
First Report of the Plasmid-mediated fosB Gene in Enterococcus faecalis from Pigs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. Antimicrobial Susceptibility Testing
2.3. S1-PFGE and Southern Blotting
2.4. Conjugation Assay
2.5. Fitness Experiment
2.6. Genome Sequence and Analysis
2.7. Nucleotide Accession Numbers
3. Results
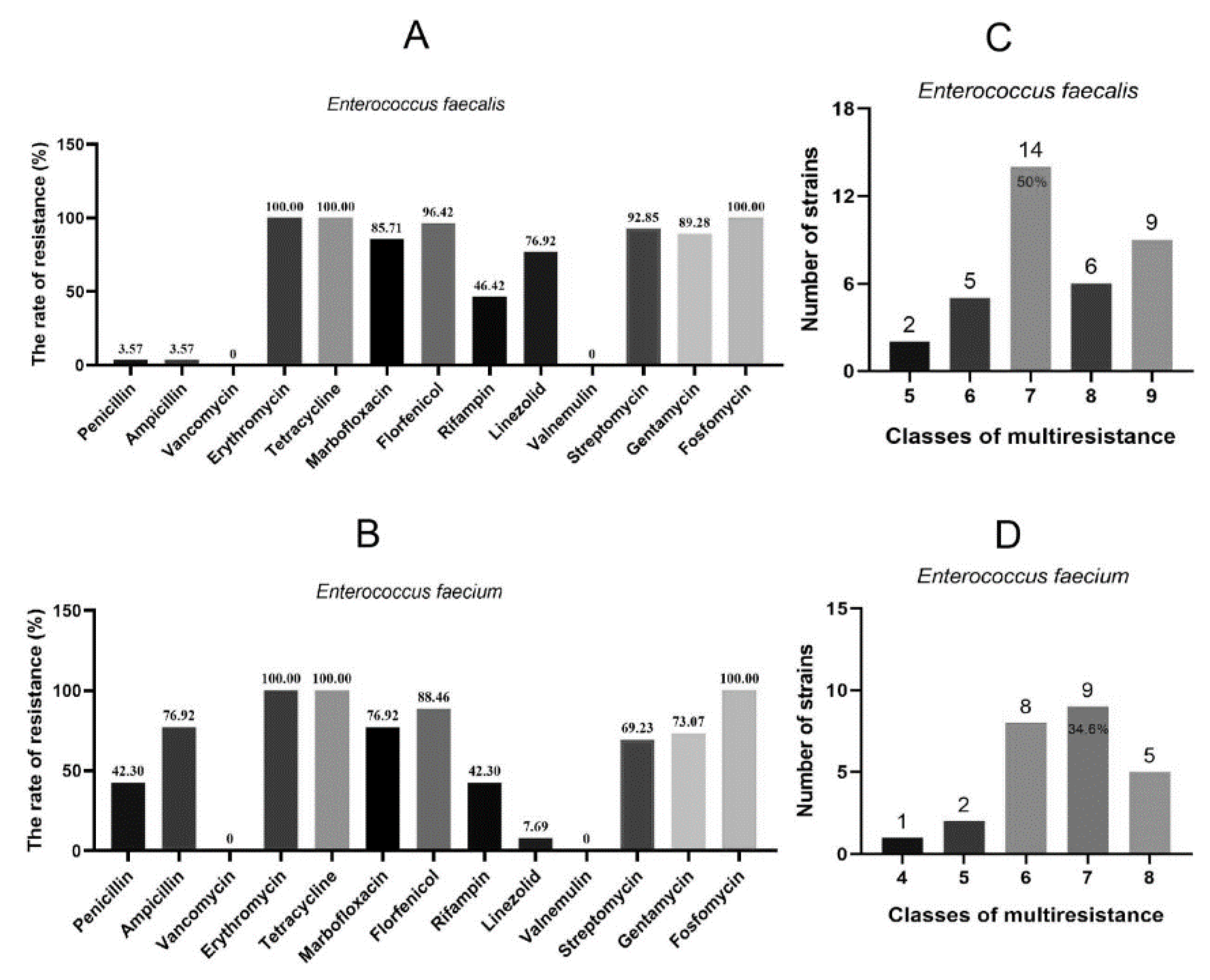
3.1. Antimicrobial Susceptibility Profiles of Fosfomycin Resistant Enterococcus spp.
3.2. Location and Transferability of fosB-Carrying Plasmid
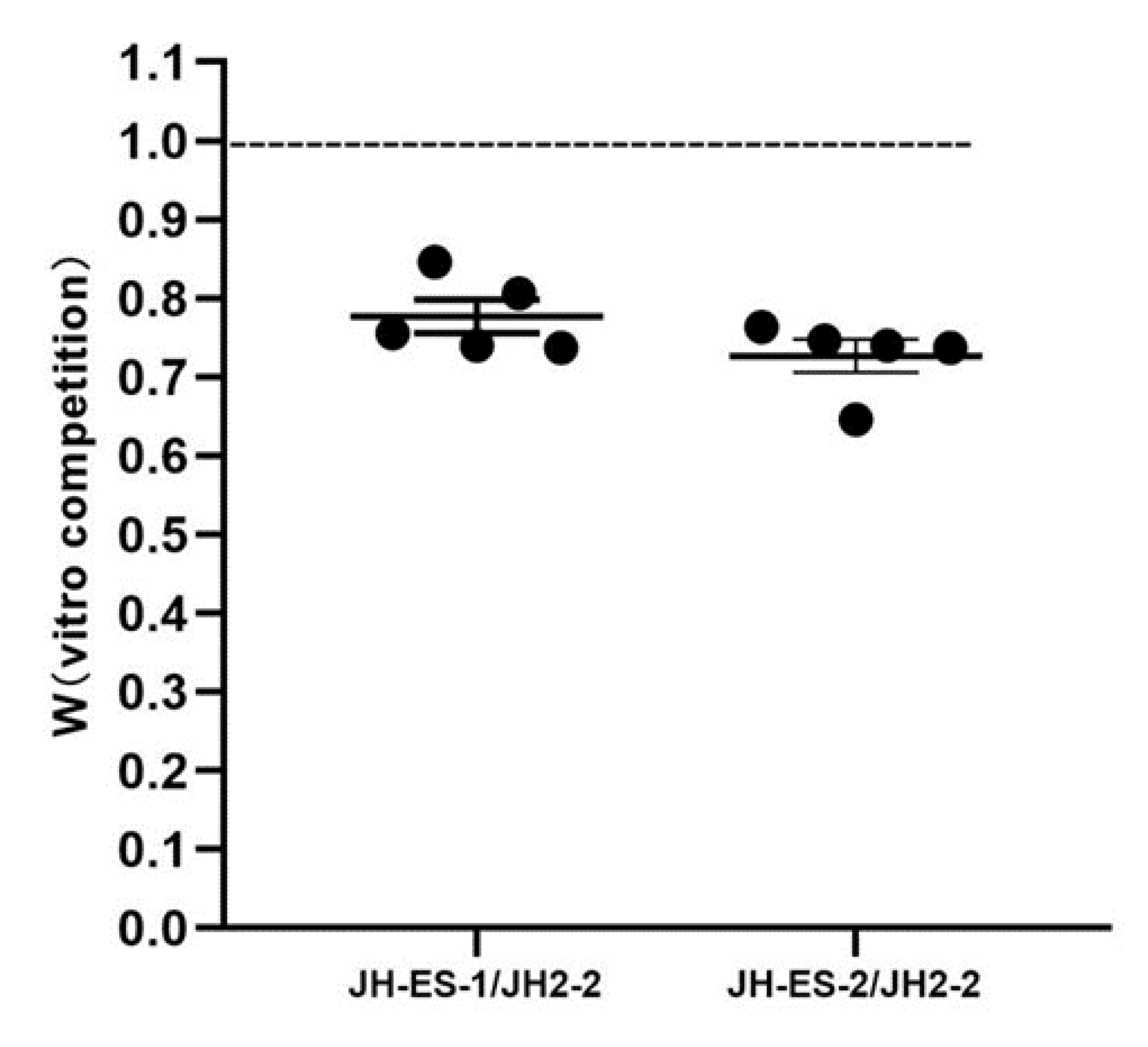
3.3. Fitness Cost of the Transconjugant
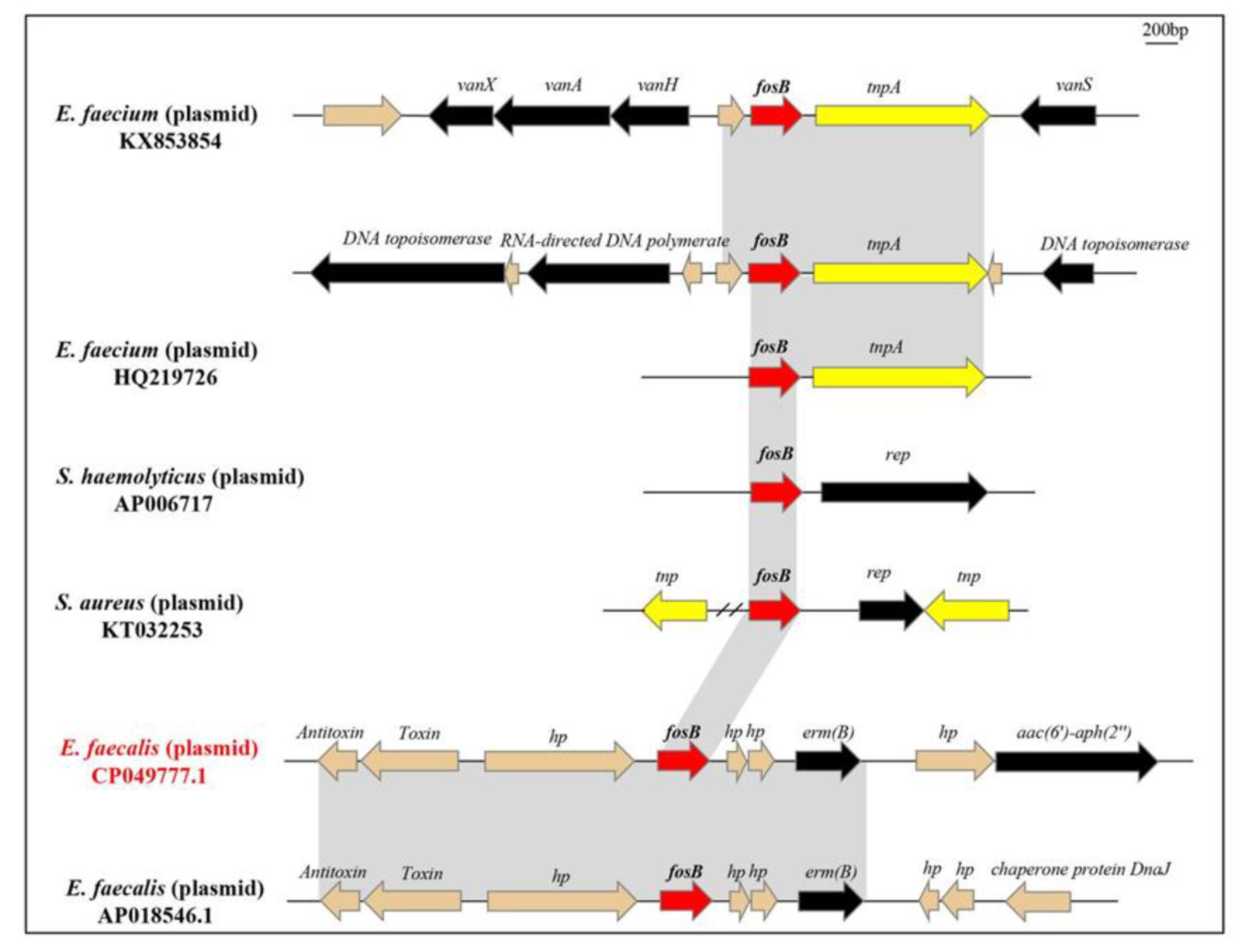
3.4. WGS Analysis of fosB-Carrying Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garcia-Solache, M.; Rice, L.B. The Enterococcus: A model of a daptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [Green Version]
- Leiro, J.; Toranzo, A.E.; Estevez, J.; Lamas, J.; Barja, J.L.; Ubeira, F.M. The humoral immune response of turbot to recently isolated pathogenic Enterococcus strains. Cross-Reactivity with other Gram-positive bacteria. Vet. Microbiol. 1996, 48, 29–39. [Google Scholar] [CrossRef]
- Snell, J.J.; Brown, D.F.; Perry, S.F.; George, R. Antimicrobial susceptibility testing of enterococci: Results of a survey conducted by the United Kingdom National External Quality Assessment Scheme for Microbiology. J. Antimicrob. Chemother. 1993, 32, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Raza, T.; Ullah, S.R.; Mehmood, K.; Andleeb, S. Vancomycin resistant Enterococci: A brief review. J. Pak. Med. Assoc. 2018, 68, 768–772. [Google Scholar]
- Reyes, K.; Bardossy, A.C.; Zervos, M. Vancomycin-Resistant Enterococci: Epidemiology, infection prevention, and control. Infect. Dis. Clin. North. Am. 2016, 30, 953–965. [Google Scholar] [CrossRef]
- Kahan, F.M.; Kahan, J.S.; Cassidy, P.J.; Kropp, H. The mechanism of action of fosfomycin (phosphonomycin). Ann. N. Y. Acad. Sci. 1974, 235, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Steinort, D.; Pillai, S.; Joukhadar, C. Fosfomycin: An old, new friend? Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 127–142. [Google Scholar] [CrossRef]
- Silver, L.L. Fosfomycin: Mechanism and resistance. Cold. Spring. Harb. Perspect. Med. 2017, 7, a025262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etienne, J.; Gerbaud, G.; Courvalin, P.; Fleurette, J. Plasmid-mediated resistance to fosfomycin in Staphylococcus epidermidis. Fems. Microbiol. Lett. 1989, 52, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bi, W.; Chen, L.; Zhang, Y.; Fang, R.; Cao, J. Molecular mechanisms and epidemiology of fosfomycin resistance in enterococci isolated from patients at a teaching hospital in China, 2013-2016. J. Glob. Antimicrob. Resist. 2020, 20, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Wu, D.; Lei, L.; Shen, Z.; Wang, Y.; Liao, K. The detection of fosfomycin resistance genes in Enterobacteriaceae from pets and their owners. Vet. Microbiol. 2016, 193, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.L.; Chan, J.; Lo, W.U.; Law, P.Y.; Chow, K.H. Plasmid-mediated fosfomycin resistance in Escherichia coli isolated from pig. Vet. Microbiol. 2013, 162, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Michalopoulos, A.S.; Livaditis, I.G.; Gougoutas, V. The revival of fosfomycin. Int. J. Infect Dis. 2011, 15, e732-9. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, C.; Lin, D.; Guo, Q.; Hu, F.; Zhu, D. The fosfomycin resistance gene fosB3 is located on a transferable, extrachromosomal circular intermediate in clinical Enterococcus faecium isolates. PLoS ONE 2013, 8, e78106. [Google Scholar] [CrossRef]
- Qu, T.T.; Shi, K.R.; Ji, J.S.; Yang, Q.; Du, X.X.; Wei, Z.Q. Fosfomycin resistance among vancomycin-resistant enterococci owing to transfer of a plasmid harbouring the fosB gene. Int. J. Antimicrob. Agents 2014, 43, 361–365. [Google Scholar] [CrossRef]
- Chen, C.; Xu, X.; Qu, T.; Yu, Y.; Ying, C.; Liu, Q. Prevalence of the fosfomycin-resistance determinant, fosB3, in Enterococcus faecium clinical isolates from China. J. Med. Microbiol. 2014, 63, 1484–1489. [Google Scholar] [CrossRef]
- Kariyama, R.; Mitsuhata, R.; Chow, J.W.; Clewell, D.B.; Kumon, H. Simple and reliable multiplex PCR assay for surveillance isolates of vancomycin-resistant enterococci. J. Clin. Microbiol. 2000, 38, 3092–3095. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, M.; Gao, Y.; Chen, L.; Wang, L. Emergence of plasmid-mediated oxazolidinone resistance gene poxtA from CC17 Enterococcus faecium of pig origin. J. Antimicrob. Chemother. 2019, 74, 2524–2530. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Huang, J.; Hu, X.; Chen, L.; Dai, X.; Sun, J. Nonconservative integration and diversity of a new family of integrative and conjugative elements associated with antibiotic resistance in zoonotic pathogen Streptococcus suis. Vet. Microbiol. 2021, 254, 109009. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents. Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, P.; Qu, T.; Chen, Y.; Hua, X.; Shi, K. Identification of novel conjugative plasmids with multiple copies of fosB that confer high-level fosfomycin resistance to vancomycin-resistant Enterococci. Front. Microbiol. 2017, 8, 1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorklund, G.; Dadar, M.; Pivina, L.; Dosa, M.D.; Semenova, Y.; Aaseth, J. The role of zinc and copper in insulin resistance and diabetes mellitus. Curr. Med. Chem. 2019, 27, 6643–6657. [Google Scholar] [CrossRef] [PubMed]
- Janka, Z. Tracing trace elements in mental functions. Ideggyogy Sz 2019, 72, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Papke, R.T.; Corral, P.; Ram-Mohan, N.; de la Haba, R.R.; Sanchez-Porro, C.; Makkay, A. Horizontal gene transfer, dispersal and haloarchaeal speciation. Life 2015, 5, 1405–1426. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents (mg/L)/Other Features | ES-1 | ES-2 | JH2-2 | JH-ES-1 | JH-ES-2 |
|---|---|---|---|---|---|
| Fosfomycin | 2048 | 2048 | 64 | 512 | 512 |
| Linezolid | 4 | 8 | 0.5 | 0.5 | 0.5 |
| Penicillin | 4 | 2 | 1 | 1 | 1 |
| Vancomycin | 2 | 4 | 2 | 2 | 2 |
| Gentamicin | HLGR | HLGR | - | HLGR | HLGR |
| Streptomycin | HLSR | HLSR | - | - | - |
| Florfenicol | 64 | 64 | 2 | 2 | 2 |
| Tetracycline | 128 | 128 | 2 | 2 | 2 |
| Erythromycin | >256 | >256 | 0.5 | 256 | 256 |
| Fusidic acid | 4 | 4 | >256 | >256 | >256 |
| Rifampicin | 2 | 2 | >256 | >256 | >256 |
| ST | 964 | 964 | |||
| Resistance genes | aph(3′)-III, dfrG, erm(A), erm(B), fexA, lsa(A), optrA, sul, aac(6′)-aph(2′′), cat, erm(B), fosB, tet(L), tet(M) | aph(3′)-III, dfrG, erm(A), erm(B), fexA, lsa(A), optrA, sul, aac(6′)-aph(2′′), cat, erm(B), fosB, tet(L), tet(M) | aac(6’)-aph(2’’), erm(B), fosB | aac(6’)-aph(2’’), erm(B), fosB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Gao, Y.; Liu, X.; Sun, N.; Huang, J.; Wang, L. First Report of the Plasmid-mediated fosB Gene in Enterococcus faecalis from Pigs. Genes 2021, 12, 1684. https://doi.org/10.3390/genes12111684
Wang X, Gao Y, Liu X, Sun N, Huang J, Wang L. First Report of the Plasmid-mediated fosB Gene in Enterococcus faecalis from Pigs. Genes. 2021; 12(11):1684. https://doi.org/10.3390/genes12111684
Chicago/Turabian StyleWang, Xiaoming, Yi Gao, Xiao Liu, Naiyan Sun, Jinhu Huang, and Liping Wang. 2021. "First Report of the Plasmid-mediated fosB Gene in Enterococcus faecalis from Pigs" Genes 12, no. 11: 1684. https://doi.org/10.3390/genes12111684
APA StyleWang, X., Gao, Y., Liu, X., Sun, N., Huang, J., & Wang, L. (2021). First Report of the Plasmid-mediated fosB Gene in Enterococcus faecalis from Pigs. Genes, 12(11), 1684. https://doi.org/10.3390/genes12111684