
Association of CYP26C1 Promoter Hypomethylation with Small Vessel Occlusion in Korean Subjects


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. DNA Methylation Analysis
2.3. Prediction of Transcription Factors in CpG Sites
2.4. Statistical Analysis
2.5. Availability of Data and Materials
2.6. Consent to Participate and Ethics Approval
3. Results
3.1. Characteristics of Subjects
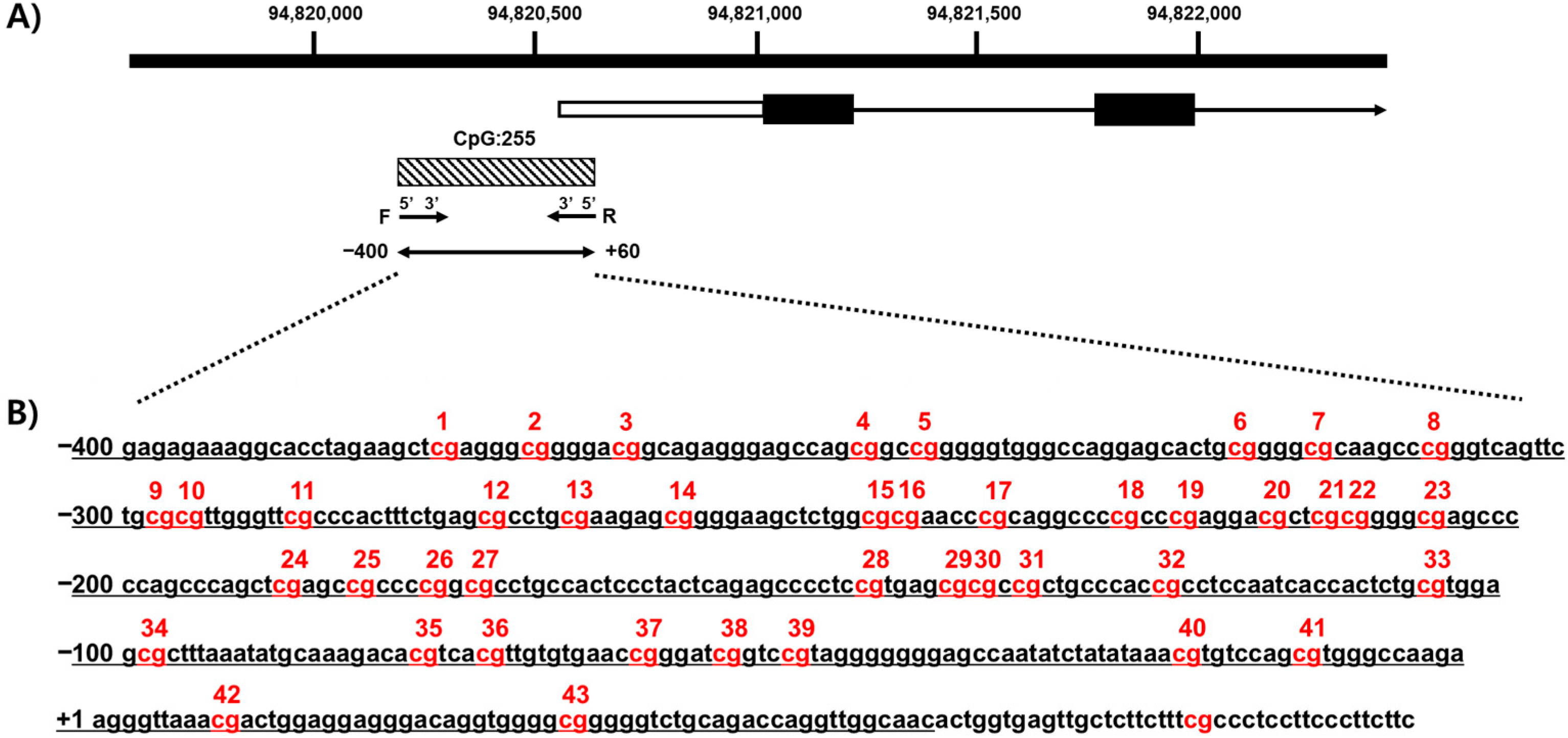
3.2. Map of CpG Islands in the CYP26C1 Promoter Region
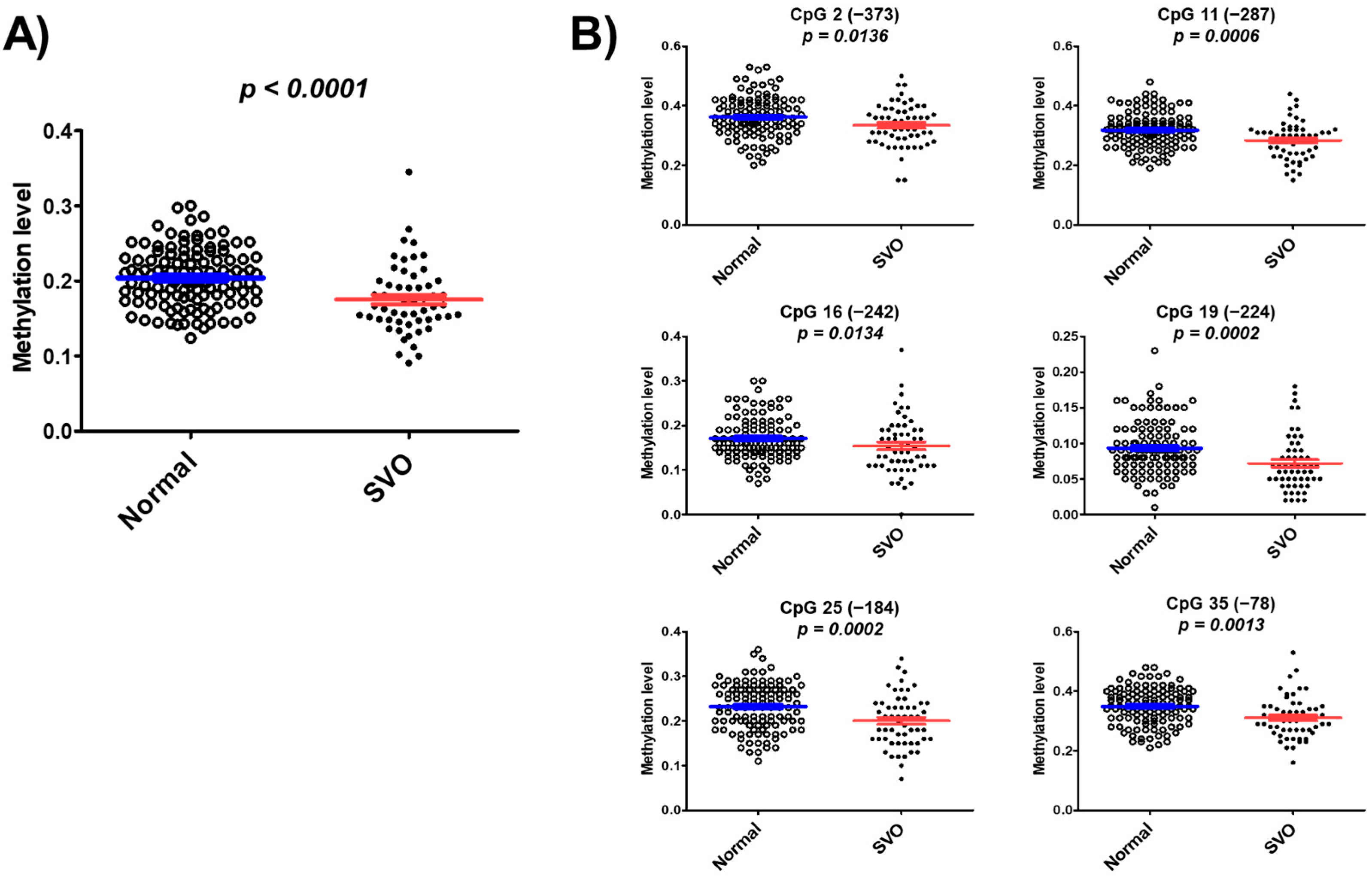
3.3. CYP26C1 Promoter Methylation in Normal Subjects and in Patients with SVO
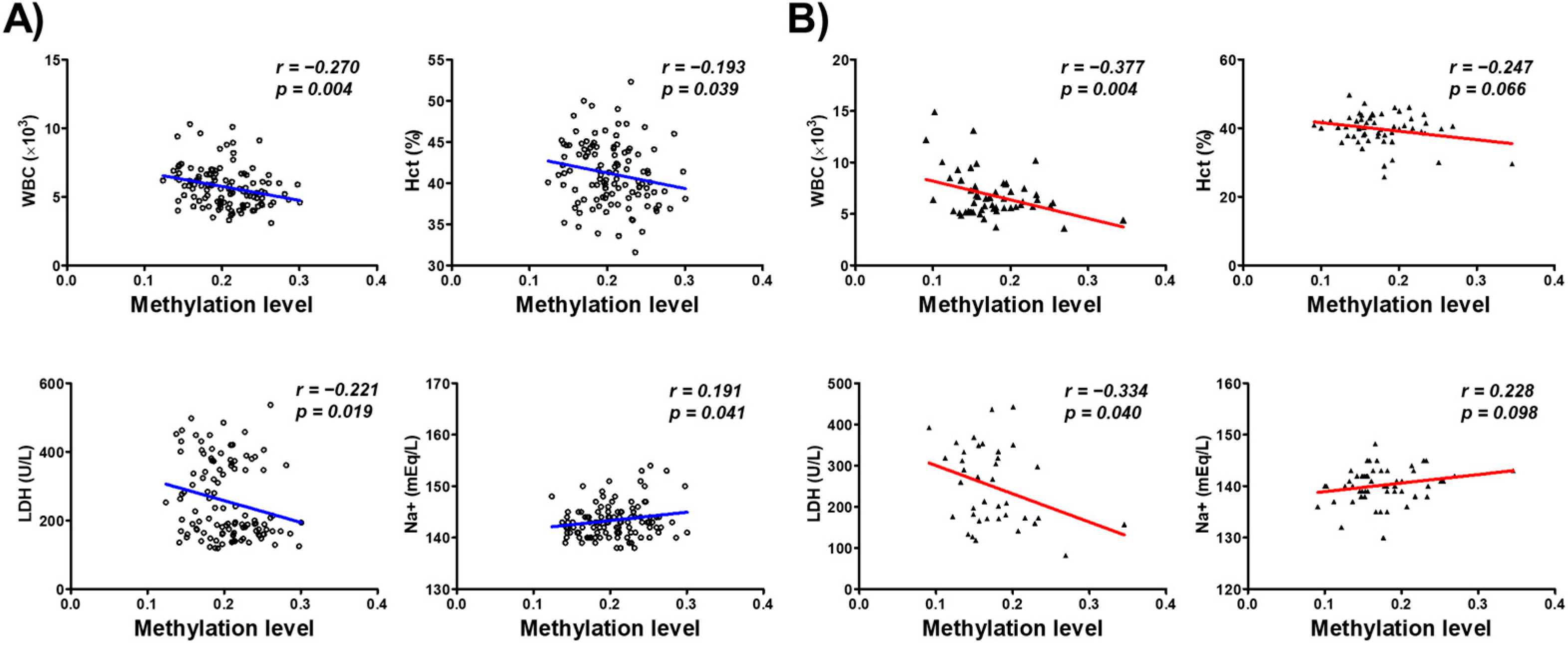
3.4. Association of CYP26C1 Promoter Methylation with Blood Parameters
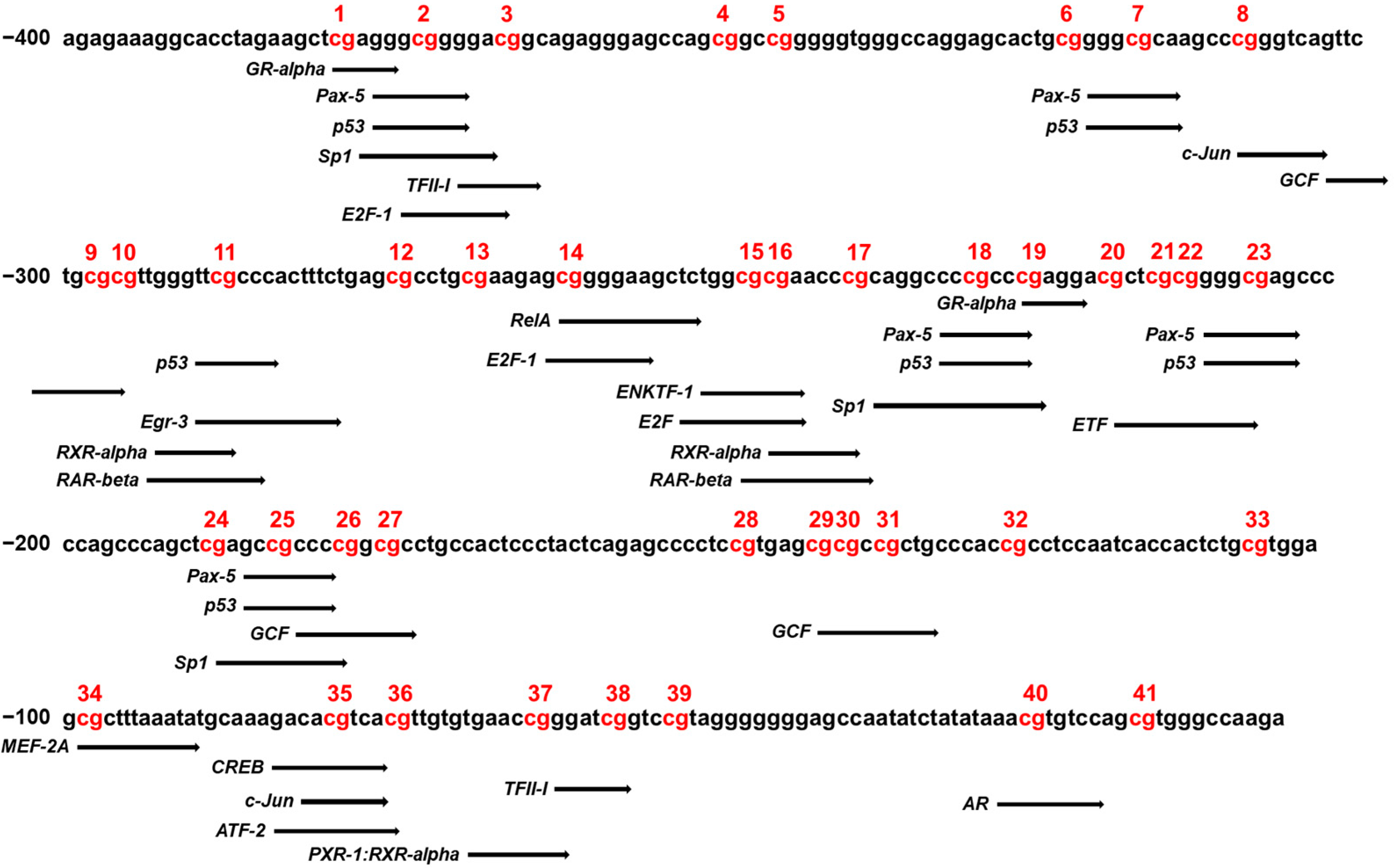
3.5. Binding Sites of Transcription Factors at CpG Sites in the CYP26C1 Promoter Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.Y.; Kang, K.; Kang, J.; Koo, J.; Kim, D.H.; Kim, B.J.; Kim, W.J.; Kim, E.G.; Kim, J.G.; Kim, J.M.; et al. Executive Summary of Stroke Statistics in Korea 2018: A Report from the Epidemiology Research Council of the Korean Stroke Society. J. Stroke 2019, 21, 42–59. [Google Scholar] [CrossRef]
- Johnson, C.O.; Nguyen, M.; Roth, G.A.; Nichols, E.; Alam, T.; Abate, D.; Abd-Allah, F.; Abdelalim., A.; Abraha, H.N.; Abu-Rmeileh, N.M.E.; et al. Global, regional, and national burden of stroke, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 439–458. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.P.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.J.; Kim, J.S. Ischemic stroke subtype classification: An asian viewpoint. J. Stroke 2014, 16, 8–17. [Google Scholar] [CrossRef]
- Bjerkreim, A.T.; Khanevski, A.N.; Thomassen, L.; Selvik, H.A.; Waje-Andreassen, U.; Naess, H.; Logallo, N. Five-year readmission and mortality differ by ischemic stroke subtype. J. Neurol. Sci. 2019, 403, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.H.; Lin, J.R.; Liou, C.W.; Lee, J.D.; Peng, T.I.; Lee, M.; Lee, T.H. Causes of Death in Different Subtypes of Ischemic and Hemorrhagic Stroke. Angiology 2018, 69, 582–590. [Google Scholar] [CrossRef]
- Kang, K.; Park, T.H.; Kim, N.; Jang, M.U.; Park, S.S.; Park, J.M.; Ko, Y.; Lee, S.; Lee, K.B.; Lee, J.; et al. Recurrent Stroke, Myocardial Infarction, and Major Vascular Events during the First Year after Acute Ischemic Stroke: The Multicenter Prospective Observational Study about Recurrence and Its Determinants after Acute Ischemic Stroke I. J. Stroke Cerebrovasc. Dis. 2016, 25, 656–664. [Google Scholar] [CrossRef]
- Lawrence, A.J.; Patel, B.; Morris, R.G.; MacKinnon, A.D.; Rich, P.M.; Barrick, T.R.; Markus, H.S. Mechanisms of cognitive impairment in cerebral small vessel disease: Multimodal MRI results from the St George’s cognition and neuroimaging in stroke (SCANS) study. PLoS ONE 2013, 8, e61014. [Google Scholar] [CrossRef]
- Liu, Y.; Braidy, N.; Poljak, A.; Chan, D.K.Y.; Sachdev, P. Cerebral small vessel disease and the risk of Alzheimer’s disease: A systematic review. Ageing Res. Rev. 2018, 47, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Jee, S.H.; Park, J.W.; Lee, S.Y.; Nam, B.H.; Ryu, H.G.; Kim, S.Y.; Kim, Y.N.; Lee, J.K.; Choi, S.M.; Yun, J.E. Stroke risk prediction model: A risk profile from the Korean study. Atherosclerosis 2008, 197, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S. Stroke Risk Factors, Genetics, and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef] [PubMed]
- Boden-Albala, B.; Sacco, R.L. Lifestyle factors and stroke risk: Exercise, alcohol, diet, obesity, smoking, drug use, and stress. Curr. Atheroscler. Rep. 2000, 2, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Picascia, A.; Grimaldi, V.; Iannone, C.; Soricelli, A.; Napoli, C. Innate and adaptive immune response in stroke: Focus on epigenetic regulation. J. Neuroimmunol. 2015, 289, 111–120. [Google Scholar] [CrossRef]
- Zeng, M.; Zhen, J.; Zheng, X.; Qiu, H.; Xu, X.; Wu, J.; Lin, Z.; Hu, J. The Role of DNA Methylation in Ischemic Stroke: A Systematic Review. Front. Neurol. 2020, 11, 566124. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Long, T.I.; Arakawa, K.; Wang, R.; Yu, M.C.; Laird, P.W. DNA methylation as a biomarker for cardiovascular disease risk. PLoS ONE 2010, 5, e9692. [Google Scholar] [CrossRef] [Green Version]
- Taimi, M.; Helvig, C.; Wisniewski, J.; Ramshaw, H.; White, J.; Amad, M.; Korczak, B.; Petkovich, M. A novel human cytochrome P450, CYP26C1, involved in metabolism of 9-cis and all-trans isomers of retinoic acid. J. Biol. Chem. 2004, 279, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Wang, J.; Hu, M.; Chen, X.; Lu, Z.; Bellanti, J.A.; Zheng, S.G. All trans-retinoic acid protects against acute ischemic stroke by modulating neutrophil functions through STAT1 signaling. J. Neuroinflamm. 2019, 16, 175. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.J.; Airavaara, M.; Wu, K.J.; Harvey, B.K.; Liu, H.S.; Yang, Y.; Zacharek, A.; Chen, J.; Wang, Y. 9-cis retinoic acid induces neurorepair in stroke brain. Sci. Rep. 2017, 7, 4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Parker, L.E.; Yu, J.; Jones, J.W.; Liu, T.; Papanicolaou, K.N.; Talbot, C.C., Jr.; Margulies, K.B.; O’Rourke, B.; Kane, M.A.; et al. Cardiac retinoic acid levels decline in heart failure. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Xu, M.; Xu, L.; Du, H.; Shan, W.; Feng, J.; Zhai, G.; Yang, X. Decreased Serum Retinoic Acid May Predict Poor Outcome in Ischemic Stroke Patients. Neuropsychiatr. Dis. Treat. 2020, 16, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.K.; Park, T.Y.; Lee, J.A.; Moon, T.W.; Ko, M.M.; Choi, J.; Lee, M.S. Reliability and validity of the Korean Standard Pattern Identification for Stroke (K-SPI-Stroke) questionnaire. BMC Complement. Altern. Med. 2012, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Lei, R.; Ding, S.W.; Zhu, S. Skewer: A fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Krupinski, J.; Carrera, C.; Muiño, E.; Torres, N.; Al-Baradie, R.; Cullell, N.; Fernandez-Cadenas, I. DNA Methylation in Stroke. Update of Latest Advances. Comput. Struct. Biotechnol. J. 2018, 16, 1–5. [Google Scholar] [CrossRef]
- Wei, Y.; Sun, Z.; Wang, Y.; Xie, Z.; Xu, S.; Xu, Y.; Zhou, X.; Bi, J.; Zhu, Z. Methylation in the TP53 promoter is associated with ischemic stroke. Mol. Med. Rep. 2019, 20, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.; Barysheva, E.; Markov, A.; Belykh, A.; Koroleva, I.; Churkin, E.; Polonikov, A.; Ivanov, V.; Nazarenko, M. DNA Hypomethylation of the MPO Gene in Peripheral Blood Leukocytes Is Associated with Cerebral Stroke in the Acute Phase. J. Mol. Neurosci. 2021, 71, 1914–1932. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.; Xu, S.; Chen, H.; Dai, S.; Peng, X.; Wang, L.; Liang, Y.; Li, C.; Tang, B.; et al. A comprehensive association analysis between homocysteine metabolic pathway gene methylation and ischemic stroke in a Chinese hypertensive population. J. Clin. Lab. Anal. 2021, 35, e23689. [Google Scholar] [CrossRef] [PubMed]
- Zakhari, S. Alcohol metabolism and epigenetics changes. Alcohol Res. 2013, 35, 6–16. [Google Scholar]
- Zhang, T.; Jiang, Y.; Zhang, S.; Tie, T.; Cheng, Y.; Su, X.; Man, Z.; Hou, J.; Sun, L.; Tian, M.; et al. The association between homocysteine and ischemic stroke subtypes in Chinese: A meta-analysis. Medicine 2020, 99, e19467. [Google Scholar] [CrossRef]
- Nebert, D.W. Proposed role of drug-metabolizing enzymes: Regulation of steady state levels of the ligands that effect growth, homeostasis, differentiation, and neuroendocrine functions. Mol. Endocrinol. 1991, 5, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Nebert, D.W.; Wikvall, K.; Miller, W.L. Human cytochromes P450 in health and disease. Philos. Trans. R. Soc. Lond B Biol. Sci. 2013, 368, 20120431. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Song, Y.; Lin, T.; Zhou, Z.; Guo, H.; Liu, L.; Wang, B.; Liu, C.; Li, J.; et al. Plasma retinol and the risk of first stroke in hypertensive adults: A nested case-control study. Am. J. Clin. Nutr. 2019, 109, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Li, Z.; Zhou, Y.; Yu, S.; Yang, H.; Zheng, L.; Liu, Y.; Sun, Y. Metabolic Profile for Prediction of Ischemic Stroke in Chinese Hypertensive Population. J. Stroke Cerebrovasc. Dis. 2019, 28, 1062–1069. [Google Scholar] [CrossRef]
- Rowling, M.J.; Schalinske, K.L. Retinoic acid and glucocorticoid treatment induce hepatic glycine N-methyltransferase and lower plasma homocysteine concentrations in rats and rat hepatoma cells. J. Nutr. 2003, 133, 3392–3398. [Google Scholar] [CrossRef] [PubMed]
- Héberlé, É.; Bardet, A.F. Sensitivity of transcription factors to DNA methylation. Essays Biochem. 2019, 63, 727–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kribelbauer, J.F.; Lu, X.J.; Rohs, R.; Mann, R.S.; Bussemaker, H.J. Toward a Mechanistic Understanding of DNA Methylation Readout by Transcription Factors. J. Mol. Biol. 2019, 432, 1801–1815. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Ruuska, S.E.; Levinthal, D.J.; Noy, N. Distinct roles for cellular retinoic acid-binding proteins I and II in regulating signaling by retinoic acid. J. Biol. Chem. 1999, 274, 23695–23698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balmer, J.E.; Blomhoff, R. Gene expression regulation by retinoic acid. J. Lipid Res. 2002, 43, 1773–1808. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Lahiri, D.K. Cdk5 activity in the brain—Multiple paths of regulation. J. Cell Sci. 2014, 127, 2391–2400. [Google Scholar] [CrossRef] [Green Version]
- Zolfaghari, R.; Mattie, F.J.; Wei, C.H.; Chisholm, D.R.; Whiting, A.; Ross, A.C. CYP26A1 gene promoter is a useful tool for reporting RAR-mediated retinoid activity. Anal. Biochem. 2019, 577, 98–109. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Normal | SVO | p-Value |
|---|---|---|---|
| n | 115 | 56 | |
| Anthropometric characteristics | |||
| Sex (M/F) | 59/56 a | 37/19 | 0.048 * |
| Age (years) | 60.41 ± 11.06 b | 61.74 ± 12.99 | 0.489 # |
| Smoking (none/stop/active) | 68/29/18 | 23/7/26 | <0.001 * |
| Drinking (none/stop/active) | 59/9/47 | 24/5/26 | 0.645 * |
| Weight (kg) | 58.94 ± 7.64 | 58.81 ± 8.15 | 0.917 # |
| BMI (kg/m2) | 22.59 ± 1.39 | 22.31 ± 1.38 | 0.225 # |
| Waist circumference (cm) | 80.92 ± 6.89 | 80.18 ± 5.29 | 0.526 # |
| WHR | 0.872 ± 0.055 | 0.914 ± 0.061 | <0.001 # |
| Medical history | |||
| Depression (yes, %) | 1 (0.9) a | 1 (1.8) | 0.549 * |
| Migraine (yes, %) | 12 (10.4) | 7 (12.5) | 0.434 * |
| Blood parameters | |||
| WBC (×103) | 5.7 ± 1.46 | 6.85 ± 2.21 | 0.003 & |
| RBC (×106) | 4.47 ± 0.43 | 4.36 ± 0.52 | 0.021 & |
| Hg (g/dL) | 13.79 ± 1.33 | 13.37 ± 1.62 | 0.001 & |
| Hct (%) | 41.17 ± 3.80 | 39.67 ± 4.58 | 0.001 & |
| Platelet (×103/μL) | 196.92 ± 68.527 | 230.05 ± 61.21 | 0.002 & |
| GOT (U/dL) | 23.03 ± 6.88 | 25.68 ± 12.58 | 0.053 & |
| GPT (U/dL) | 19.26 ± 8.26 | 23.40 ± 19.06 | 0.008 & |
| Total cholesterol (mg/dL) | 200.30 ± 38.60 | 188.64 ± 43.34 | 0.147 & |
| Triglyceride (mg/dL) | 127.18 ± 64.56 | 132.81 ± 55.44 | 0.896 & |
| HDL cholesterol (mg/dL) | 56.97 ± 13.15 | 48.50 ± 14.41 | 0.005 & |
| LDL cholesterol (mg/dL) | 119.09 ± 32.98 | 115.88 ± 38.51 | 0.0.869 & |
| FBS (mg/dL) | 96.89 ± 9.02 | 113.707 ± 33.14 | <0.001 & |
| LDH (U/L) | 255.68 ± 111.30 | 246.94 ± 95.71 | 0.230 & |
| Na+ (mEq/L) | 143.41 ± 3.26 | 140.23 ± 3.40 | <0.001 & |
| K+ (mEq/L) | 4.39 ± 0.37 | 4.16 ± 0.63 | 0.004 & |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.-J.; Kim, M.-S.; Yim, N.-H.; Cha, M.H. Association of CYP26C1 Promoter Hypomethylation with Small Vessel Occlusion in Korean Subjects. Genes 2021, 12, 1622. https://doi.org/10.3390/genes12101622
Lee E-J, Kim M-S, Yim N-H, Cha MH. Association of CYP26C1 Promoter Hypomethylation with Small Vessel Occlusion in Korean Subjects. Genes. 2021; 12(10):1622. https://doi.org/10.3390/genes12101622
Chicago/Turabian StyleLee, Eun-Ji, Myung-Sunny Kim, Nam-Hui Yim, and Min Ho Cha. 2021. "Association of CYP26C1 Promoter Hypomethylation with Small Vessel Occlusion in Korean Subjects" Genes 12, no. 10: 1622. https://doi.org/10.3390/genes12101622
APA StyleLee, E.-J., Kim, M.-S., Yim, N.-H., & Cha, M. H. (2021). Association of CYP26C1 Promoter Hypomethylation with Small Vessel Occlusion in Korean Subjects. Genes, 12(10), 1622. https://doi.org/10.3390/genes12101622