Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Semen Collection
2.2. Semen Assessment
2.2.1. Visual Assessment Immediately Following Collection
2.2.2. Computer Assisted Semen Assessment
2.3. RNA Isolation
2.4. Library Preparation and Sequencing
2.5. Bioinformatics Analysis
2.6. Differential Expression Analysis
3. Results
3.1. Quality of Ejaculate
3.2. Gene Expression
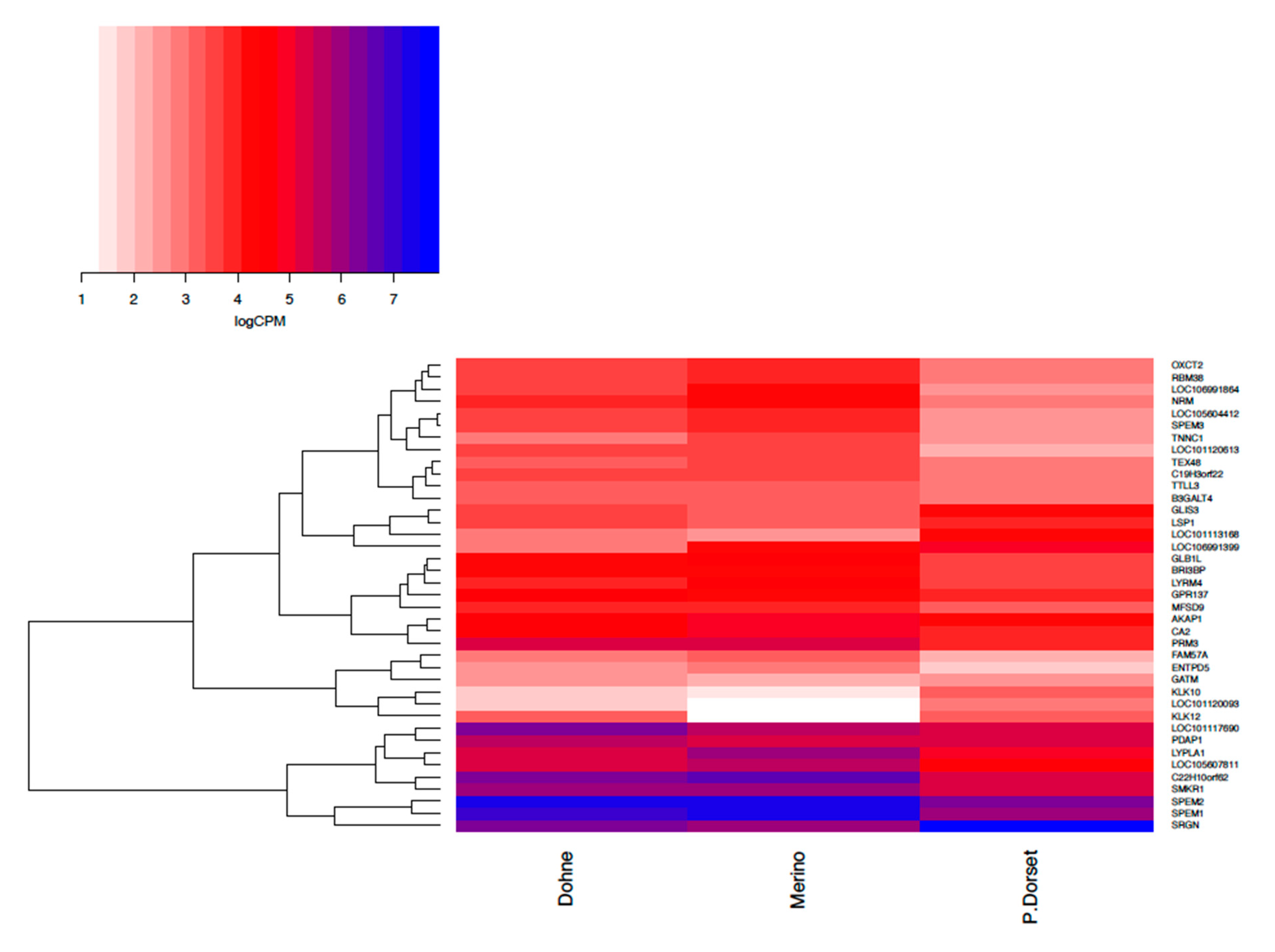
3.3. Differentially Expressed Genes (DEG) Analysis and Functional Analysis
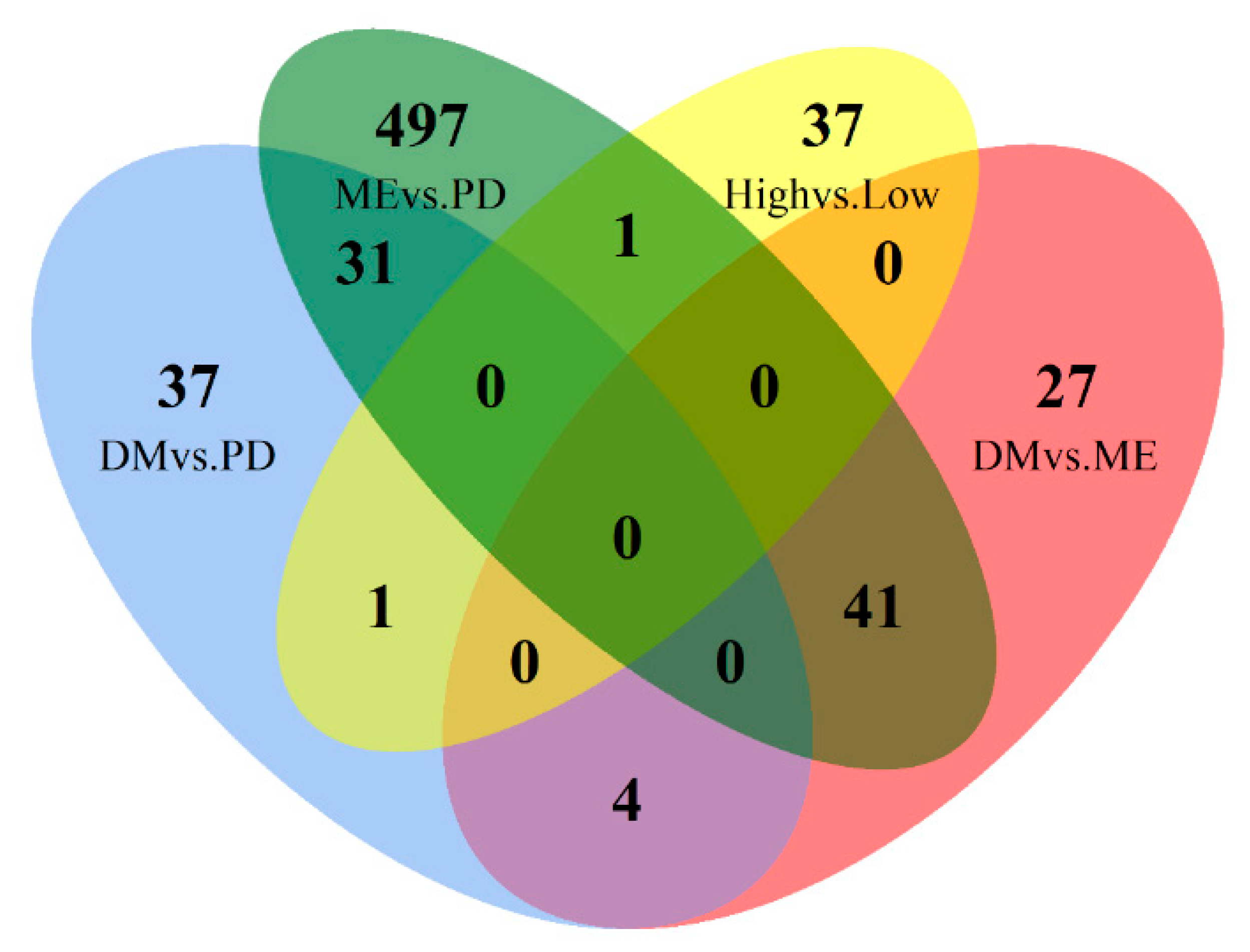
3.4. Common Differentially Expressed Genes between Breed Contrasts
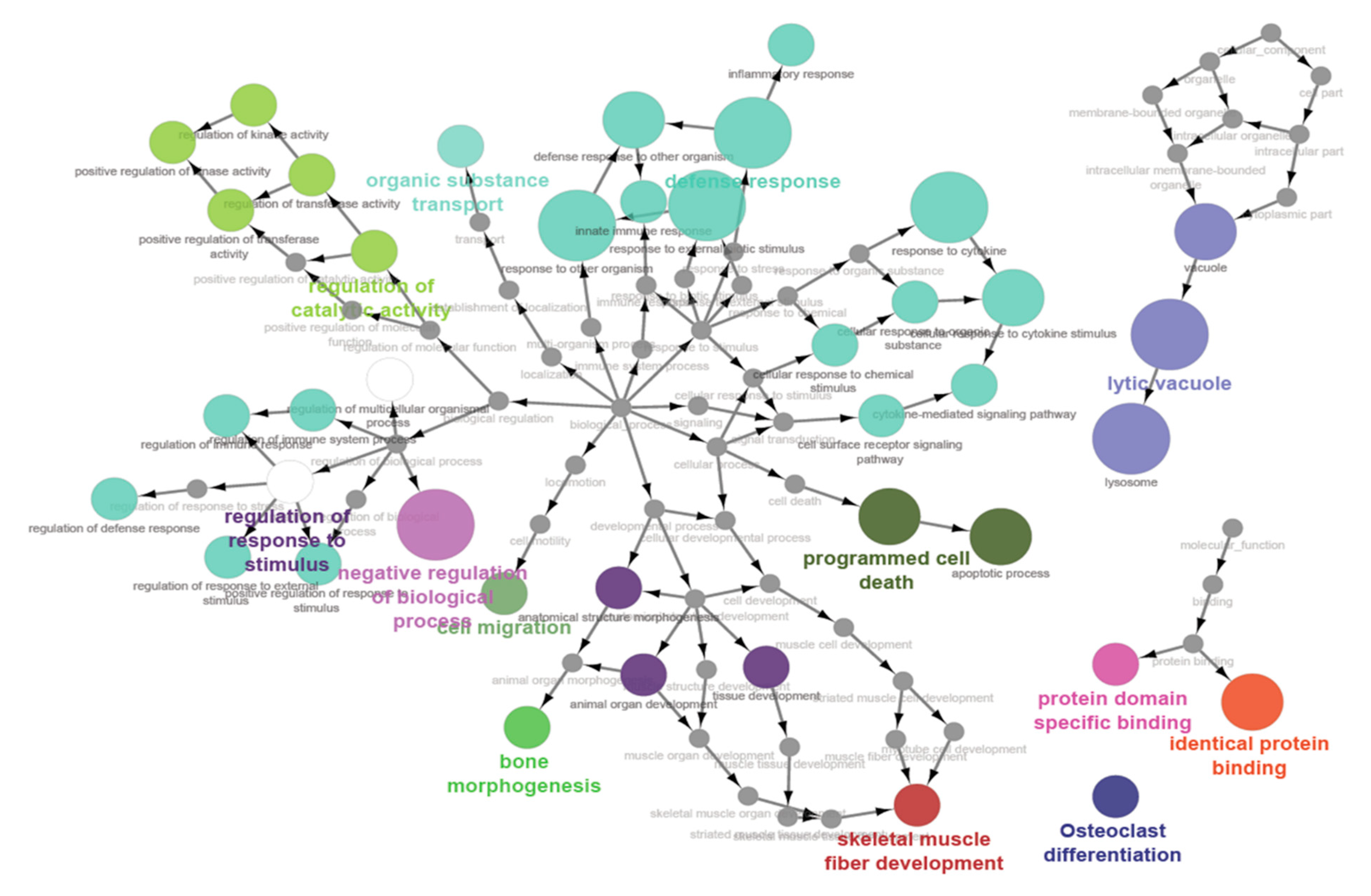
3.5. Pathway Enrichment and Gene Ontology Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbott, K. The Practice of Sheep Veterinary Medicine; University of Adelaide Press: Adelaide, Australia, 2019. [Google Scholar]
- Saacke, R.G. Sperm morphology: Its relevance to compensable and uncompensable traits in semen. Theriogenology 2008, 70, 473–478. [Google Scholar] [CrossRef]
- Reproduction in Farm Animals, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000.
- Yaniz, J.L.; Soler, C.; Santolaria, P. Computer assisted sperm morphometry in mammals: A review. Anim. Reprod. Sci. 2015, 156, 1–12. [Google Scholar] [CrossRef]
- Vijayalakshmy, K.; Kumar, D.; Virmani, M.; Jacob, N.; Kumar, P. Sperm Transcriptomics: An Emerging Technique to Assess Male Fertility. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1188–1200. [Google Scholar] [CrossRef]
- Boerke, A.; Dieleman, S.J.; Gadella, B.M. A possible role for sperm RNA in early embryo development. Theriogenology 2007, 68 (Suppl. 1), S147–S155. [Google Scholar] [CrossRef] [PubMed]
- Rando, O.J. Daddy issues: Paternal effects on phenotype. Cell 2012, 151, 702–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellappan, S.; Sivashanmugam, P.; Lakshminarayana, S.; Atul, P.K.; Binsila, B.K.; Arunachalam, A.; Janivara Parameshwaraiah, R. Occurrence and functional significance of the transcriptome in bovine (Bos taurus) spermatozoa. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Kasimanickam, V.; Kastelic, J. MicroRNA in sperm from Duroc, Landrace and Yorkshire boars. Sci Rep 2016, 6, 32954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suliman, Y.; Becker, F.; Wimmers, K. Implication of transcriptome profiling of spermatozoa for stallion fertility. Reprod. Fertil. Dev. 2018, 30, 1087. [Google Scholar] [CrossRef] [PubMed]
- Sendler, E.; Johnson, G.D.; Mao, S.; Goodrich, R.J.; Diamond, M.P.; Hauser, R.; Krawetz, S.A. Stability, delivery and functions of human sperm RNAs at fertilization. Nucleic Acids Res. 2013, 41, 4104–4117. [Google Scholar] [CrossRef] [PubMed]
- Kadivar, A.; Heidari Khoei, H.; Hassanpour, H.; Golestanfar, A.; Ghanaei, H. Correlation of Adiponectin mRNA Abundance and Its Receptors with Quantitative Parameters of Sperm Motility in Rams. Int. J. Fertil. Steril. 2016, 10, 127–135. [Google Scholar]
- Luna, C.; Serrano, E.; Domingo, J.; Casao, A.; Pérez-Pé, R.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Expression, cellular localization, and involvement of the pentose phosphate pathway enzymes in the regulation of ram sperm capacitation. Theriogenology 2016, 86, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Ureña, I.; González, C.; Ramón, M.; Godia, M.; Clop, A.; Calvo, J.H.; Carabaño, M.J.; Serrano, M. Deciphering climatic conditions effect on the ovine sperm transcription by RNA-seq. In Proceedings of the 37th International Society for Animal Genetics Conference (ISAG 2019), Lleida, Spain, 7–12 July 2019. [Google Scholar]
- Abril-Sánchez, S.; Freitas-de-Melo, A.; Damián, J.P.; Giriboni, J.; Villagrá-García, A.; Ungerfeld, R. Ejaculation does not contribute to the stress response to electroejaculation in sheep. Reprod. Domest. Anim. 2017, 52, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Damián, J.P.; Ungerfeld, R. The Stress Response of Frequently Electroejaculated Rams to Electroejaculation: Hormonal, Physiological, Biochemical, Haematological and Behavioural Parameters: Stress Response to Electroejaculation in Rams. Reprod. Domest. Anim. 2011, 46, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Evans, G. Salamon’s Artificial Insemination of Sheep and Goats; Butterworths: Sydney, Australia, 1987. [Google Scholar]
- Saacke, R.G. Components of Semen Quality. J. Anim. Sci. 1982, 55, 1–13. [Google Scholar] [PubMed]
- Gillan, L.; Kroetsch, T.; Chis Maxwell, W.M.; Evans, G. Assessment of in vitro sperm characteristics in relation to fertility in dairy bulls. Anim. Reprod. Sci. 2008, 103, 201–214. [Google Scholar] [CrossRef]
- Kasimanickam, V.; Kasimanickam, R. An Efficient Approach for RNA Extraction from Boar Sperm and Seminal Plasma. BioProtoc 2019, 9. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinform. Oxf. Engl. 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barter, R.L.; Yu, B. Superheat: An R Package for Creating Beautiful and Extendable Heatmaps for Visualizing Complex Data. J. Comput. Graph. Stat. 2018, 27, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerri, R.L.A.; Thompson, I.M.; Kim, I.H.; Ealy, A.D.; Hansen, P.J.; Staples, C.R.; Li, J.L.; Santos, J.E.P.; Thatcher, W.W. Effects of lactation and pregnancy on gene expression of endometrium of Holstein cows at day 17 of the estrous cycle or pregnancy. J. Dairy Sci. 2012, 95, 5657–5675. [Google Scholar] [CrossRef]
- Kiewisz, J.; Krawczynski, K.; Lisowski, P.; Blitek, A.; Zwierzchowski, L.; Ziecik, A.J.; Kaczmarek, M.M. Global gene expression profiling of porcine endometria on Days 12 and 16 of the estrous cycle and pregnancy. Theriogenology 2014, 82, 897–909. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, Z.; Khan, M.J.; Gao, J.; Loor, J.J. Clock circadian regulator (CLOCK) gene network expression patterns in bovine adipose, liver, and mammary gland at 3 time points during the transition from pregnancy into lactation. J. Dairy Sci. 2015, 98, 4601–4612. [Google Scholar] [CrossRef]
- Lienau, J.; Schell, H.; Epari, D.R.; Schütze, N.; Jakob, F.; Duda, G.N.; Bail, H.J. CYR61 (CCN1) Protein Expression during Fracture Healing in an Ovine Tibial Model and Its Relation to the Mechanical Fixation Stability. J. Orthop. Res. 2006, 24, 254–262. [Google Scholar] [CrossRef]
- Oien, D.B.; Osterhaus, G.L.; Latif, S.A.; Pinkston, J.W.; Fulks, J.; Johnson, M.; Fowler, S.C.; Moskovitz, J. MsrA knockout mouse exhibits abnormal behavior and brain dopamine levels. Free Radic. Biol. Med. 2008, 45, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Ren, M.X.; Chen, G.p.; Jin, Z.m.; Wang, G. Rrp15 affects cell cycle, proliferation, and apoptosis in NIH3T3 cells. Febs Open Bio. 2016, 6, 1085–1092. [Google Scholar] [CrossRef]
- Crookenden, M.A.; Heiser, A.; Murray, A.; Dukkipati, V.S.R.; Kay, J.K.; Loor, J.J.; Meier, S.; Mitchell, M.D.; Moyes, K.M.; Walker, C.G.; et al. Corrigendum to “Parturition in dairy cows temporarily alters the expression of genes in circulating neutrophils” (J. Dairy Sci. 99:6470–6483). J. Dairy Sci. 2018, 101, 3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicik, Z.; Gajewska, M.; Majewska, A.; Walkiewicz, D.; Osińska, E.; Motyl, T. Characterization of microRNA profile in mammary tissue of dairy and beef breed heifers. J. Anim. Breed. Genet. (1986) 2016, 133, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Liang, Y.; Kim, T.; Cooper, J.A. Physiological role of the interaction between CARMIL1 and capping protein. Mol. Biol. Cell 2013, 24, 3047–3055. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Waurich, B.; Wensch-Dorendorf, M.; Bickhart, D.M.; Swalve, H.H. A genome-wide association study of calf birth weight in Holstein cattle using single nucleotide polymorphisms and phenotypes predicted from auxiliary traits. J. Dairy Sci. 2014, 97, 3156–3172. [Google Scholar] [CrossRef] [Green Version]
- Katarzyna, P.; Paul, A.M.; Colm, J.R.; John, A.B.; Jane, A.I.; Tharmala, T.; Anthony, C.; Alan, B.; Brendan, J.L.; Alexander, C.O.E.; et al. Molecular aspects of mucin biosynthesis and mucus formation in the bovine cervix during the periestrous period. Physiol. Genom. 2012, 44, 1165–1178. [Google Scholar] [CrossRef]
- Parker Gaddis, K.L.; Null, D.J.; Cole, J.B. Explorations in genome-wide association studies and network analyses with dairy cattle fertility traits. J. Dairy Sci. 2016, 99, 6420–6435. [Google Scholar] [CrossRef]
- Grigoletto, L.; Santana, M.H.A.; Bressan, F.F.; Eler, J.P.; Nogueira, M.F.G.; Kadarmideen, H.N.; Baruselli, P.S.; Ferraz, J.B.S.; Brito, L.F. Genetic Parameters and Genome-Wide Association Studies for Anti-Müllerian Hormone Levels and Antral Follicle Populations Measured After Estrus Synchronization in Nellore Cattle. Animal 2020, 10, 1185. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Shen, M.; Xie, X.-L.; Liu, G.-J.; Xu, Y.-X.; Lv, F.-H.; Yang, H.; Yang, Y.-L.; Liu, C.-B.; et al. Whole-genome resequencing of wild and domestic sheep identifies genes associated with morphological and agronomic traits. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Wang, W.; Li, C.; Li, F.; Wang, X.; Zhang, X.; Liu, T.; Nian, F.; Yue, X.; Li, F.; Pan, X.; et al. Effects of early feeding on the host rumen transcriptome and bacterial diversity in lambs. Sci. Rep. 2016, 6, 32479. [Google Scholar] [CrossRef]
- González-Prendes, R.; Quintanilla, R.; Cánovas, A.; Manunza, A.; Figueiredo Cardoso, T.; Jordana, J.; Noguera, J.L.; Pena, R.N.; Amills, M. Joint QTL mapping and gene expression analysis identify positional candidate genes influencing pork quality traits. Sci. Rep. 2017, 7, 39830. [Google Scholar] [CrossRef] [Green Version]
- Cockrum, R.R.; Austin, K.J.; Kim, J.W.; Garbe, J.R.; Fahrenkrug, S.C.; Taylor, J.F.; Cammack, K.M. Differential gene expression of ewes varying in tolerance to dietary nitrate. J. Anim. Sci. 2010, 88, 3187. [Google Scholar] [CrossRef] [PubMed]
- Rengaraj, D.; Gao, F.; Liang, X.-H.; Yang, Z.-M. Expression and regulation of type II integral membrane protein family members in mouse male reproductive tissues. Endocrine 2007, 31, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Chen, D.; Zhen, Z.; Ao, J.; Yuan, X.; Gao, X. Annexin A2 positively regulates milk synthesis and proliferation of bovine mammary epithelial cells through the mTOR signaling pathway. J. Cell. Physiol. 2018, 233, 2464–2475. [Google Scholar] [CrossRef] [PubMed]
- Kuzmany, A.; Havlicek, V.; Wrenzycki, C.; Wilkening, S.; Brem, G.; Besenfelder, U. Expression of mRNA, before and after freezing, in bovine blastocysts cultured under different conditions. Theriogenology 2011, 75, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, W.; Zheng, X.; Liu, Z.; Yang, G.; Hu, X.; Mou, C. Comparative investigation of coarse and fine wool sheep skin indicates the early regulators for skin and wool diversity. Gene 2020, 758, 144968. [Google Scholar] [CrossRef] [PubMed]
- Killeen, A.P.; Morris, D.G.; Kenny, D.A.; Mullen, M.P.; Diskin, M.G.; Waters, S.M. Global gene expression in endometrium of high and low fertility heifers during the mid-luteal phase of the estrous cycle. Bmc Genom. 2014, 15, 234. [Google Scholar] [CrossRef] [Green Version]
- Gorni, C.; Garino, C.; Iacuaniello, S.; Castiglioni, B.; Stella, A.; Restelli, G.L.; Pagnacco, G.; Mariani, P. Transcriptome analysis to identify differential gene expression affecting meat quality in heavy Italian pigs: Transcriptome analysis to identify differential gene expression. Anim. Genet. 2011, 42, 161–171. [Google Scholar] [CrossRef]
- Knight, J.S.; Baird, D.B.; Hein, W.R.; Pernthaner, A. The gastrointestinal nematode Trichostrongylus colubriformis down-regulates immune gene expression in migratory cells in afferent lymph. BMC Immunol. 2010, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Elmes, M.; Szyszka, A.; Pauliat, C.; Clifford, B.; Daniel, Z.; Cheng, Z.; Wathes, C.; McMullen, S. Maternal age effects on myometrial expression of contractile proteins, uterine gene expression, and contractile activity during labor in the rat. Physiol. Rep. 2015, 3, e12305-n/a. [Google Scholar] [CrossRef] [Green Version]
- Waters, S.M.; Coyne, G.S.; Kenny, D.A.; Morris, D.G. Effect of dietary n-3 polyunsaturated fatty acids on transcription factor regulation in the bovine endometrium. Mol. Biol. Rep. 2014, 41, 2745–2755. [Google Scholar] [CrossRef]
- Ogi, T.; Mimura, J.; Hikida, M.; Fujimoto, H.; Fujii-Kuriyama, Y.; Ohmori, H. Expression of human and mouse genes encoding polκ: Testis-specific developmental regulation and AhR-dependent inducible transcription. Genes Cells Devoted Mol. Cell. Mech. 2001, 6, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Tizioto, P.C.; Coutinho, L.L.; Decker, J.E.; Schnabel, R.D.; Rosa, K.O.; Oliveira, P.S.N.; Souza, M.M.; Mourão, G.B.; Tullio, R.R.; Chaves, A.S.; et al. Global liver gene expression differences in Nelore steers with divergent residual feed intake phenotypes. BMC Genom. 2015, 16, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholizadeh, M.; Rahimi-Mianji, G.; Nejati-Javaremi, A. Genomewide association study of body weight traits in Baluchi sheep. J. Genet. 2015, 94, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Puttabyatappa, M.; Guo, X.; Dou, J.; Dumesic, D.; Bakulski, K.M.; Padmanabhan, V. Developmental Programming: Sheep Granulosa and Theca Cell–Specific Transcriptional Regulation by Prenatal Testosterone. Endocrinology (Philadelphia) 2020, 161. [Google Scholar] [CrossRef]
- Kern, R.J.; Lindholm-Perry, A.K.; Freetly, H.C.; Kuehn, L.A.; Rule, D.C.; Ludden, P.A. Rumen papillae morphology of beef steers relative to gain and feed intake and the association of volatile fatty acids with kallikrein gene expression. Livest. Sci. 2016, 187, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Stafuzza, N.B.; Silva, R.M.d.O.; Fragomeni, B.d.O.; Masuda, Y.; Huang, Y.; Gray, K.; Lourenco, D.A.L. A genome-wide single nucleotide polymorphism and copy number variation analysis for number of piglets born alive. BMC Genom. 2019, 20, 321. [Google Scholar] [CrossRef]
- Chen, L.; Liu, K.; Zhao, Z.; Blair, H.T.; Zhang, P.; Li, D.; Ma, R.Z. Identification of Sheep Ovary Genes Potentially Associated with Off-season Reproduction. J. Genet. Genom. 2012, 39, 181–190. [Google Scholar] [CrossRef]
- Bhide, A.; Vuolteenaho, O.; Haapsamo, M.; Erkinaro, T.; Rasanen, J.; Acharya, G. Effect of Hypoxemia with or without Increased Placental Vascular Resistance on Fetal Left and Right Ventricular Myocardial Performance Index in Chronically Instrumented Sheep. Ultrasound Med. Biol. 2016, 42, 2589–2598. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, F.; Mukiibi, R.; Chen, L.; Vinsky, M.; Plastow, G.; Basarab, J.; Stothard, P.; Li, C. Genetic architecture of quantitative traits in beef cattle revealed by genome wide association studies of imputed whole genome sequence variants: II: Carcass merit traits. BMC Genom. 2020, 21, 38. [Google Scholar] [CrossRef]
- Cheng, Y.; Gaughan, J.; Midic, U.; Han, Z.; Liang, C.-G.; Patel, B.G.; Latham, K.E. Systems genetics implicates cytoskeletal genes in oocyte control of cloned embryo quality. Genetics (Austin) 2013, 193, 877–896. [Google Scholar] [CrossRef] [Green Version]
- Georgiadis, A.P.; Kishore, A.; Zorrilla, M.; Jaffe, T.M.; Sanfilippo, J.S.; Volk, E.; Rajkovic, A.; Yatsenko, A.N. High quality RNA in semen and sperm: Isolation, analysis and potential application in clinical testing. J. Urol. 2015, 193, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Matteo, B.; Christian, M.; Justin, F.; Clint, S.; Francesco, T. Genome-wide association study for carcass quality traits and growth in purebred and crossbred pigs 1. J. Anim. Sci. 2020, 98, 1–11. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Ma, L.; Lin, X.; Zhang, D.; Li, Z.; Wu, Y.; Zheng, C.; Feng, X.; Liao, S.; et al. Retinoic Acid Is Sufficient for the In Vitro Induction of Mouse Spermatocytes. Stem Cell Rep. 2016, 7, 80–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm-Perry, A.K.; Butler, A.R.; Kern, R.J.; Hill, R.; Kuehn, L.A.; Wells, J.E.; Oliver, W.T.; Hales, K.E.; Foote, A.P.; Freetly, H.C. Differential gene expression in the duodenum, jejunum and ileum among crossbred beef steers with divergent gain and feed intake phenotypes. Anim. Genet. 2016, 47, 408–427. [Google Scholar] [CrossRef] [PubMed]
- Forde, N.; Carter, F.; Fair, T.; Crowe, M.A.; Evans, A.C.O.; Spencer, T.E.; Bazer, F.W.; McBride, R.; Boland, M.P.; Gaora, P.O.; et al. Progesterone-Regulated Changes in Endometrial Gene Expression Contribute to Advanced Conceptus Development in Cattle. Biol. Reprod. 2009, 81, 784–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.F.; Li, Y.K.; Ren, K.; Xie, Y.J.; Yin, W.D.; Mo, Z.C. Characterization of cholesterol metabolism in Sertoli cells and spermatogenesis (Review). Mol. Med. Rep. 2018, 17, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, H.; Sheng, X.; Cui, X.; Gu, M.; Liu, Y.; Qi, X.; Xing, S.; Guo, Y. Epidermal growth factor-mediated mitogen-activated protein kinase3/1 pathway is conducive to in vitro maturation of sheep oocytes. PLoS ONE 2015, 10, e0120418. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.H.; Li, D.J.; Su, H.; Hu, J.Q.; Li, N.; Li, S.J. Molecular cloning, expression, and imprinting status of maternally expressed gene 8 (Meg8) in dairy cattle. Russ. J. Genet. 2011, 47, 994–998. [Google Scholar] [CrossRef]
- Stephen, G.M.; Jennie, E.P.; Ben, J.H.; Amanda, J.C.; Kathryn, E.K.; Donagh, P.B.; Matt, M.; Paul, C.; Pat, L.; Trudee, F.; et al. Differentially Expressed Genes in Endometrium and Corpus Luteum of Holstein Cows Selected for High and Low Fertility Are Enriched for Sequence Variants Associated with Fertility1. Biol. Reprod. 2016, 94. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, J.; Campanile, G.; Plastow, G.; Zhang, C.; Wang, Z.; Cassandro, M.; Gasparrini, B.; Salzano, A.; Hua, G.; et al. Novel insights into the genetic basis of buffalo reproductive performance. BMC Genom. 2018, 19, 814. [Google Scholar] [CrossRef]
- Hatzirodos, N.; Irving-Rodgers, H.F.; Hummitzsch, K.; Harland, M.L.; Morris, S.E.; Rodgers, R.J. Transcriptome profiling of granulosa cells of bovine ovarian follicles during growth from small to large antral sizes. BMC Genom. 2014, 15, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-S.; Shin, D. Genome-Wide Association Studies Associated with Backfat Thickness in Landrace and Yorkshire Pigs. Genom. Inform. 2018, 16, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, P.; Carmelo, V.A.O.; Kadarmideen, H.N. Genome-Wide Epistatic Interaction Networks Affecting Feed Efficiency in Duroc and Landrace Pigs. Front. Genet. 2020, 11, 121. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, J.; Wang, J.; Ren, Y.; Zhu, Y.; Sun, H. Dynamic changes of miR166s at both the transcriptional and post-transcriptional levels during somatic embryogenesis in Lilium. Sci. Hortic. 2020, 261, 108928. [Google Scholar] [CrossRef]
- Liu, G.; Liu, R.; Tang, X.; Cao, J.; Zhao, S.; Yu, M. Expression profiling reveals genes involved in the regulation of wool follicle bulb regression and regeneration in sheep. Int. J. Mol. Sci. 2015, 16, 9152–9166. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Li, S.; Zheng, X.; Chen, W.; Li, X.; Liu, Z.; Hu, Y.; Qiao, H.; Qi, Q.; Pei, Q.; et al. Transcriptome Reveals Long Non-coding RNAs and mRNAs Involved in Primary Wool Follicle Induction in Carpet Sheep Fetal Skin. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Norris, B.J.; Bower, N.I.; Smith, W.J.M.; Cam, G.R.; Reverter, A. Gene expression profiling of ovine skin and wool follicle development using a combined ovine–bovine skin cDNA microarray. Anim. Prod. Sci. 2005, 45, 867–877. [Google Scholar] [CrossRef]
- Cargill, E.J.; Nissing, N.J.; Grosz, M.D. Single nucleotide polymorphisms concordant with the horned/polled trait in Holsteins. BMC Res. Notes 2008, 1, 128. [Google Scholar] [CrossRef] [Green Version]
- Tachikawa, M.; Watanabe, M.; Hori, S.; Fukaya, M.; Ohtsuki, S.; Asashima, T.; Terasaki, T. Distinct spatio-temporal expression of ABCA and ABCG transporters in the developing and adult mouse brain. J. Neurochem. 2005, 95, 294–304. [Google Scholar] [CrossRef]
- Luo, K.; Yuan, W.; Zhu, C.; Li, Y.; Wang, Y.; Zeng, W.; Jiao, W.; Liu, M.; Wu, X. Expression of a novel Krüpple-like zinc-finger gene, ZNF382, in human heart. Biochem. Biophys. Res. Commun. 2002, 299, 606–612. [Google Scholar] [CrossRef]
- Hudson, N.J.; Reverter, A.; Griffiths, W.J.; Yutuc, E.; Wang, Y.; Jeanes, A.; McWilliam, S.; Pethick, D.W.; Greenwood, P.L. Gene expression identifies metabolic and functional differences between intramuscular and subcutaneous adipocytes in cattle. BMC Genom. 2020, 21, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, C.-C.; Yang, W.-X. New insights to the ubiquitin–proteasome pathway (UPP) mechanism during spermatogenesis. Mol. Biol. Rep. 2012, 40, 3213–3230. [Google Scholar] [CrossRef] [PubMed]
- Nian, H.; Fan, C.; Liao, S.; Shi, Y.; Zhang, K.; Liu, Y.; Han, C. RNF151, a testis-specific RING finger protein, interacts with dysbindin. Arch. Biochem. Biophys. 2007, 465, 157–163. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Assessment Parameters | Merino | Dohne | Poll Dorset |
|---|---|---|---|
| Motility | 64.25 ± 6.69 | 75.99 ± 4.30 | 74.62 ± 3.86 |
| VSL (μm/s) | 110.51 ± 11.30 | 105.85 ± 8.18 | 125.85 ± 11.73 |
| VCL (μm/s) | 278.86 ± 27.16 | 238.71 ± 22.95 | 277.10 ± 23.11 |
| VAP (μm/s) | 10.17 ± 1.00 | 9.15 ± 1.04 | 10.87 ± 1.05 |
| ALH (μm/s) | 149.93 ± 12.78 | 139.18 ± 10.08 | 161.65 ± 11.58 |
| LIN | 43.50 ± 3.63 | 37.93 ± 3.89 | 51.19 ± 4.93 |
| STR (%) | 80.00 ± 2.49 | 57.69 ± 3.64 | 75.57 ± 3.44 |
| WOB | 54.55 ± 2.92 | 47.86 ± 3.42 | 57.55 ± 3.69 |
| BCF (Hz) | 36.35 ± 1.21 | 26.67 ± 2.28 | 33.34 ± 1.74 |
| Membrane Integrity (%) | 89.18 ± 2.40 | 82.44 ± 2.61 | 91.54 ± 2.80 |
| Morphologically Normal (%) | 83.51 ± 3.31 | 96.18 ± 0.43 | 89.28 ± 1.79 |
| Total Raw Reads | Total Clean Reads | Mapped Reads | Mapped Rate % | |
|---|---|---|---|---|
| Merino | 564,403,692 | 533,122,904 | 693,063,473 | 85.8% |
| Dohne | 548,882,390 | 517,471,028 | 222,273,782 | 89.3% |
| Poll Dorset | 683,941,522 | 643,781,418 | 271,998,185 | 89.0% |
| Total | 1,797,227,604 | 1,694,375,350 | 1,187,335,440 | 88.0% |
| Differentially Expressed Genes | Upregulated | Downregulated | |
|---|---|---|---|
| Dohne vs. Merino | 72 | 25 | 47 |
| Dohne vs. Poll Dorset | 73 | 41 | 31 |
| Merino vs. Poll Dorset | 570 | 234 | 336 |
| Ejaculate Quality Contrasts | 39 | 6 | 33 |
| Gene | logFC | Reference |
|---|---|---|
| Dohne vs. Merino | ||
| Mevalonate Kinase (MVK) A | 2.16 | [29] |
| Tensin 3 (TNS3) A | −1.82 | [30] |
| 5′-Aminolevulinate Synthase 1 (ALAS1) A | 1.31 | [31] |
| Cellular Communication Network Factor 1 (CCN1) A | −2.17 | [32] |
| Methionine Sulfoxide Reductase A (MSRA) A | 2.33 | [33] |
| Ribosomal RNA Processing 15 Homolog (RRP15) A | −2.57 | [34] |
| Peroxisome Proliferator Activated Delta (PPARD) B | 1.47 | [35] |
| Mediator Complex Subunit 6 (MED6) A | −1.35 | [36] |
| Capping Protein Regulator And Myosin 1 Linked (CARMIL1) A | 1.26 | [37] |
| Fibronectin Leucine Rich Transmembrane Protein 2 (FLRT2) A | −2.08 | [38] |
| Merino vs. Poll Dorset | ||
| ATPase Sarcoplasmic/Endoplasmic Reticulum Ca2+ Transporting 3 (ATP2A3) A | −2.59 | [39] |
| Solute Carrier Family 35 Member A5 (SLC35A5) A | 1.12 | [40] |
| BLOC-1 Related Complex Subunit 5 (BORCS5) A | 2.08 | [41] |
| Methionine Sulfoxide Reductase A (MSRA) B | −2.64 | [42] |
| Keratin 4 (KRT4) B | −1.78 | [43] |
| Family With Sequence Similarity 210 Member B (FAM210B) B | −1.47 | [44] |
| NTPase KAP Family P-Loop Domain Containing 1 (NKPD1) B | 2.89 | [45] |
| Integral Membrane Protein 2C (ITM2C) A | −3.05 | [46] |
| Annexin A2 (ANAX2) A | −1.46 | [47] |
| Solute Carrier Family 2 Member 3 (SLC2A3) A | −2.98 | [48] |
| Dohne vs. Poll Dorset | ||
| Peptidase Domain Containing Associated With Muscle Regeneration 1 (PAMR1) B | −2.56 | [49] |
| Lymphocyte Antigen 6 Family Member E (LY6E) A | −1.33 | [50] |
| Arginine And Glutamate Rich 1 (ARGLU1) B | 1.64 | [51] |
| Cysteine Dioxygenase Type 1 (CDO1) B | 2.10 | [52] |
| Valyl-TRNA Synthetase 2, Mitochondrial (VARS2) A | −2.13 | [53] |
| Tumor Protein P53 Inducible Protein 11 (TP53I11) A | 2.15 | [54] |
| DNA Polymerase Kappa (POLK) A | 1.65 | [55] |
| F-Box And Leucine Rich Repeat Protein 14 (FBXL14) B | −1.25 | [56] |
| Mannosidase Alpha Class 1A Member 1 (MAN1A1) A | −2.26 | [57] |
| Chintase 3 Like 1 (CHI3L1) A | −2.29 | [58] |
| Ejaculate Quality Contrast | ||
| Kallikrein Related Peptidase 12 (KLK12) B | 2.65 | [59] |
| SPEM Family Member 2 (SPEM2) A | −2.46 | [60] |
| Lysophospholipase 1 (LYPLA1) A | −2.31 | [61] |
| Troponin C1, Slow Skeletal And Cardiac Type (TNNC1) B | −2.23 | [62] |
| LYR Motif Containing 4 (LYRM4) B | −1.97 | [63] |
| Major Facilitator Superfamily Domain Containing 9 (MFSD9) A | −1.63 | [64] |
| 3-Oxoacid CoA-Transferase 2 (OXCT2) A | −1.88 | [65] |
| Small Lysine Rich Protein 1 (SMKR1) B | −1.96 | [66] |
| Family With Sequence Similarity 57 Member A (FAM57A) A | −1.72 | [67] |
| Serglycin (SRGN) A | 2.38 | [68] |
| Comparison Groups | Number of Common Differentially Expressed Genes | Common Differentially Expressed Genes |
|---|---|---|
| Merino vs. (Poll Dorset and Dohne) | 41 | RGCC, MVK A, LOC101101919, LOC101121185, TNS3 A, NFYB, MSRA, ZNF706, RRP15, PPARD, MED6 A, LOC101102105, LOC105612466, LMBR1 B, TGFB2 C, ARHGDIA B, MAPK3 A, DHX40 B, LOC114114020, MKLN1 B, PITX1 B, CHST4 B, FCAMR B, CSGALNACT1 A, COMMD3 A, SFN, LOC114115624, RIC8A, PRR14L A, CHPF, BORCS5, LOC101103973, KRT4 C, ITSN1 C, IVL C, GTL2 A, FAM210B B, SPTBN2 A, DEAF1 B, LOC114110979, OBSL1 A |
| Dohne vs. (Poll Dorset and Merino) | 4 | CARMIL1 A, FLRT2 A, ABCA4, ZNF382 |
| Poll Dorset vs. (Dohne and Merino) | 31 | LOC101108654, PAMR1 B, LY6E B, ARGLU1 C, LOC101113735, CDO1 C, TP53I11, MAN1A1 ∇, CHI3L1 A, GAA, SORBS2 B, APOM C, BPIFB1, HEMK1 C, LPO, CDH26 B, NUAK1, UNC5B A, SGSH B, VGF C, GSDMA C, FCGR3A B, HSPB8 B, RNF151 A, AFAP1L2 B, ANKEF1, SLC48A1 C, DHRS3 B, G0S2 B, UBXN8 B, LOC101103862 |
| GO Term | p-Value | Number of Genes |
|---|---|---|
| Molecular Functions | ||
| Identical protein binding | 2.16 × 10−5 | 64 |
| Protein domain specific binding | 1.05 × 10−4 | 28 |
| Cellular Components | ||
| Vacuole | 9.96 × 10−6 | 24 |
| Lysosome | 9.96 × 10−6 | 22 |
| Lytic vacuole | 9.96 × 10−6 | 22 |
| Biological Processes | ||
| Defence response to other organism | 4.98 × 10−8 | 34 |
| Cellular response to cytokine stimulus | 4.98 × 10−8 | 35 |
| Cellular response to organic substance | 4.98 × 10−8 | 67 |
| Cellular response to chemical stimulus | 4.98 × 10−8 | 81 |
| Response to other organism | 4.98 × 10−8 | 49 |
| Regulation of multicellular organismal process | 4.98 × 10−8 | 89 |
| Regulation of immune response | 4.98 × 10−8 | 30 |
| Positive regulation of response to stimulus | 4.98 × 10−8 | 67 |
| Regulation of response to stimulus | 4.98 × 10−8 | 107 |
| Innate immune response | 4.98 × 10−8 | 24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodge, M.J.; de las Heras-Saldana, S.; Rindfleish, S.J.; Stephen, C.P.; Pant, S.D. Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia. Genes 2021, 12, 203. https://doi.org/10.3390/genes12020203
Hodge MJ, de las Heras-Saldana S, Rindfleish SJ, Stephen CP, Pant SD. Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia. Genes. 2021; 12(2):203. https://doi.org/10.3390/genes12020203
Chicago/Turabian StyleHodge, Marnie J., Sara de las Heras-Saldana, Sally J. Rindfleish, Cyril P. Stephen, and Sameer D. Pant. 2021. "Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia" Genes 12, no. 2: 203. https://doi.org/10.3390/genes12020203
APA StyleHodge, M. J., de las Heras-Saldana, S., Rindfleish, S. J., Stephen, C. P., & Pant, S. D. (2021). Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia. Genes, 12(2), 203. https://doi.org/10.3390/genes12020203