kb) Exon 8 | c.1247_1248ins(5kb) | insertion | <1 | 86–120 | Hemarthrosis, intramuscular hematoma, epistaxis | Single case |
| German Wirehaired Pointer [11] | g.109521130_109521131insLINE1 Intron 5 | - | insertion | 2.4–6.4 | Prolonged d | Hematoma, hemarthrosis, excessive bleeding after surgery & minor wounds | several cases in Northern America & Europe |
| Rhodesian Ridgeback [12] | g.109530847G>A Exon 7 | c.731G>A p.Gly237Glu | missense | ∼1 | Prolonged d | Spontaneous hemorrhage & severe bleeding after minor surgeries | Family |
| Hovawart [13] | g.109501492delC Promoter | - | missense | 2–5 | 3–5 fold increase | Umbilical & gingival bleeding, hemarthrosis, hematomas & hemorrhage after surgery & vaccination | Family |
| Newfoundland & Standard Poodle hybrid dogs * | g.109531586_109531587insA Exon 8 | c.821_822insA p.Asn274LysfsTer23 | insertion | 11/3 | 25 e | Hemarthrosis, hematoma, hemorrhage when losing deciduous teeth & after vaccination | Family |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuder, H.; Sandzhieva-Vuzzo, L.; Kehl, A.; Rappaport, J.M.; Müller, E.; Giger, U. A Single Base Insertion in F9 Causing Hemophilia B in a Family of Newfoundland–Parti Standard Poodle Hybrid Dogs. Genes 2021, 12, 1491. https://doi.org/10.3390/genes12101491
Kuder H, Sandzhieva-Vuzzo L, Kehl A, Rappaport JM, Müller E, Giger U. A Single Base Insertion in F9 Causing Hemophilia B in a Family of Newfoundland–Parti Standard Poodle Hybrid Dogs. Genes. 2021; 12(10):1491. https://doi.org/10.3390/genes12101491
Chicago/Turabian StyleKuder, Henrike, Liubov Sandzhieva-Vuzzo, Alexandra Kehl, Jonathan M. Rappaport, Elisabeth Müller, and Urs Giger. 2021. "A Single Base Insertion in F9 Causing Hemophilia B in a Family of Newfoundland–Parti Standard Poodle Hybrid Dogs" Genes 12, no. 10: 1491. https://doi.org/10.3390/genes12101491
APA StyleKuder, H., Sandzhieva-Vuzzo, L., Kehl, A., Rappaport, J. M., Müller, E., & Giger, U. (2021). A Single Base Insertion in F9 Causing Hemophilia B in a Family of Newfoundland–Parti Standard Poodle Hybrid Dogs. Genes, 12(10), 1491. https://doi.org/10.3390/genes12101491