
The Genetic Basis of Tomato Aroma




Abstract
:1. Introduction
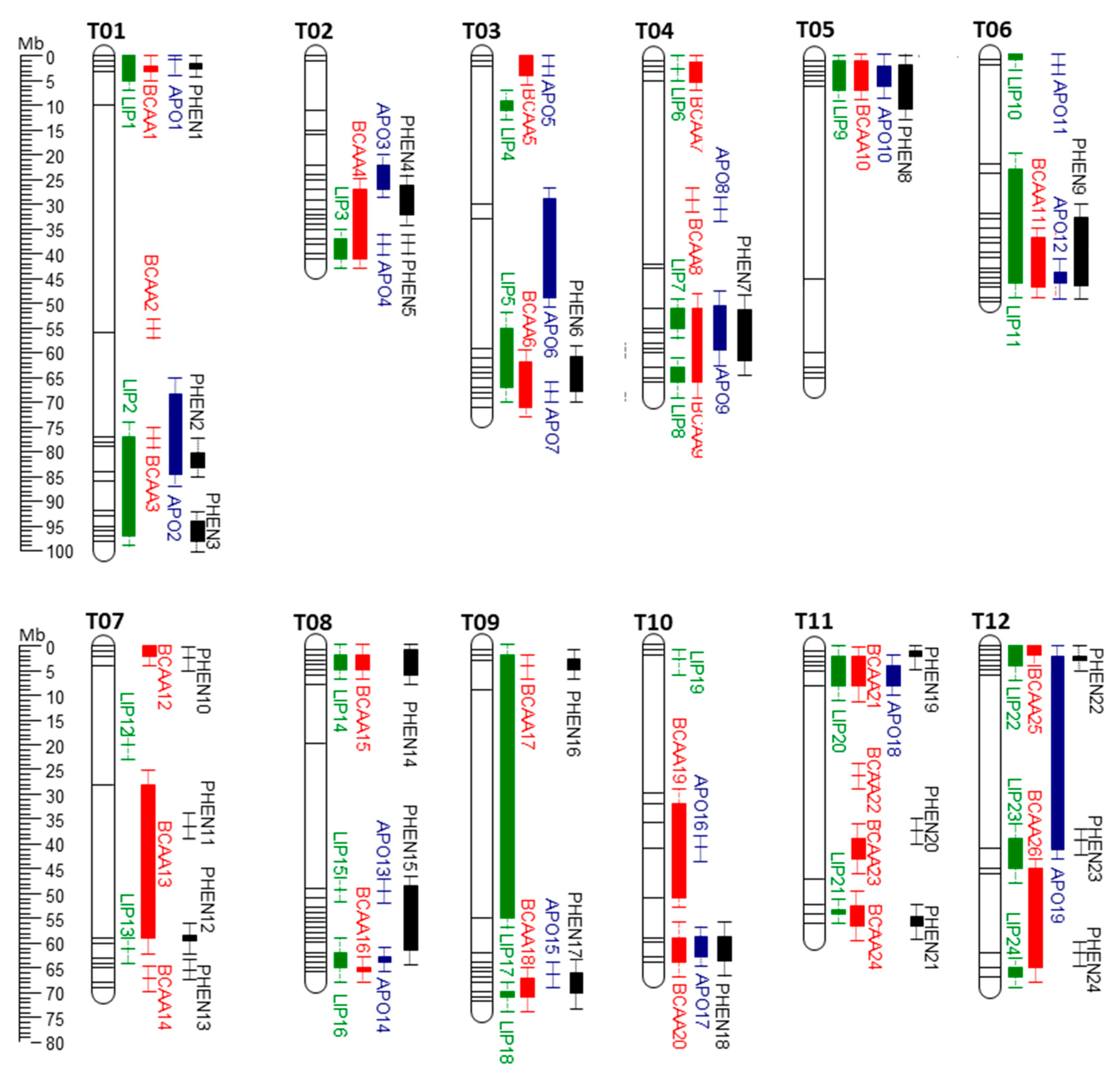
2. Construction of a Unified QTL Map of Tomato Aroma
3. Fatty Acids Derived Volatiles (FA VOCs)
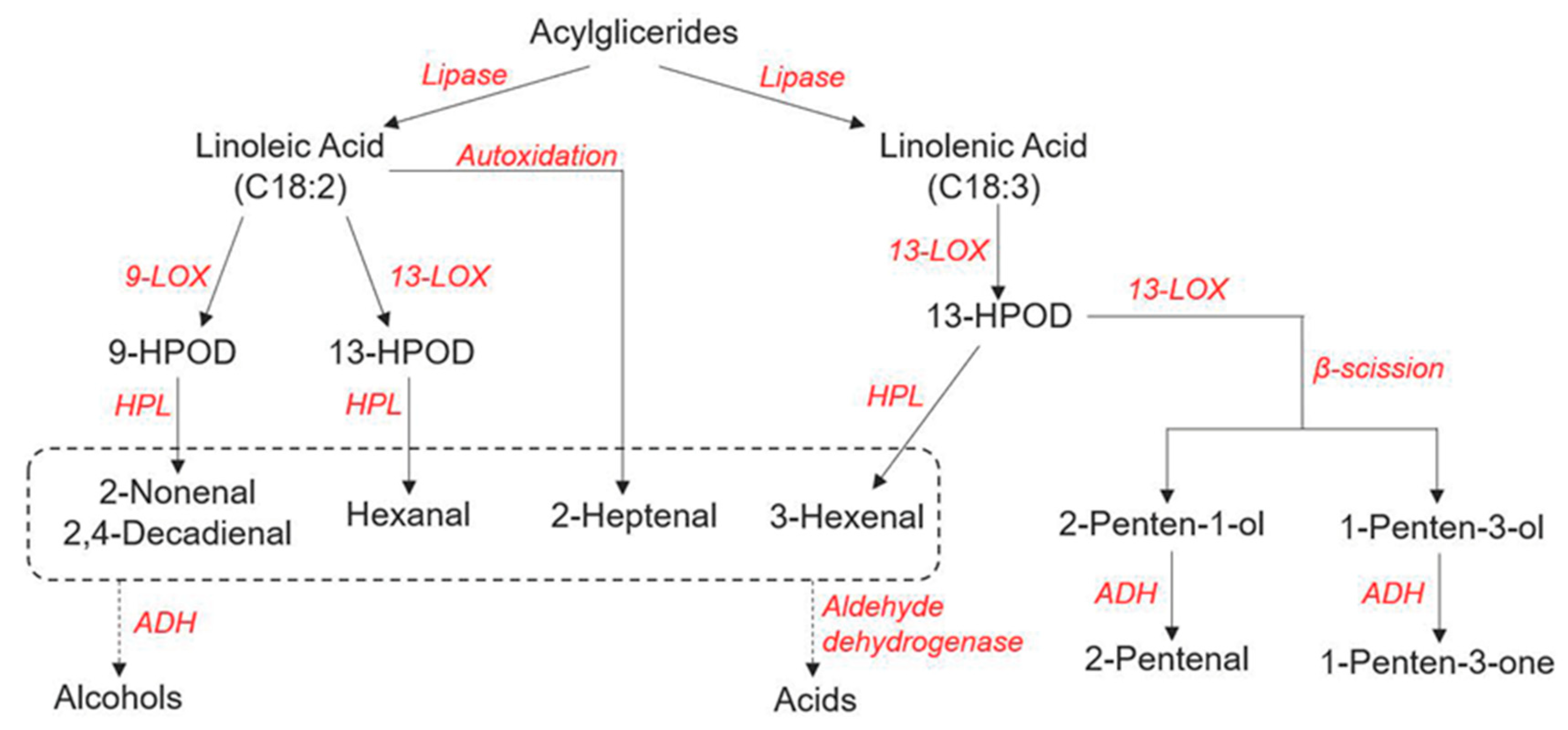
3.1. Biosynthesis of FA VOCs
3.2. QTLs for FA VOCs
3.3. FA VOCs’ Candidate Genes
4. Branched-Chain Amino Acids Derivatives (BCAA VOCs)
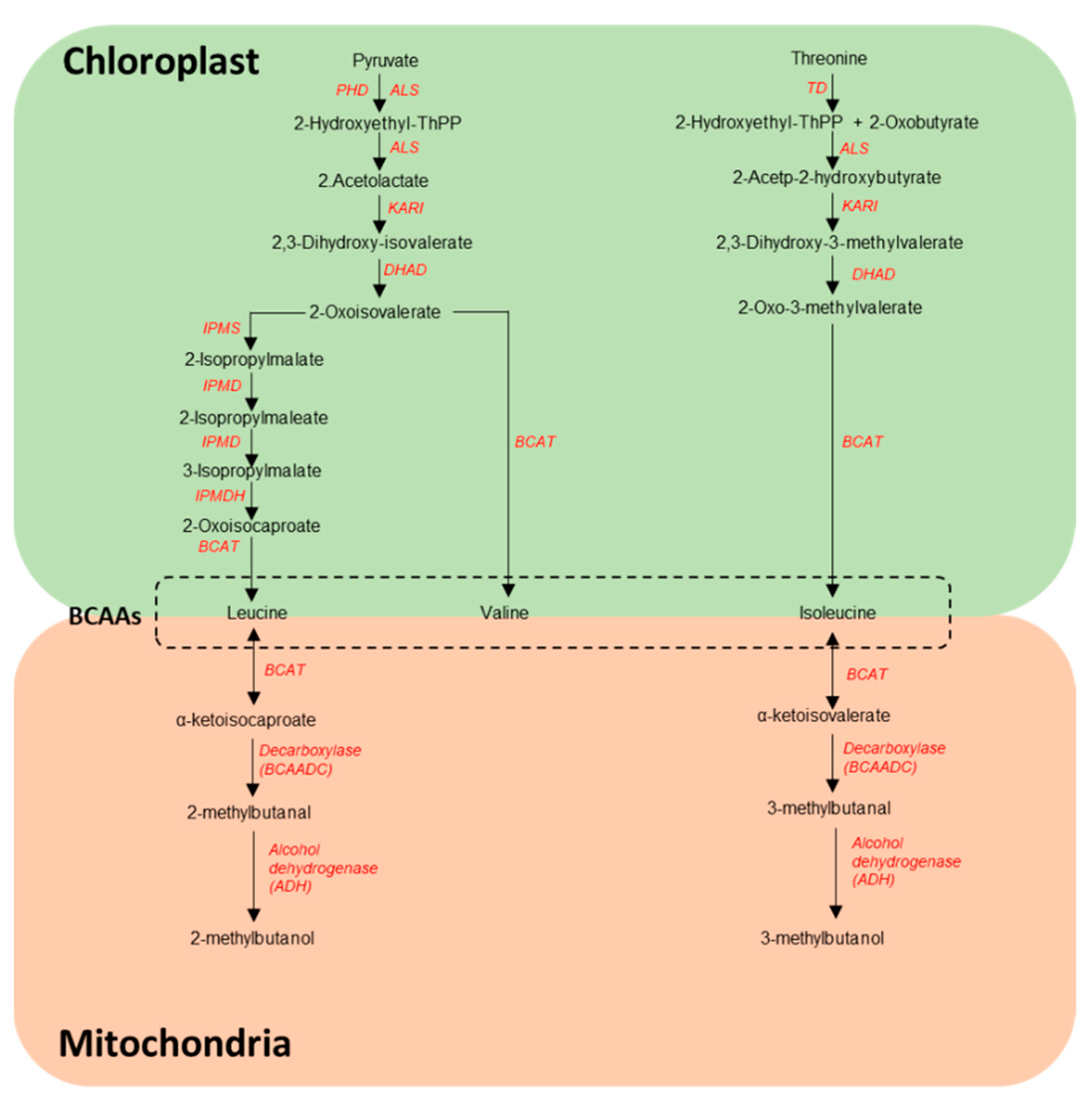
4.1. Biosynthesis of BCAA VOCs
4.2. QTLs for BCAA VOCs
4.3. BCAA VOC Candidate Genes
5. Carotenoid-Derived VOCs
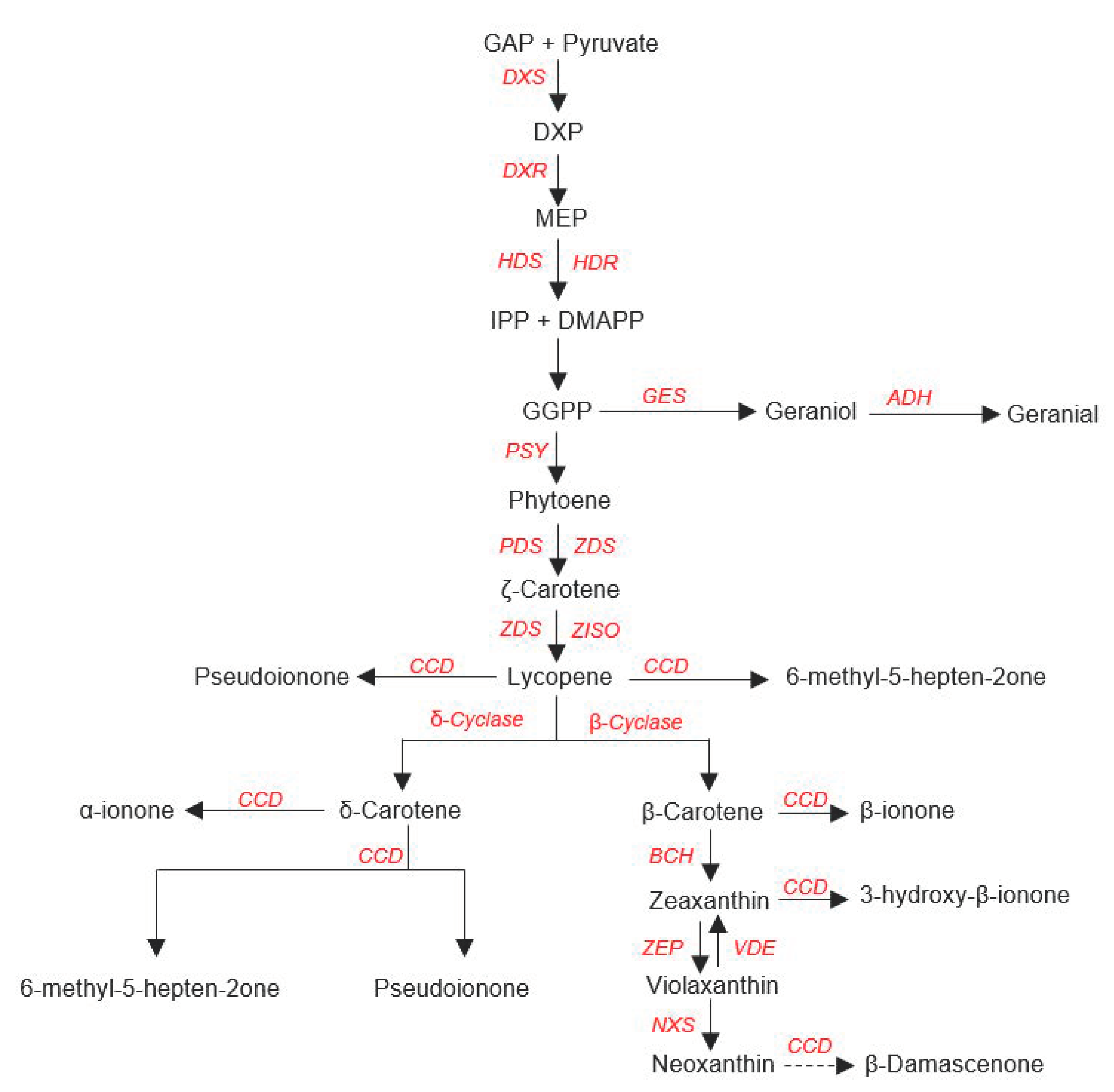
5.1. Biosynthesis
5.2. QTLs for Apocarotenoid VOCs
5.3. Apocarotenoid VOCs’ Candidate Genes
6. Phenylalanine-Derived Volatiles (Phe VOCs)
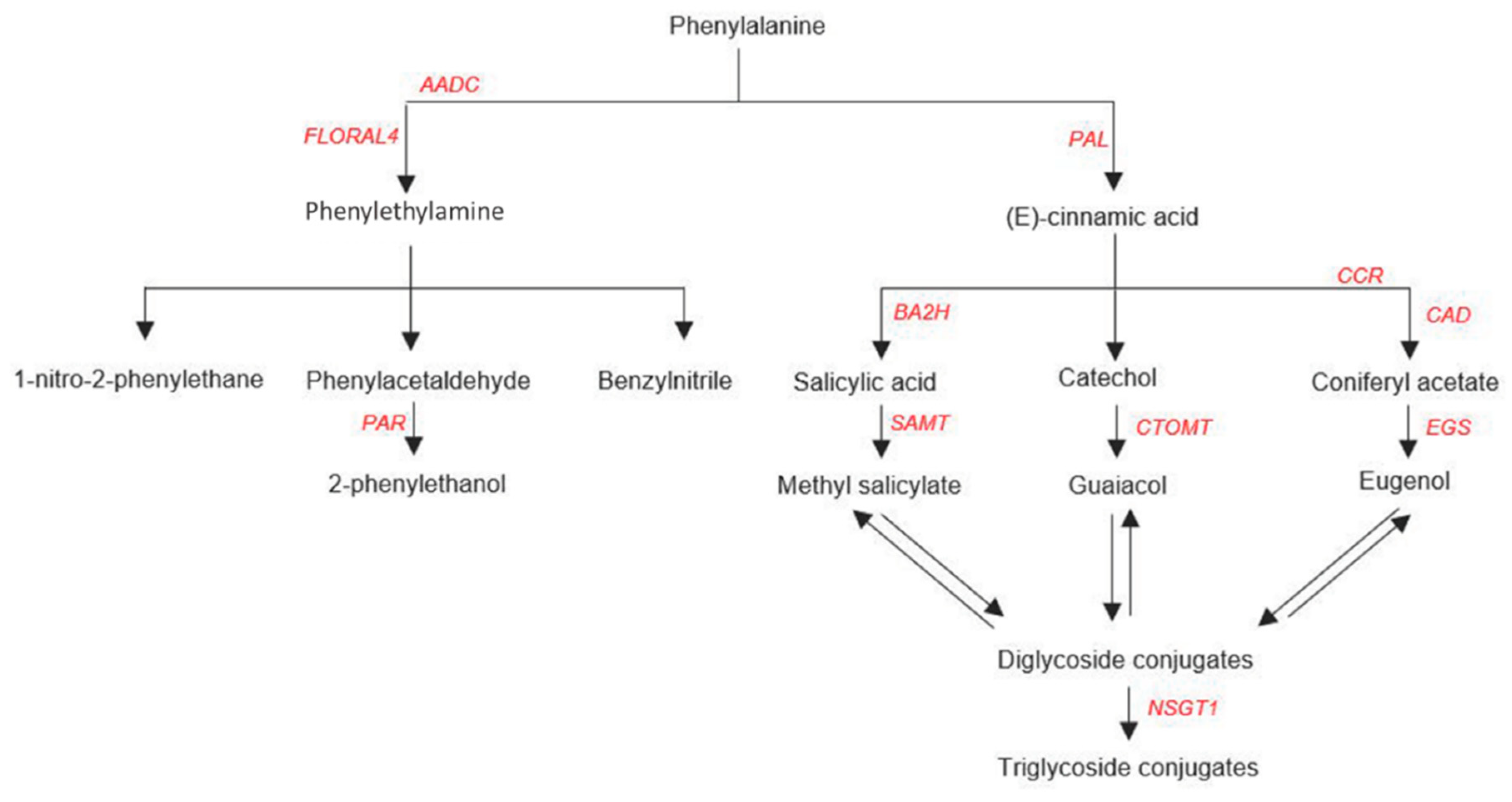
6.1. Biosynthesis of Phe VOCs
6.2. QTLs for Phe VOCs
6.3. Phe VOCs’ Candidate Genes
7. Concluding Remarks
- (i)
- The QTL information presented in this review can be directly used to support marker-assisted breeding programs aimed at introgressing large-effect aroma QTLs into elite germplasm, for example using the donors indicated in Table S5.
- (ii)
- The current development of pangenome projects paves the way for a new step in tomato breeding research. Advances in computational genomics and long-read sequencing allow an easier and more comprehensive investigation of the genetic variation in tomato collections worldwide. This makes it possible to identify genetic elements that are missing in the reference genome and to discover and use novel markers—such as Structural Variants (SVs) and Present Absent Variants (PAVs) [190]. Thanks to the ongoing reduction in sequencing costs, large sets of genotypes can nowadays be re-sequenced, allowing the application of SV markers in GWAS projects [191,192]. Furthermore, SV identification in large sets of genotypes can lead to downstream breeding approaches. For example, SV-based linkage mapping can be applied by genotyping mapping populations using SV markers that showed polymorphism in the parental lines [193]. Furthermore, SV studies may help to get more insight in the mechanisms leading to a certain phenotype [26,65,194,195], pinpointing the best donors for a certain allele cross. By providing a comprehensive set of candidate genes for tomato aroma, our review may guide researchers and breeders in the selection of the most interesting genes that can be investigated for structural variation.
- (iii)
- The data in this review can be used to support the identification and use of the key genes underlying these QTLs. In combination with available (pan)genomic and transcriptomic information candidate genes present in the QTL regions can be selected and tested for their effects in vivo using either stable transgenic approaches such as CRISPR-CAS9 mediated gene editing or quicker transient overexpression or silencing in tomato fruit. This may not only lead to the identification of the causal genes controlling a trait, but also to the detection of the causal genetic variants underlying trait variation. Such variants, also called functional markers, are the best possible molecular markers for MAS, since they are functionally linked to the trait rather than genetically linked and their use as marker does not need validation in other populations, which is always required with genetically-linked markers [196].
- (iv)
- The information on large effect aroma QTLs provided in this review can alo be used to improve the performance of genomic prediction models, since both genetically-linked markers and in particular functional markers have been shown to significantly improve the prediction power of GP models compared to the use of random neutral markers [197].
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peralta, I.; Knapp, S.; Spooner, D.M. New species of wild tomatoes (Solanum section lycopersicon: Solanaceae) from Northern Peru. Syst. Bot. 2005, 30, 424–434. [Google Scholar] [CrossRef]
- Schouten, H.J.; Tikunov, Y.; Verkerke, W.; Finkers, R.; Bovy, A.; Bai, Y.; Visser, R.G. Breeding has increased the diversity of cultivated tomato in The Netherlands. Front. Plant Sci. 2019, 10, 1606. [Google Scholar] [CrossRef] [PubMed]
- Víquez-Zamora, M.; Vosman, B.; Van De Geest, H.; Bovy, A.; Visser, R.G.F.; Finkers, R.; Van Heusden, A.W. Tomato breeding in the genomics era: Insights from a SNP array. BMC Genom. 2013, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfermeijer, F.C.; Dijkhuis, J.; Sturre, M.J.; De Haan, P.; Hille, J. Cloning and characterization of the durable tomato mosaic virus resistance gene Tm-22 from Lycopersicon esculentum. Plant Mol. Biol. 2003, 52, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Zhu, G.; Zhang, J.; Xu, X.; Yu, Q.; Zheng, Z.; Zhang, Z.; Lun, Y.; Li, S.; Wang, X.; et al. Genomic analyses provide insights into the history of tomato breeding. Nat. Genet. 2014, 46, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, S.; Cin, V.D.; Fei, Z.; Li, H.; Bliss, P.; Taylor, M.G.; Klee, H.J.; Tieman, D.M. Flavour compounds in tomato fruits: Identification of loci and potential pathways affecting volatile composition. J. Exp. Bot. 2008, 60, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Petro-Turza, M. Flavor of tomato and tomato products. Food Rev. Int. 1986, 2, 309–351. [Google Scholar] [CrossRef]
- Tieman, D.; Bliss, P.; McIntyre, L.M.; Blandon-Ubeda, A.; Bies, D.; Odabasi, A.Z.; Rodríguez, G.R.; Van Der Knaap, E.; Taylor, M.G.; Goulet, C.; et al. The chemical interactions underlying tomato flavor preferences. Curr. Biol. 2012, 22, 1035–1039. [Google Scholar] [CrossRef] [Green Version]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of Plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Buttery, R.G.; Teranishi, R.; Flath, R.A.; Ling, L.C. Fresh Tomato Volatiles: Composition and Sensory Studies. In Flavour Chemistry; Teranishi, R., Buttery, R.G., Shahidi, F., Eds.; American Chemical Society: Washington, DC, USA, 1989; Volume 388, pp. 213–222. [Google Scholar]
- Rambla, J.L.; Tikunov, Y.M.; Monforte, A.J.; Bovy, A.G.; Granell, A. The expanded tomato fruit volatile landscape. J. Exp. Bot. 2013, 65, 4613–4623. [Google Scholar] [CrossRef] [Green Version]
- Tandon, K.; Baldwin, E.A.; Scott, J.; Shewfelt, R. Linking sensory descriptors to volatile and nonvolatile components of fresh tomato flavor. J. Food Sci. 2003, 68, 2366–2371. [Google Scholar] [CrossRef]
- Baldwin, E.A.; Goodner, K.; Plotto, A. Interaction of volatiles, sugars, and acids on perception of tomato aroma and flavor descriptors. J. Food Sci. 2008, 73, S294–S307. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, E.A.; Goodner, K.L.; Plotto, A.; Pritchett, K.; Einstein, M.A. Effect of volatiles and their concentration on perception of tomato descriptors. J. Food Sci. 2004, 69, S310–S318. [Google Scholar] [CrossRef]
- Vogel, J.T.; Tieman, D.M.; Sims, C.A.; Odabasi, A.Z.; Clark, D.G.; Klee, H.J. Carotenoid content impacts flavor acceptability in tomato (Solanum lycopersicum). J. Sci. Food Agric. 2010, 90, 2233–2240. [Google Scholar] [CrossRef]
- Abegaz, E.; Tandon, K.; Scott, J.; Baldwin, E.; Shewfelt, R. Partitioning taste from aromatic flavor notes of fresh tomato (Lycopersicon esculentum, Mill) to develop predictive models as a function of volatile and nonvolatile components. Postharvest Biol. Technol. 2004, 34, 227–235. [Google Scholar] [CrossRef]
- Piombino, P.; Sinesio, F.; Moneta, E.; Cammareri, M.; Genovese, A.; Lisanti, M.T.; Mogno, M.R.; Peparaio, M.; Termolino, P.; Moio, L.; et al. Investigating physicochemical, volatile and sensory parameters playing a positive or a negative role on tomato liking. Food Res. Int. 2013, 50, 409–419. [Google Scholar] [CrossRef] [Green Version]
- The Good Scents Company-Flavour, Fragrance, Food and Cosmetics Ingredients Information. Available online: http://www.thegoodscentscompany.com/ (accessed on 26 November 2020).
- Buttery, R.G.; Teranishi, R.; Ling, L.C. Fresh tomato aroma volatiles: A quantitative study. J. Agric. Food Chem. 1987, 35, 540–544. [Google Scholar] [CrossRef]
- Buttery, R.G.; Teranishi, R.; Ling, L.C.; Flath, R.A.; Stern, D.J. Quantitative studies on origins of fresh tomato aroma volatiles. J. Agric. Food Chem. 1988, 36, 1247–1250. [Google Scholar] [CrossRef]
- Baldwin, E.A.; Scott, J.W.; Shewmaker, C.K.; Schuch, W. Flavor trivia and tomato aroma: Biochemistry and possible mechanisms for control of important aroma components. HortScience 2000, 35, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Tandon, K.; Baldwin, E.; Shewfelt, R. Aroma perception of individual volatile compounds in fresh tomatoes (Lycopersicon esculentum, Mill.) as affected by the medium of evaluation. Postharvest Biol. Technol. 2000, 20, 261–268. [Google Scholar] [CrossRef]
- Tandon, K.S.; Baldwin, E.A.; Goodner, K.L.; Jordán, M. Characterization of fresh tomato aroma volatiles using GC-Olfatometry. In Proceedings of the Annual Meeting of the Florida State Horticultural Society, Stuart, FL, USA, 10–12 June 2001. [Google Scholar]
- Selli, S.; Kelebek, H.; Ayseli, M.T.; Tokbas, H. Characterization of the most aroma-active compounds in cherry tomato by application of the aroma extract dilution analysis. Food Chem. 2014, 165, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Tieman, D.; Zhu, G.; Resende, M.F.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavor. Science 2017, 355, 391–394. [Google Scholar] [CrossRef]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Burzynski-Chang, E.A.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L.; et al. The tomato pan-genome uncovers new genes and a rare allele regulating fruit flavor. Nat. Genet. 2019, 51, 1044–1051. [Google Scholar] [CrossRef]
- Gur, A.; Semel, Y.; Osorio, S.; Friedmann, M.; Seekh, S.; Ghareeb, B.; Mohammad, A.; Pleban, T.; Gera, G.; Fernie, A.R.; et al. Yield quantitative trait loci from wild tomato are predominately expressed by the shoot. Theor. Appl. Genet. 2010, 122, 405–420. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Alseekh, S.; Brotman, Y.; Zheng, Y.; Fei, Z.; Tieman, D.M.; Giovannoni, J.J.; Fernie, A.R.; Klee, H.J. Identification of a Solanum pennellii chromosome 4 fruit flavor and nutritional quality-associated metabolite QTL. Front. Plant Sci. 2016, 7, 1671. [Google Scholar] [CrossRef] [Green Version]
- Eshed, Y.; Zamir, D. An introgression line population of lycopersicon pennellii in the cultivated tomato enables the identi-fication and fine mapping of yield-associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [CrossRef]
- Fridman, E.; Carrari, F.; Liu, Y.-S.; Fernie, A.R.; Zamir, D. Zooming in on a quantitative trait for tomato yield using interspecific introgressions. Science 2004, 305, 1786–1789. [Google Scholar] [CrossRef] [PubMed]
- Canady, M.A.; Meglic, V.; Chetelat, R.T. A library of Solanum lycopersicoides introgression lines in cultivated tomato. Genome 2005, 48, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Barrantes, W.; López-Casado, G.; García-Martínez, S.; Alonso, A.; Rubio, F.; Ruiz, J.J.; Fernández-Muñoz, R.; Granell, A.; Monforte, A. Exploring new alleles involved in tomato fruit quality in an introgression line library of Solanum pimpinellifolium. Front. Plant Sci. 2016, 7, 1172. [Google Scholar] [CrossRef] [Green Version]
- Rambla, J.L.; Medina, A.; Fernández-Del-Carmen, A.; Barrantes, W.; Grandillo, S.; Cammareri, M.; López-Casado, G.; Rodrigo, G.; Alonso, A.; García-Martínez, S.; et al. Identification, introgression, and validation of fruit volatile QTLs from a red-fruited wild tomato species. J. Exp. Bot. 2016, 68, 429–442. [Google Scholar] [CrossRef]
- Blanca, J.; Montero-Pau, J.; Sauvage, C.; Bauchet, G.; Illa-Berenguer, E.; Díez, M.J.; Francis, D.M.; Causse, M.; Van Der Knaap, E.; Canizares, J. Genomic variation in tomato, from wild ancestors to contemporary breeding accessions. BMC Genom. 2015, 16, 257. [Google Scholar] [CrossRef] [Green Version]
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Ozminkowski, R.; Ozminkowski, R. Pedigree of variety heinz 1706. Rep. Tomato Genet. Coop. 2004, 54, 26. [Google Scholar]
- Hamilton, J.P.; Sim, S.-C.; Stoffel, K.; Van Deynze, A.; Buell, C.R.; Francis, D.M. Single nucleotide polymorphism discovery in cultivated tomato via sequencing by synthesis. Plant Genome 2012, 5, 17–29. [Google Scholar] [CrossRef]
- Sim, S.-C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; Van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a large SNP genotyping array and generation of high-density genetic maps in tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Celik, I.; Gurbuz, N.; Uncu, A.T.; Frary, A.; Doganlar, S. Genome-wide SNP discovery and QTL mapping for fruit quality traits in inbred backcross lines (IBLs) of Solanum pimpinellifolium using genotyping by sequencing. BMC Genom. 2017, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ohlson, E.W.; Ashrafi, H.; Foolad, M.R. Identification and mapping of late blight resistance quantitative trait loci in tomato accession PI 163245. Plant Genome 2018, 11, 180007. [Google Scholar] [CrossRef] [Green Version]
- Gonda, I.; Ashrafi, H.; Lyon, D.A.; Strickler, S.R.; Hulse-Kemp, A.M.; Ma, Q.; Sun, H.; Stoffel, K.; Powell, A.F.; Futrell, S.; et al. Sequencing-based bin map construction of a tomato mapping population, facilitating high-resolution quantitative trait loci detection. Plant Genome 2019, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-P.; Liu, C.-Y.; Chen, K.-Y. Assessment of genetic differentiation and linkage disequilibrium in Solanum pimpinellifolium using genome-wide high-density SNP markers. G3 Genes Genomes Genet. 2019, 9, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Acquadro, A.; Barchi, L.; Gramazio, P.; Portis, E.; Vilanova, S.; Comino, C.; Plazas, M.; Prohens, J.; Lanteri, S. Coding SNPs analysis highlights genetic relationships and evolution pattern in eggplant complexes. PLoS ONE 2017, 12, e0180774. [Google Scholar] [CrossRef] [Green Version]
- Edae, E.A.; Olivera, P.D.; Jin, Y.; Rouse, M.N. Genotyping-by-sequencing facilitates a high-density consensus linkage map forAegilops umbellulata, a wild relative of cultivated wheat. G3 Genes Genomes Genet. 2017, 7, 1551–1561. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Cericola, F.; Fè, D.; Pedersen, M.G.; Lenk, I.; Jensen, C.S.; Jensen, J.; Janss, L.L. Genomic prediction in tetraploid ryegrass using allele frequencies based on genotyping by sequencing. Front. Plant Sci. 2018, 9, 1165. [Google Scholar] [CrossRef] [Green Version]
- Hussain, W.; Baenziger, P.S.; Belamkar, V.; Guttieri, M.J.; Venegas, J.P.; Easterly, A.; Sallam, A.; Poland, J. Genotyping-by-sequencing derived high-density linkage map and its application to QTL mapping of flag leaf traits in bread wheat. Sci. Rep. 2017, 7, 16394. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Chen, G.; Song, C.; Keefe, J.; Mendelson, M.; Huan, T.; Sun, B.B.; Laser, A.; Maranville, J.C.; Wu, H.; et al. Genome-wide mapping of plasma protein QTLs identifies putatively causal genes and pathways for cardiovascular disease. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Jin, Y.; Zhao, W.; Nie, S.; Liu, S.-S.; El-Kassaby, Y.A.; Wang, X.-R.; Mao, J.-F. Genome-wide variant identification and high-density genetic map construction using RADseq for Platycladus orientalis (Cupressaceae). G3 Genes Genomes Genet. 2019, 9, 3663–3672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, P.; Eudy, D.; Schnable, J.C.; Schmutz, J.; Raymer, P.L.; Devos, K.M. High density genetic maps of Seashore paspalum using genotyping-by-sequencing and their relationship to the sorghum bicolor genome. Sci. Rep. 2019, 9, 12183. [Google Scholar] [CrossRef]
- Toppino, L.; Barchi, L.; Mercati, F.; Acciarri, N.; Perrone, D.; Martina, M.; Gattolin, S.; Sala, T.; Fadda, S.; Mauceri, A.; et al. A new intra-specific and high-resolution genetic map of eggplant based on a RIL population, and location of QTLs related to Plant anthocyanin pigmentation and seed vigour. Genes 2020, 11, 745. [Google Scholar] [CrossRef]
- Jordon-Thaden, I.E.; Beck, J.B.; Rushworth, C.A.; Windham, M.D.; Diaz, N.; Cantley, J.T.; Martine, C.T.; Rothfels, C.J. A basic dd RAD seq two-enzyme protocol performs well with herbarium and silica-dried tissues across four genera. Appl. Plant Sci. 2020, 8, e11344. [Google Scholar] [CrossRef] [Green Version]
- Sunde, J.; Yıldırım, Y.; Tibblin, P.; Forsman, A. Comparing the performance of microsatellites and RADseq in population genetic studies: Analysis of data for pike (Esox lucius) and a synthesis of previous studies. Front. Genet. 2020, 11, 218. [Google Scholar] [CrossRef] [Green Version]
- Tsujimoto, M.; Araki, K.S.; Honjo, M.N.; Yasugi, M.; Nagano, A.J.; Akama, S.; Hatakeyama, M.; Shimizu-Inatsugi, R.; Sese, J.; Shimizu, K.K.; et al. Genet assignment and population structure analysis in a clonal forest-floor herb, Cardamine leucantha, using RAD-seq. AoB Plants 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Zhao, S.; Li, M.; Zhang, C.; Qu, H.; Li, Q.; Li, J.; Lin, Y.; Pu, Z. Genome-wide genetic diversity detection and population structure analysis in sweetpotato (Ipomoea batatas) using RAD-seq. Genomics 2020, 112, 1978–1987. [Google Scholar] [CrossRef]
- Aflitos, S.A.; Schijlen, E.; De Jong, H.; De Ridder, D.; Smit, S.; Finkers, R.; Wang, J.; Zhang, G.; Li, N.; Mao, L.; et al. Exploring genetic variation in the tomato (Solanumsection Lycopersicon) clade by whole-genome sequencing. Plant J. 2014, 80, 136–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roohanitaziani, R.; De Maagd, R.A.; Lammers, M.; Molthoff, J.; Meijer-Dekens, F.; Van Kaauwen, M.P.W.; Finkers, R.; Tikunov, Y.; Visser, R.G.F.; Bovy, A. Exploration of a resequenced tomato core collection for phenotypic and genotypic variation in Plant growth and fruit quality traits. Genes 2020, 11, 1278. [Google Scholar] [CrossRef]
- Hirsch, C.N.; Foerster, J.M.; Johnson, J.M.; Sekhon, R.S.; Muttoni, G.; Vaillancourt, B.; Peñagaricano, F.; Lindquist, E.; Pedraza, M.A.; Barry, K.; et al. Insights into the maize pan-genome and pan-transcriptome. Plant Cell 2014, 26, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-H.; Zhou, G.; Ma, J.; Jiang, W.; Jin, L.-G.; Zhang, Z.; Guo, Y.; Zhang, J.; Sui, Y.; Zheng, L.; et al. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat. Biotechnol. 2014, 32, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Schatz, M.C.; Maron, L.G.; Stein, J.C.; Wences, A.H.; Gurtowski, J.; Biggers, E.; Lee, H.; Kramer, M.; Antoniou, E.; Ghiban, E.; et al. Whole genome de novo assemblies of three divergent strains of rice, Oryza sativa, document novel gene space of aus and indica. Genome Biol. 2014, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Golicz, A.A.; Bayer, P.E.; Barker, G.C.; Edger, P.P.; Kim, H.; Martinez, P.A.; Chan, C.-K.K.; Severn-Ellis, A.; McCombie, W.R.; Parkin, I.A.P.; et al. The pangenome of an agronomically important crop Plant Brassica oleracea. Nat. Commun. 2016, 7, 13390. [Google Scholar] [CrossRef]
- Song, J.-M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef]
- Khan, A.W.; Garg, V.; Roorkiwal, M.; Golicz, A.A.; Edwards, D.; Varshney, R.K. Super-pangenome by integrating the wild side of a species for accelerated crop improvement. Trends Plant Sci. 2020, 25, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major impacts of widespread structural variation on gene expression and crop improvement in tomato. Cell 2020, 182, 145–161.e23. [Google Scholar] [CrossRef]
- Ou, L.; Li, D.; Lv, J.; Chen, W.; Zhang, Z.; Li, X.; Yang, B.; Zhou, S.; Yang, S.; Li, W.; et al. Pan-genome of cultivated pepper (Capsicum) and its use in gene presence-absence variation analyses. New Phytol. 2018, 220, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Hackett, R.; Walker, D.; Taylor, A.; Lin, Z.; Grierson, D. Identification of a specific isoform of tomato lipoxygenase (TomloxC) involved in the generation of fatty acid-derived flavor compounds. Plant Physiol. 2004, 136, 2641–2651. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Tieman, D.; Jones, J.B.; Taylor, M.G.; Schmelz, E.; Huffaker, A.; Bies, D.; Chen, K.; Klee, H.J. A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 flavour volatiles in tomato. J. Exp. Bot. 2014, 65, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Guo, Z.; Huang, C.; Duan, L.; Chen, G.; Jiang, N.; Fang, W.; Feng, H.; Xie, W.; Lian, X.; et al. Combining high-throughput phenotyping and genome-wide association studies to reveal natural genetic variation in rice. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jiang, H.; Yeh, C.; Yu, J.; Jeddeloh, J.A.; Nettleton, D.; Schnable, P.S. Extreme-phenotype genome-wide association study (XP-GWAS): A method for identifying trait-associated variants by sequencing pools of individuals selected from a diversity panel. Plant J. 2015, 84, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Minamikawa, M.F.; Takada, N.; Terakami, S.; Saito, T.; Onogi, A.; Kajiya-Kanegae, H.; Hayashi, T.; Yamamoto, T.; Iwata, H. Genome-wide association study and genomic prediction using parental and breeding populations of Japanese pear (Pyrus pyrifolia Nakai). Sci. Rep. 2018, 8, 11994. [Google Scholar] [CrossRef] [Green Version]
- Mir, R.R.; Reynolds, M.; Pinto, F.; Khan, M.A.; Bhat, M.A. High-throughput phenotyping for crop improvement in the genomics era. Plant Sci. 2019, 282, 60–72. [Google Scholar] [CrossRef]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS discovery: Biology, function, and translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvage, C.; Segura, V.; Bauchet, G.; Stevens, R.; Do, P.T.; Nikoloski, Z.; Fernie, A.R.; Causse, M. Genome-wide association in tomato reveals 44 candidate loci for fruit metabolic traits. Plant Physiol. 2014, 165, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhao, J.; Xu, Y.; Liang, J.; Chang, P.; Yan, F.; Li, M.; Liang, Y.; Zou, Z. Genome-wide association mapping for tomato volatiles positively contributing to tomato flavor. Front. Plant Sci. 2015, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Bauchet, G.; Grenier, S.; Samson, N.; Bonnet, J.; Grivet, L.; Causse, M. Use of modern tomato breeding germplasm for deciphering the genetic control of agronomical traits by genome wide association study. Theor. Appl. Genet. 2017, 130, 875–889. [Google Scholar] [CrossRef]
- Zhao, J.; Sauvage, C.; Zhao, J.; Bitton, F.; Bauchet, G.; Liu, D.; Huang, S.; Tieman, D.M.; Klee, H.J.; Causse, M. Meta-analysis of genome-wide association studies provides insights into genetic control of tomato flavor. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, N.T.; Trinh, L.T.; Rho, M.-Y.; Park, T.-S.; Kim, O.-R.; Zhao, J.; Kim, H.-M.; Sim, S.-C. Identification of loci associated with fruit traits using genome-wide single nucleotide polymorphisms in a core collection of tomato (Solanum lycopersicum L.). Sci. Hortic. 2019, 243, 567–574. [Google Scholar] [CrossRef]
- Ye, J.; Li, W.; Ai, G.; Li, C.; Liu, G.; Chen, W.; Wang, B.; Wang, W.; Lu, Y.; Zhang, J.; et al. Genome-wide association analysis identifies a natural variation in basic helix-loop-helix transcription factor regulating ascorbate biosynthesis via D-mannose/L-galactose pathway in tomato. PLoS Genet. 2019, 15, e1008149. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Teng, J.; Ye, S.; Yuan, X.; Huang, S.; Zhang, H.; Zhang, X.; Li, J.; Zhang, Z. Genomic prediction of complex phenotypes using genic similarity based relatedness matrix. Front. Genet. 2018, 9, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daetwyler, H.D.; Calus, M.P.L.; Pong-Wong, R.; Campos, G.D.L.; Hickey, J.M. Genomic prediction in animals and plants: Simulation of data, validation, reporting, and benchmarking. Genetics 2013, 193, 347–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Kajiya-Kanegae, H.; Minamikawa, M.; Suzuki, A.; Shirasawa, K.; Hirakawa, H.; Nunome, T.; Yamaguchi, H.; et al. A simulation-based breeding design that uses whole-genome prediction in tomato. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Tusell, L.; Pérez-Rodríguez, P.; Forni, S.; Wu, X.-L.; Gianola, D. Genome-enabled methods for predicting litter size in pigs: A comparison. Animal 2013, 7, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Ni, G.; Cavero, D.; Fangmann, A.; Erbe, M.; Simianer, H. Whole-genome sequence-based genomic prediction in laying chickens with different genomic relationship matrices to account for genetic architecture. Genet. Sel. Evol. 2017, 49, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duenk, P.; Calus, M.P.L.; Wientjes, Y.C.J.; Breen, V.P.; Henshall, J.; Hawken, R.J.; Bijma, P. Validation of genomic predictions for body weight in broilers using crossbred information and considering breed-of-origin of alleles. Genet. Sel. Evol. 2019, 51, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.K.; Chaudhari, S.; Jarquin, D.; Janila, P.; Crossa, J.; Patil, S.C.; Sundravadana, S.; Khare, D.; Bhat, R.S.; Radhakrishnan, T.; et al. Genome-based trait prediction in multi-environment breeding trials in groundnut. Theor. Appl. Genet. 2020, 133, 3101–3117. [Google Scholar] [CrossRef]
- Lenz, P.; Nadeau, S.; Azaiez, A.; Gérardi, S.; DesLauriers, M.; Perron, M.; Isabel, N.; Beaulieu, J.; Bousquet, J. Genomic prediction for hastening and improving efficiency of forward selection in conifer polycross mating designs: An example from white spruce. Heredity 2020, 124, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Robertsen, C.D.; Hjortshøj, R.L.; Janss, L. Genomic selection in cereal breeding. Agronomy 2019, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Cortés, A.J.; Restrepo-Montoya, M.; Bedoya-Canas, L.E. Modern strategies to assess and breed forest tree adaptation to changing climate. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Saliba-Colombani, V.; Causse, M.; Langlois, D.; Philouze, J.; Buret, M. Genetic analysis of organoleptic quality in fresh market tomato. 1. Mapping QTLs for physical and chemical traits. Theor. Appl. Genet. 2001, 102, 259–272. [Google Scholar] [CrossRef]
- Fulton, T.; Bucheli, P.; Voirol, E.; Lopez, J.; Pétiard, V.; Tanksley, S. Quantitative trait loci (QTL) affecting sugars, organic acids and other biochemical properties possibly contributing to flavor, identified in four advanced backcross populations of tomato. Euphytica 2002, 127, 163–177. [Google Scholar] [CrossRef]
- Tadmor, Y.; Fridman, E.; Gur, A.; Larkov, O.; Lastochkin, E.; Ravid, U.; Zamir, D.; Lewinsohn, E. Identification of malodorous, a wild species allele affecting tomato aroma that was selected against during domestication. J. Agric. Food Chem. 2002, 50, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- Tieman, D.M.; Zeigler, M.; Schmelz, E.A.; Taylor, M.G.; Bliss, P.; Kirst, M.; Klee, H.J. Identification of loci affecting flavour volatile emissions in tomato fruits. J. Exp. Bot. 2006, 57, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Kochevenko, A.; Fernie, A.R. The genetic architecture of branched-chain amino acid accumulation in tomato fruits. J. Exp. Bot. 2011, 62, 3895–3906. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, V.; Francese, G.; Sacco, A.; D’Alessandro, A.; Rigano, M.M.; Parisi, M.; Milone, M.; Cardi, T.; Mennella, G.; Barone, A.; et al. An association mapping approach to identify favourable alleles for tomato fruit quality breeding. BMC Plant Biol. 2014, 14, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Capel, C.; Fernández-Del-Carmen, A.; Alba, J.M.; Lima-Silva, V.; Gras, F.H.; Salinas, M.; Boronat, A.; Angosto, T.; Botella, J.R.; Fernández-Muñoz, R.; et al. Wide-genome QTL mapping of fruit quality traits in a tomato RIL population derived from the wild-relative species Solanum pimpinellifolium L. Theor. Appl. Genet. 2015, 128, 2019–2035. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, Y.; Ding, Q.; Huang, X.; Zhang, Y.; Zou, Z.; Li, M.; Cui, L.; Zhang, J. Association mapping of main tomato fruit sugars and organic acids. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Ebaldina, S.; Picarella, M.E.; Troise, A.D.; Epucci, A.; Eruggieri, V.; Eferracane, R.; Ebarone, A.; Efogliano, V.; Mazzucato, A. Metabolite profiling of italian tomato landraces with different fruit types. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Bauchet, G.; Grenier, S.; Samson, N.; Segura, V.; Kende, A.; Beekwilder, J.; Cankar, K.; Gallois, J.; Gricourt, J.; Bonnet, J.; et al. Identification of major loci and genomic regions controlling acid and volatile content in tomato fruit: Implications for flavor improvement. New Phytol. 2017, 215, 624–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbowicz, K.; Liu, Z.; Alseekh, S.; Tieman, D.; Taylor, M.; Kuhalskaya, A.; Ofner, I.; Zamir, D.; Klee, H.J.; Fernie, A.R.; et al. Quantitative trait loci analysis identifies a prominent gene involved in the production of fatty acid-derived flavor volatiles in tomato. Mol. Plant 2018, 11, 1147–1165. [Google Scholar] [CrossRef] [Green Version]
- Kimbara, J.; Ohyama, A.; Chikano, H.; Ito, H.; Hosoi, K.; Negoro, S.; Miyatake, K.; Yamaguchi, H.; Nunome, T.; Fukuoka, H.; et al. QTL mapping of fruit nutritional and flavor components in tomato (Solanum lycopersicum) using genome-wide SSR markers and recombinant inbred lines (RILs) from an intra-specific cross. Euphytica 2018, 214, 210. [Google Scholar] [CrossRef]
- Tikunov, Y.; Roohanitaziani, R.; Meijer-Dekens, F.; Molthoff, J.; Paulo, J.; Finkers, R.; Capel, I.; Moreno, F.C.; Maliepaard, C.; Vries, M.N.; et al. The genetic and functional analysis of flavor in commercial tomato: The FLORAL4 gene underlies a QTL for floral aroma volatiles in tomato fruit. Plant J. 2020, 103, 1189–1204. [Google Scholar] [CrossRef]
- Zouine, M.; Maza, E.; Djari, A.; Lauvernier, M.; Frasse, P.; Smouni, A.; Pirrello, J.; Bouzayen, M. TomExpress, a unified tomato RNA-Seq platform for visualization of expression data, clustering and correlation networks. Plant J. 2017, 92, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, R.; Karl, T.; Hansel, A.; Jordan, A.; Lindinger, W. Volatile organic compounds emitted after leaf wounding: On-line analysis by proton-transfer-reaction mass spectrometry. J. Geophys. Res. Space Phys. 1999, 104, 15963–15974. [Google Scholar] [CrossRef]
- Ferrie, B. The cloning of two tomato lipoxygenase genes and their differential expression during fruit ripening. Plant Physiol. 1994, 106, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Jansen, G.A.; Wanders, R.J. α-Oxidation. Biochim. Biophys. Acta BBA Bioenerg. 2006, 1763, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Schilmiller, A.L.; Liu, G.; Lee, G.I.; Jayanty, S.; Sageman, C.; Vrebalov, J.; Giovannoni, J.J.; Yagi, K.; Kobayashi, Y.; et al. Role of β-Oxidation in Jasmonate biosynthesis and systemic wound signaling in tomato. Plant Cell 2005, 17, 971–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liavonchanka, A.; Feussner, I. Lipoxygenases: Occurrence, functions and catalysis. J. Plant Physiol. 2006, 163, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Prestage, S.; Linforth, R.S.; Taylor, A.; Lee, E.; Speirs, J.; Schuch, W. Volatile production in tomato fruit with modified alcohol dehydrogenase activity. J. Sci. Food Agric. 1999, 79, 131–136. [Google Scholar] [CrossRef]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef]
- Chen, H.; Cao, S.; Jin, Y.; Tang, Y.; Qi, H. The relationship between CmADHs and the diversity of volatile organic compounds of three aroma types of melon (Cucumis melo). Front. Physiol. 2016, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Vieira, D.D.S.S.; Emiliani, G.; Michelozzi, M.; Centritto, M.; Luro, F.; Morillon, R.; Loreto, F.; Gesteira, A.S.; Maserti, B. Polyploidization alters constitutive content of volatile organic compounds (VOC) and improves membrane stability under water deficit in Volkamer lemon (Citrus limonia Osb.) leaves. Environ. Exp. Bot. 2016, 126, 1–9. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, Q.; Yao, Z.; Deng, H.; Liu, B.; Yue, C.; Deng, T.; Lai, Z.; Sun, Y. Dynamics of ADH and related genes responsible for the transformation of C 6-aldehydes to C 6-alcohols during the postharvest process of oolong tea. Food Sci. Nutr. 2019, 8, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, T.; Ishii, R.; Onishi, H.; Ota, S. Elucidation of biochemical pathways underlying VOCs production in A549 cells. Front. Mol. Biosci. 2020, 7, 116. [Google Scholar] [CrossRef]
- Domínguez, T.; Hernández, M.L.; Pennycooke, J.C.; Jiménez, P.; Martínez-Rivas, J.M.; Sanz, C.; Stockinger, E.J.; Sánchez-Serrano, J.J.; Sanmartín, M. Increasing ω-3 desaturase expression in tomato results in altered aroma profile and enhanced resistance to cold stress. Plant Physiol. 2010, 153, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Speirs, J.; Lee, E.; Holt, K.; Yong-Duk, K.; Scott, N.S.; Loveys, B.; Schuch, W. Genetic manipulation of alcohol dehydrogenase levels in ripening tomato fruit affects the balance of some flavor aldehydes and alcohols. Plant Physiol. 1998, 117, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moummou, H.; Kallberg, Y.; Tonfack, L.B.; Persson, B.; Van Der Rest, B. The Plant short-chain dehydrogenase (SDR) superfamily: Genome-wide inventory and diversification patterns. BMC Plant Biol. 2012, 12, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosset, V.; Harmel, N.; Göbel, C.; Francis, F.; Haubruge, E.; Wathelet, J.-P.; Du Jardin, P.; Feussner, I.; Fauconnier, M.-L. Attacks by a piercing-sucking insect (Myzus persicae Sultzer) or a chewing insect (Leptinotarsa decemlineata Say) on potato plants (Solanum tuberosum L.) induce differential changes in volatile compound release and oxylipin synthesis. J. Exp. Bot. 2009, 60, 1231–1240. [Google Scholar] [CrossRef] [Green Version]
- Campestre, C.; Angelini, G.; Gasbarri, C.; Angerosa, F. The compounds responsible for the sensory profile in monovarietal virgin olive oils. Molecules 2017, 22, 1833. [Google Scholar] [CrossRef] [PubMed]
- Vong, W.C.; Liu, S.-Q. Bioconversion of green volatiles in okara (Soybean residue) into esters by coupling enzyme catalysis and yeast (Lindnera saturnus) fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 10017–10026. [Google Scholar] [CrossRef]
- Mariutto, M.; Duby, F.; Adam, A.; Bureau, C.; Fauconnier, M.-L.; Ongena, M.; Thonart, P.; Dommes, J. The elicitation of a systemic resistance by Pseudomonas putida BTP1 in tomato involves the stimulation of two lipoxygenase isoforms. BMC Plant Biol. 2011, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Longhurst, T.J.; Tung, H.F.; Brady, C.J. Developmental Regulation of the Expression of Alcohol Dehydrogenase in Ripening Tomato Fruits. J. Food Biochem. 1990, 14, 421–433. [Google Scholar] [CrossRef]
- Van Der Straeten, D.; Rodrigues-Pousada, R.A.; Gielen, J.; Van Montagu, M. Tomato alcohol dehydrogenase: Expression during fruit ripening and under hypoxic conditions. FEBS Lett. 1991, 295, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Buttery, R.G.; Teranishi, R.; Flath, R.A.; Ling, L.C. Identification of additional tomato paste volatiles. J. Agric. Food Chem. 1990, 38, 792–795. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. Branched-chain amino acids: Enzyme and substrate regulation. J. Nutr. 2006, 136, 207S–211S. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.; Binder, S. The mitochondrial branched-chain aminotransferase (AtBCAT-1) is capable to initiate degradation of leucine, isoleucine and valine in almost all tissues in Arabidopsis thaliana. Plant Mol. Biol. 2005, 57, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Maloney, G.S.; Kochevenko, A.; Tieman, D.M.; Tohge, T.; Krieger, U.; Zamir, D.; Taylor, M.G.; Fernie, A.R.; Klee, H.J. Characterization of the branched-chain amino acid aminotransferase enzyme family in tomato. Plant Physiol. 2010, 153, 925–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochevenko, A.; Klee, H.J.; Fernie, A.R.; Araújo, W.L. Molecular identification of a further branched-chain aminotransferase 7 (BCAT7) in tomato plants. J. Plant Physiol. 2012, 169, 437–443. [Google Scholar] [CrossRef]
- Tohge, T.; Alseekh, S.; Fernie, A.R. On the regulation and function of secondary metabolism during fruit development and ripening. J. Exp. Bot. 2013, 65, 4599–4611. [Google Scholar] [CrossRef] [Green Version]
- Marilley, L. Flavours of cheese products: Metabolic pathways, analytical tools and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef]
- Mooney, B.P.; Miernyk, J.A.; Randall, D.D. Thecomplexfate ofα-ketoacids. Annu. Rev. Plant Biol. 2002, 53, 357–375. [Google Scholar] [CrossRef]
- Simkin, A.J.; Schwartz, S.H.; Auldridge, M.; Taylor, M.G.; Klee, H.J. The tomato carotenoid cleavage dioxygenase 1 genes contribute to the formation of the flavor volatiles β-ionone, pseudoionone, and geranylacetone. Plant J. 2004, 40, 882–892. [Google Scholar] [CrossRef]
- Wei, Y.; Wan, H.; Wu, Z.; Wang, R.; Ruan, M.; Ye, Q.; Li, Z.; Zhou, G.; Yao, Z.; Yang, Y.; et al. A comprehensive analysis of carotenoid cleavage dioxygenases genes in Solanum lycopersicum. Plant Mol. Biol. Rep. 2015, 34, 512–523. [Google Scholar] [CrossRef]
- Floss, D.S.; Walter, M.H. Role of carotenoid cleavage dioxygenase 1 (CCD1) in apocarotenoid biogenesis revisited. Plant Signal. Behav. 2009, 4, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Vogel, J.T.; Tan, B.-C.; Mccarty, D.R.; Klee, H.J. The carotenoid cleavage dioxygenase 1 enzyme has broad substrate specificity, cleaving multiple carotenoids at two different bond positions. J. Biol. Chem. 2008, 283, 11364–11373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lashbrooke, J.G.; Young, P.R.; Dockrall, S.J.; Vasanth, K.; A Vivier, M. Functional characterisation of three members of the Vitis vinifera L. carotenoid cleavage dioxygenase gene family. BMC Plant Biol. 2013, 13, 156. [Google Scholar] [CrossRef] [Green Version]
- Frusciante, S.; Diretto, G.; Bruno, M.; Ferrante, P.; Pietrella, M.; Prado-Cabrero, A.; Rubio, A.-M.; Beyer, P.D.; Gomez-Gomez, L.; Al-Babili, S.; et al. Novel carotenoid cleavage dioxygenase catalyzes the first dedicated step in saffron crocin biosynthesis. Proc. Natl. Acad. Sci. USA 2014, 111, 12246–12251. [Google Scholar] [CrossRef] [Green Version]
- Ilg, A.; Bruno, M.; Beyer, P.; Al-Babili, S. Tomato carotenoid cleavage dioxygenases 1A and 1B: Relaxed double bond specificity leads to a plenitude of dialdehydes, mono-apocarotenoids and isoprenoid volatiles. FEBS Open Bio 2014, 4, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Fischer, K.; Yu, S.; Dubcovsky, J.; Tian, L. Distinct expression and function of carotenoid metabolic genes and homoeologs in developing wheat grains. BMC Plant Biol. 2016, 16, 155. [Google Scholar] [CrossRef] [Green Version]
- Klee, H.J. Improving the flavor of fresh fruits: Genomics, biochemistry, and biotechnology. New Phytol. 2010, 187, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.; Quiroz, L.F.; Rodriguez-Concepción, M.; Stange, C. Differential contribution of the first two enzymes of the MEP pathway to the supply of metabolic precursors for carotenoid and chlorophyll biosynthesis in carrot (Daucus carota). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoharan, R.K.; Jung, H.-J.; Hwang, I.; Jeong, N.; Kho, K.H.; Chung, M.-Y.; Nou, I.S. Molecular breeding of a novel orange-brown tomato fruit with enhanced β-carotene and chlorophyll accumulation. Hereditas 2017, 154, 1. [Google Scholar] [CrossRef] [Green Version]
- Enfissi, E.M.; Nogueira, M.; Bramley, P.M.; Fraser, P. The regulation of carotenoid formation in tomato fruit. Plant J. 2017, 89, 774–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Shao, Z.; Zhang, M.; Wang, Q. Regulation of carotenoid metabolism in tomato. Mol. Plant 2015, 8, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Alcázar, M.; Giménez, E.; Pineda, B.; Capel, C.; García-Sogo, B.; Sánchez, S.; Yuste-Lisbona, F.J.; Angosto, T.; Capel, J.; Moreno, V.; et al. Albino T-DNA tomato mutant reveals a key function of 1-deoxy-D-xylulose-5-phosphate synthase (DXS1) in Plant development and survival. Sci. Rep. 2017, 7, 45333. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, L.; Simon-Moya, M.; Llorente, B.; Llamas, E.; Marro, M.; Loza-Alvarez, P.; Li, L.; Rodríguez-Concepción, M. Interference with Clp protease impairs carotenoid accumulation during tomato fruit ripening. J. Exp. Bot. 2018, 69, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- McQuinn, R.P.; Wong, B.; Giovannoni, J.J. AtPDSoverexpression in tomato: Exposing unique patterns of carotenoid self-regulation and an alternative strategy for the enhancement of fruit carotenoid content. Plant Biotechnol. J. 2018, 16, 482–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chetelat, R.T.; Qin, X.; Tan, M.; Burkart-Waco, D.; Moritama, Y.; Huo, X.; Wills, T.; Pertuzé, R. Introgression lines of Solanum sitiens a wild nightshade of the Atacama Desert, in the genome of cultivated tomato. Plant J. 2019, 100, 836–850. [Google Scholar] [CrossRef]
- Xiong, C.; Luo, D.; Lin, A.; Zhang, C.; Shan, L.; He, P.; Li, B.; Zhang, Q.; Hua, B.; Yuan, Z.; et al. A tomato B-box proteinSlBBX20 modulates carotenoid biosynthesis by directly activatingPHYTOENE SYNTHASE 1, and is targeted for 26S proteasome-mediated degradation. New Phytol. 2019, 221, 279–294. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.; Pfander, H.; Liaaen-Jensen, S. Carotenoids; Birkhäuser Verlag: Basel, Switzerland, 1998; Volume 3. [Google Scholar]
- Galpaz, N.; Wang, Q.; Menda, N.; Zamir, D.; Hirschberg, J. Abscisic acid deficiency in the tomato mutant high-pigment 3 leading to increased plastid number and higher fruit lycopene content. Plant J. 2008, 53, 717–730. [Google Scholar] [CrossRef]
- Mann, V.; Pecker, I.; Hirschberg, J. Cloning and characterization of the gene for phytoene desaturase (Pds) from tomato (Lycopersicon esculentum). Plant Mol. Biol. 1994, 24, 429–434. [Google Scholar] [CrossRef]
- Pecker, I.; Gabbay, R.; Cunningham, F.X.; Hirschberg, J. Cloning and characterization of the cDNA for lycopene β-cyclase from tomato reveals decrease in its expression during fruit ripening. Plant Mol. Biol. 1996, 30, 807–819. [Google Scholar] [CrossRef]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to β-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lois, L.M.; Rodriguez-Concepcion, M.; Gallego, F.; Campos, N.; Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J. 2000, 22, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Scolnik, P.A. cDNA cloning, expression during development, and genome mapping of PSY2, a second tomato gene encoding phytoene synthase. J. Biol. Chem. 1993, 268, 25718–25721. [Google Scholar] [CrossRef]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development (Evidence for tissue-specific gene expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Enfissi, E.M.; Halket, J.M.; Truesdale, M.R.; Yu, D.; Gerrish, C.; Bramley, P.M. Manipulation of phytoene levels in tomato fruit: Effects on isoprenoids, plastids, and intermediary metabolism. Plant Cell 2007, 19, 3194–3211. [Google Scholar] [CrossRef] [Green Version]
- Giorio, G.; Stigliani, A.L.; D’Ambrosio, C. Phytoene synthase genes in tomato (Solanumlycopersicum, L.) -new data on the structures, the deduced amino acid sequences and the expression patterns. FEBS J. 2007, 275, 527–535. [Google Scholar] [CrossRef]
- Rodríguez-Villalón, A.; Gas, E.; Rodríguez-Concepción, M. Phytoene synthase activity controls the biosynthesis of carotenoids and the supply of their metabolic precursors in dark-grown Arabidopsis seedlings. Plant J. 2009, 60, 424–435. [Google Scholar] [CrossRef]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Ronen, G.; Cohen, M.; Zamir, D.; Hirschberg, J. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant delta. Plant J. 1999, 17, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Apel, W.; Bock, R. Enhancement of carotenoid biosynthesis in transplastomic tomatoes by induced Lycopene-to-Provitamin a conversion. Plant Physiol. 2009, 151, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ambrosio, C.; Stigliani, A.L.; Giorio, G. Overexpression of CrtR-b2 (carotene β hydroxylase 2) from S. lycopersicum L. differentially affects xanthophyll synthesis and accumulation in transgenic tomato plants. Transgenic Res. 2010, 20, 47–60. [Google Scholar] [CrossRef]
- Gonzalez-Jorge, S.; Mehrshahi, P.; Magallanes-Lundback, M.; Lipka, A.E.; Angelovici, R.; Gore, M.A.; DellaPenna, D. zeaxanthin epoxidase activity potentiates carotenoid degradation in maturing seed. Plant Physiol. 2016, 171, 1837–1851. [Google Scholar] [CrossRef] [Green Version]
- Alder, A.; Jamil, M.; Marzorati, M.; Bruno, M.; Vermathen, M.; Bigler, P.; Ghisla, S.; Bouwmeester, H.; Beyer, P.; Al-Babili, S.; et al. The path from-carotene to carlactone, a strigolactone-like plant hormone. Science 2012, 335, 1348–1351. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.-C.; Molnár, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.; Ducreux, L.J.; Morris, W.L.; Morris, J.A.; Suttle, J.C.; Ramsay, G.; Bryan, G.J.; Hedley, P.E.; Taylor, M.A. The metabolic and developmental roles of carotenoid cleavage dioxygenase4 from potato. Plant Physiol. 2010, 154, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Brandi, F.; Bar, E.; Mourgues, F.; Horváth, G.; Turcsi, E.; Giuliano, G.; Liverani, A.; Tartarini, S.; Lewinsohn, E.; Rosati, C. Study of “Redhaven” peach and its white-fleshed mutant suggests a key role of CCD4 carotenoid dioxygenase in carotenoid and norisoprenoid volatile metabolism. BMC Plant Biol. 2011, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.-J.; Jeong, H.-B.; Jung, A.; Kang, M.-Y.; Kim, S.; Ha, S.-H.; Kwon, J.-K.; Kang, B.-C. Phytoene synthase 2 can compensate for the absence of PSY1 in the control of color in Capsicum fruit. J. Exp. Bot. 2020, 71, 3417–3427. [Google Scholar] [CrossRef]
- Fantini, E.; Falcone, G.; Frusciante, S.; Giliberto, L.; Giuliano, G. Dissection of tomato Lycopene biosynthesis through virus-induced gene silencing. Plant Physiol. 2013, 163, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Hirata, H.; Sayama, H.; Sekiguchi, K.; Itano, H.; Asai, T.; Dohra, H.; Hara, M.; Watanabe, N. Production of 2-Phenylethanol in roses as the dominant floral scent compound fromL-phenylalanine by two key enzymes, a PLP-dependent decarboxylase and a phenylacetaldehyde reductase. Biosci. Biotechnol. Biochem. 2007, 71, 2408–2419. [Google Scholar] [CrossRef]
- Xie, Q.; Liu, Z.; Meir, S.; Rogachev, I.; Aharoni, A.; Klee, H.J.; Galili, G. Altered metabolite accumulation in tomato fruits by coexpressing a feedback-insensitive AroG and the Ph ODO 1 MYB-type transcription factor. Plant Biotechnol. J. 2016, 14, 2300–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieman, D.M.; Loucas, H.M.; Kim, J.Y.; Clark, D.G.; Klee, H.J. Tomato phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma volatile 2-phenylethanol. Phytochemistry 2007, 68, 2660–2669. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.A.; Marques, J.V.; Dalisay, D.S.; Herman, B.; Bedgar, D.L.; Davin, L.B.; Lewis, N.G. Transgenic hybrid poplar for sustainable and scalable production of the commodity/specialty chemical, 2-Phenylethanol. PLoS ONE 2013, 8, e83169. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.-A.; Deng, X. Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of Penicillium molds of citrus fruits. BMC Microbiol. 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Rastogi, S.; Dwivedi, U.N. Phenylpropanoid metabolism in ripening fruits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 398–416. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [Green Version]
- Biała, W.; Jasiński, M. The phenylpropanoid case-it is transport that matters. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Koeduka, T.; Fridman, E.; Gang, D.R.; Vassão, D.G.; Jackson, B.L.; Kish, C.M.; Orlova, I.; Spassova, S.M.; Lewis, N.G.; Noel, J.P.; et al. Eugenol and isoeugenol, characteristic aromatic constituents of spices, are biosynthesized via reduction of a coniferyl alcohol ester. Proc. Natl. Acad. Sci. USA 2006, 103, 10128–10133. [Google Scholar] [CrossRef] [Green Version]
- Tieman, D.; Zeigler, M.; Schmelz, E.; Taylor, M.G.; Rushing, S.; Jones, J.B.; Klee, H.J. Functional analysis of a tomato salicylic acid methyl transferase and its role in synthesis of the flavor volatile methyl salicylate. Plant J. 2010, 62, 113–123. [Google Scholar] [CrossRef]
- Mageroy, M.H.; Tieman, D.M.; Floystad, A.; Taylor, M.G.; Klee, H.J. A Solanum lycopersicum catechol-O-methyltransferase involved in synthesis of the flavor molecule guaiacol. Plant J. 2012, 69, 1043–1051. [Google Scholar] [CrossRef]
- Tikunov, Y.; Molthoff, J.; De Vos, R.C.; Beekwilder, J.; Van Houwelingen, A.; Van Der Hooft, J.J.; Vries, M.N.-D.; Labrie, C.W.; Verkerke, W.; Van De Geest, H.; et al. Non-smoky glycosyltransferase1 prevents the release of smoky aroma from tomato fruit. Plant Cell 2013, 25, 3067–3078. [Google Scholar] [CrossRef] [Green Version]
- Klee, H.J.; Tieman, D.M. The genetics of fruit flavour preferences. Nat. Rev. Genet. 2018, 19, 347–356. [Google Scholar] [CrossRef]
- Rigano, M.M.; Raiola, A.; Docimo, T.; Ruggieri, V.; Calafiore, R.; Vitaglione, P.; Ferracane, R.; Frusciante, L.; Barone, A. Metabolic and molecular changes of the phenylpropanoid pathway in tomato (Solanum lycopersicum) lines carrying different Solanum pennellii wild chromosomal regions. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.Y.; Kim, H.M.; Ma, S.H.; Park, S.Y.; Dat, M.T.; Jang, G.; Joung, Y.H. The promoter of tomato Histidine decarboxylase A is fruit-specific, and its expression is stably maintained in fruits during ripening. Plant Biotechnol. Rep. 2019, 13, 43–50. [Google Scholar] [CrossRef]
- Kumar, R. Evolutionary trails of plant group ii pyridoxal phosphate-dependent decarboxylase genes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellenreuther, M.; Mérot, C.; Berdan, E.; Bernatchez, L. Going beyond SNPs: The role of structural genomic variants in adaptive evolution and species diversification. Mol. Ecol. 2019, 28, 1203–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Cao, K.; Deng, C.; Li, Y.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wu, J.; Guan, L.; et al. An integrated peach genome structural variation map uncovers genes associated with fruit traits. Genome Biol. 2020, 21, 258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mao, L.; Chen, H.; Bu, F.; Li, G.; Sun, J.; Li, S.; Sun, H.; Jiao, C.; Blakely, R.; et al. Genome-wide mapping of structural variations reveals a copy number variant that determines reproductive morphology in cucumber. Plant Cell 2015, 27, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Liu, Z.-Q.; Mocoeur, A.; Xia, Y.; Jing, H.-C. PAV markers in Sorghum bicolour: Genome pattern, affected genes and pathways, and genetic linkage map construction. Theor. Appl. Genet. 2015, 128, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHale, L.K.; Haun, W.J.; Xu, W.W.; Bhaskar, P.B.; Anderson, J.E.; Hyten, D.L.; Gerhardt, D.J.; Jeddeloh, J.A.; Stupar, R.M. Structural variants in the soybean genome localize to clusters of biotic stress-response genes. Plant Physiol. 2012, 159, 1295–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabur, I.; Chawla, H.S.; Lopisso, D.T.; Von Tiedemann, A.; Snowdon, R.J.; Obermeier, C. Gene presence-absence variation associates with quantitative Verticillium longisporum disease resistance in Brassica napus. Sci. Rep. 2020, 10, 4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgotra, R.K.; Stewart, C.N. Functional markers for precision Plant breeding. Int. J. Mol. Sci. 2020, 21, 4792. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-B.; Yang, M.-H.; Zeng, F.; Biligetu, B. Searching for an accurate marker-based prediction of an individual quantitative trait in molecular plant breeding. Front. Plant Sci. 2017, 8, 1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compound | Odor Descriptors * |
|---|---|---|
| Apocarotenoid | 6-Methyl-5-hepten-2-one | Citrus, green, musty, lemongrass, apple |
| Apocarotenoid | Geranial | Sharp, lemon, sweet |
| Apocarotenoid | β-Damascenone | Apple, rose, honey, tobacco, sweet |
| Apocarotenoid | Geranylacetone | Fresh, green, fruity, waxy, rose, woody, magnolia, tropical |
| Apocarotenoid | β-Ionone | Floral, woody, sweet, fruity, berry, tropical, beeswax |
| BCAA | 3-Methylbutanal | Ethereal, aldehydic, chocolate, peach, fatty |
| BCAA | 2-Methylbutanal | Musty, cocoa, coffee, nutty |
| BCAA | 3-Methylbutanol | Fusel, oil, alcoholic, whiskey, fruity, banana |
| BCAA | 2-Isobuthylthiazole | Green, wasabi, privet, tomato, leaf, earthy, vegetable, metallic |
| Lipid | 1-Penten-3-one | Pungent, peppery, mustard, garlic, onion |
| Lipid | (Z)-3-Hexenal | Green, fatty, grassy, weedy, fruity, apple |
| Lipid | Hexenal | Sweet, almond, fruity, green, leafy, apple, plum, vegetable |
| Lipid | (E)-2-Hexenal | Sharp, fresh, leafy, green, clean, fruity, spicy, herbal |
| Lipid | (E)-2-Heptenal | Pungent, green, vegetable, fresh, fatty |
| Lipid | (E,E)-2.4-Decadienal | Oily, cucumber, melon, citrus, pumpkin, nut, meat |
| Phenolic | Phenylacetaldehyde | Green, sweet, floral, hyacinth, clover, honey, cocoa |
| Phenolic | 2-Phenylethanol | Sweet, floral, fresh, bready, rose, honey |
| Phenolic | 1-Nitro-2-phenylethane | Flower, spice |
| Phenylpropanoid | Guaiacol | Phenolic, smoke, spice, vanilla, woody |
| Phenylpropanoid | Methyl salicylate | Wintergreen, mint |
| Phenylpropanoid | Eugenol | Sweet, spicy, clove, woody |
| Author, Year | Population | References |
|---|---|---|
| Saliba-Colombani et al., 2001 | RILs | [90] |
| Fulton et al., 2002 | BCs | [91] |
| Tadmor et al., 2002 | ILs | [92] |
| Tieman et al., 2006 | ILs | [93] |
| Mathieu et al., 2009 | ILs | [6] |
| Kochevenko & Fernie, 2011 | ILs | [94] |
| Ruggieri et al., 2014 | GWAS | [95] |
| Sauvage et al., 2014 | GWAS | [74] |
| Capel et al., 2015 | RILs | [96] |
| Zhao et al., 2016 | GWAS | [97] |
| Baldina et al., 2016 | GWAS | [98] |
| Bauchet et al., 2017 | GWAS | [99] |
| Tieman et al., 2017 | GWAS | [25] |
| Garbowicz et al., 2018 | ILs | [100] |
| Kimbara et al., 2018 | RILs | [101] |
| Tikunov et al., 2020 | F2, F6, DP | [102] |
| Solyc ID | Name | QGR | Start (bp) | Annotation | Reference | Expressed (Y/N) |
|---|---|---|---|---|---|---|
| Solyc01g006540 | loxC | LIP1 | 1,113,718 | Lipoxygenase | [68] | Y |
| Solyc01g006560 | loxF | LIP1 | 1,128,815 | Lipoxygenase | [121] | N |
| Solyc01g099210 | - | LIP1 | 89,509,072 | Lipoxygenase | Y | |
| Solyc01g108150 | - | LIP2 | 95,505,678 | Oxidoreductase zinc-binding dehydrogenase family protein | Y | |
| Solyc02g090930 | - | LIP2 | 52,398,542 | Lipase | Y | |
| Solyc03g093360 | - | LIP3 | 54,632,041 | Lipoxygenase | Y | |
| Solyc03g095360 | - | LIP5 | 56,412,064 | Alcohol dehydrogenase zinc-binding | Y | |
| Solyc03g111550 | - | LIP5 | 62,149,409 | GDSL esterase/lipase | Y | |
| Solyc04g010250 | - | LIP6 | 3,577,656 | Lipase-like protein | Y | |
| Solyc04g054980 | - | LIP7 | 53,517,056 | Lipoxygenase homology domain-containing protein 1 | Y | |
| Solyc04g054990 | - | LIP7 | 53,522,447 | Lipoxygenase homology domain-containing protein 1 | Y | |
| Solyc04g064710 | - | LIP7 | 55,858,388 | Alcohol dehydrogenase 2 | Y | |
| Solyc05g005480 | EO QR | LIP9 | 352,211 | Oxidoreductase zinc-binding dehydrogenase | Y | |
| Solyc05g009390 | - | LIP9 | 3,544,526 | Lipase-like protein | Y | |
| Solyc06g059740 | ADH2 | LIP11 | 37,606,747 | Alcohol dehydrogenase 2 | Y | |
| Solyc07g045090 | - | LIP13 | 58,206,449 | Alcohol dehydrogenase zinc-binding domain protein | [116] | Y |
| Solyc08g014000 | LOXA | LIP14 | 3,516,113 | Lipoxygenase | Y | |
| Solyc09g059030 | - | LIP17 | 53,010,842 | Alcohol dehydrogenase zinc-containing | Y | |
| Solyc09g059040 | - | LIP17 | 53,050,984 | Alcohol dehydrogenase zinc-containing | Y | |
| Solyc09g091050 | - | LIP18 | 70,389,731 | Lipase | Y | |
| Solyc11g010960 | - | LIP20 | 4,006,687 | Alcohol dehydrogenase | Y | |
| Solyc11g011330 | - | LIP20 | 4,375,432 | Cinnamyl alcohol dehydrogenase | Y | |
| Solyc11g065530 | - | LIP21 | 50,971,492 | Lipase (Fragment) | Y | |
| Solyc11g071290 | - | LIP21 | 54,819,572 | Alcohol dehydrogenase | Y | |
| Solyc12g010950 | - | LIP22 | 3,827,848 | Alcohol dehydrogenase zinc-containing | Y | |
| Solyc12g011040 | - | LIP22 | 3,894,074 | Lipoxygenase | Y | |
| Solyc12g096760 | - | LIP24 | 65,539,079 | Alcohol dehydrogenase zinc-containing | Y | |
| Solyc12g096780 | - | LIP24 | 65,557,520 | Mitochondrial trans-2-enoyl-CoA reductase | Y | |
| Solyc ID | Name | QGR | Start (bp) | Annotation | Family | References | Expressed (Y/N) |
|---|---|---|---|---|---|---|---|
| Solyc01g098700 | SlBCAT7 | 89,114,957 | Branched-chain-amino-acid aminotransferase 7 | BCAT | [128] | Y | |
| Solyc02g091970 | SlBCAT3 | BCAA4 | 53,178,240 | Branched-chain-amino-acid aminotransferase 3 | BCAT | [127] | Y |
| Solyc03g005730 | BCAA5 | 509,260 | 3-isopropylmalate dehydratase large subunit 2 | IPMD | Y | ||
| Solyc03g007200 | BCAA5 | 1,775,153 | Oxidoreductase zinc-containing alcohol dehydrogenase family | ADH | Y | ||
| Solyc03g043880 | SlBCAT4 | 7,542,329 | Branched-chain amino acid aminotransferase 4 | BCAT | [127] | N | |
| Solyc03g097680 | BCAA6 | 60,009,332 | Pyruvate dehydrogenase E1 component subunit β | PDH | Y | ||
| Solyc04g008590 | BCAA7 | 2,195,990 | Pyruvate dehydrogenase E1 component subunit β | PDH | Y | ||
| Solyc04g063350 | FLORAL4 | BCAA9 | 55,462,543 | 3-methyl-2-oxobutanoate dehydrogenase | [102] | Y | |
| Solyc04g064710 | BCAA9 | 55,858,388 | Alcohol dehydrogenase 2 | ADH | Y | ||
| Solyc05g005480 | EO QR | BCAA10 | 352,211 | Oxidoreductase zinc-binding dehydrogenase | ADH | Y | |
| Solyc06g059880 | BCAA11 | 37,781,458 | Acetolactate synthase | ALS | Y | ||
| Solyc06g060790 | BCAA11 | 38,814,018 | 3-isopropylmalate dehydratase small subunit | IPMD | [94] | N | |
| Solyc07g021630 | SlBCAT2 | 20,381,557 | Branched-chain amino acid aminotransferase 2 | BCAT | [127] | Y | |
| Solyc07g045090 | BCAA13 | 58,206,449 | Alcohol dehydrogenase zinc-binding domain protein | ADH | Y | ||
| Solyc07g053280 | BCAA14 | 61,749,215 | Ketol-acid reductoisomerase | KARI | Y | ||
| Solyc07g061940 | BCAA14 | 64,814,098 | Acetolactate synthase | ALS | Y | ||
| Solyc08g014130 | BCAA15 | 3,972,785 | 2-isopropylmalate synthase 1 | IPMS | Y | ||
| Solyc09g008670 | TD | BCAA17 | 2,123,840 | Threonine ammonia-lyase biosynthetic | TD | Y | |
| Solyc11g010960 | BCAA21 | 4,006,687 | Alcohol dehydrogenase | ADH | Y | ||
| Solyc11g011330 | BCAA21 | 4,375,432 | Cinnamyl alcohol dehydrogenase | ADH | Y | ||
| Solyc11g071280 | LOC778238 | BCAA24 | 54,809,918 | Branched-chain amino acid aminotransferase like protein | BCAT | Y | |
| Solyc11g071290 | BCAA24 | 54,819,572 | Alcohol dehydrogenase | ADH | Y | ||
| Solyc12g005860 | BCAA25 | 490,745 | 3-isopropylmalate dehydratase large subunit | IPMD | Y | ||
| Solyc12g009400 | BCAA25 | 2,682,210 | Pyruvate dehydrogenase E1 component α subunit | PDH | Y | ||
| Solyc12g009410 | BCAA25 | 2,687,666 | Pyruvate dehydrogenase E1 component α subunit | PDH | Y | ||
| Solyc12g010840 | BCAA25 | 3,773,065 | Ketol-acid reductoisomerase | KARI | Y | ||
| Solyc12g010950 | BCAA25 | 3,827,848 | Alcohol dehydrogenase zinc-containing | ADH | Y | ||
| Solyc12g088220 | SlBCAT1 | BCAA26 | 63,663,328 | Branched-chain-amino-acid aminotransferase 1 | BCAT | [127] | Y |
| Solyc12g096760 | BCAA26 | 65,539,079 | Alcohol dehydrogenase zinc-containing | ADH | Y | ||
| Solyc12g096780 | BCAA26 | 65,557,520 | Mitochondrial trans-2-enoyl-CoA reductase | ADH | Y |
| Solyc ID | Name | QGR | Start (bp) | Annotation | References | Expressed (Y/N) |
|---|---|---|---|---|---|---|
| Solyc01g005940 | PSY3 | APO1 | 613,955 | Phytoene synthase 3 | [171] | N |
| Solyc01g067890 | DXS1 | APO2 | 76,868,469 | 1-Deoxy-d-xylulose 5-phosphate synthase 1 | [145,155] | N |
| Solyc01g087250 | CCD1A | APO2 | 82,184,585 | Carotenoid cleavage dioxygenase 1A | [132,138] | N |
| Solyc01g087260 | CCD1B | APO2 | 82,196,996 | Carotenoid cleavage dioxygenase 1B | [132,138] | N |
| Solyc01g087270 | CCD2 | APO2 | 82,209,237 | Carotenoid cleavage dioxygenase 2 | [137] | N |
| Solyc01g090660 | CCD7 | APO2 | 84,307,951 | Carotenoid cleavage dioxygenase 7 | [166] | N |
| Solyc02g081330 | PSY2 | APO3/4 | 45,335,358 | Phytoene synthase 2 | N | |
| Solyc02g090890 | ZEP | APO4 | 52,369,708 | Zeaxanthin epoxidase, 2C chloroplastic | [151] | N |
| Solyc03g007960 | CrtZ-2 | APO5 | 2,447,872 | β-Carotene hydroxylase 2 | [164] | Y |
| Solyc03g031860 | PSY1 | APO5 | 4,326,134 | Phytoene synthase 1 | [171] | Y |
| Solyc03g114340 | DXR | 64,347,674 | 1-Deoxy-d-xylulose-5-phosphate reductoisomerase | [143] | N | |
| Solyc03g123760 | PDS | 70,501,011 | Phytoene desaturase | [152] | Y | |
| Solyc04g040190 | LCY-B1 | 11,947,053 | Lycopene β-cyclase 1 | [153] | N | |
| Solyc06g036260 | CrtZ-1 | 25,742,578 | β-Carotene hydroxylase 1 | [162] | Y | |
| Solyc06g074240 | LCY-B2 | APO12 | 45,898,227 | Lycopene β-cyclase 2 | [153] | N |
| Solyc08g075480 | CCD4A | 59,627,153 | Carotenoid cleavage dioxygenase 4A | [167] | N | |
| Solyc08g075490 | CCD4B | 59,643,898 | Carotenoid cleavage dioxygenase 4B | [167] | N | |
| Solyc10g079480 | LCY-b | APO17 | 61,024,821 | Lycopene β-cyclase | [153] | N |
| Solyc10g081650 | CRTISO | APO17 | 62,682,440 | Carotenoid isomerase, 2C chloroplastic | [161] | Y |
| Solyc11g010850 | DXS2 | APO18 | 3,870,455 | 1-Deoxy-d-xylulose 5-phosphate synthase 2 | [145,155] | N |
| Solyc ID | Name | QGR | Start (bp) | Annotation | References | Expressed (Y/N) |
|---|---|---|---|---|---|---|
| Solyc01g107910 | PHEN3 | 95,296,421 | Caffeoyl CoA 3-O-methyltransferase | Y | ||
| Solyc02g093270 | PHEN5 | 54,178,272 | Caffeoyl CoA O-methyltransferase | Y | ||
| Solyc03g097700 | PHEN6 | 60,029,697 | O-methyltransferase | Y | ||
| Solyc03g111830 | PHEN6 | 62,444,512 | O-methyltransferase | Y | ||
| Solyc04g063350 | FLORAL4 | PHEN7 | 55,462,543 | 3-methyl-2-oxobutanoate dehydrogenase | [102] | Y |
| Solyc04g071140 | PHEN7 | 58,070,304 | Decarboxylase family protein | Y | ||
| Solyc05g013440 | PHEN8 | 6,501,962 | Primary amine oxidase | Y | ||
| Solyc06g059840 | LOC778303 | PHEN9 | 37,729,682 | 3-methyl-2-oxobutanoate dehydrogenase | Y | |
| Solyc08g006740 | AADC2 | PHEN14 | 1,295,712 | Decarboxylase family protein | [175] | N |
| Solyc08g066240 | PHEN15 | 54,722,572 | Decarboxylase family protein | Y | ||
| Solyc08g066250 | Hdc | PHEN15 | 54,745,923 | Decarboxylase family protein | Y | |
| Solyc08g068600 | PHEN15 | 57,730,921 | Decarboxylase family protein | Y | ||
| Solyc08g068610 | AADC1B | PHEN15 | 57,740,004 | Decarboxylase family protein | [175] | Y |
| Solyc08g068680 | AADC1A | PHEN15 | 57,812,621 | Decarboxylase family protein | [175] | N |
| Solyc08g079430 | PHEN15 | 62,954,528 | Primary amine oxidase | Y | ||
| Solyc09g007890 | PHEN16 | 1,413,536 | Phenylalanine ammonia-lyase | Y | ||
| Solyc09g007900 | PHEN16 | 1,419,041 | Phenylalanine ammonia-lyase | Y | ||
| Solyc09g007910 | PHEN16 | 1,429,132 | Phenylalanine ammonia-lyase | Y | ||
| Solyc09g007920 | PHEN16 | 1,435,451 | Phenylalanine ammonia-lyase | Y | ||
| Solyc09g091550 | SAMT1 | PHEN17 | 70,802,564 | Salicylate methyltransferase 1 | [183] | N |
| NSGT1 | PHEN17 | 64,653,692 | Glycosyltransferase | [185] | Y | |
| Solyc10g005060 | CTOMT1 | 53,330 | Catechol-O-methyltransferase 1 | [184] | Y | |
| Solyc10g085830 | PHEN18 | 64,899,145 | O-methyltransferase 1 | Y | ||
| Solyc10g086180 | PHEN18 | 65,098,335 | Phenylalanine ammonia-lyase | Y | ||
| Solyc12g013690 | SlFMO1 | PHEN22 | 4,532,510 | Monooxygenase FAD-binding protein | [175] | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martina, M.; Tikunov, Y.; Portis, E.; Bovy, A.G. The Genetic Basis of Tomato Aroma. Genes 2021, 12, 226. https://doi.org/10.3390/genes12020226
Martina M, Tikunov Y, Portis E, Bovy AG. The Genetic Basis of Tomato Aroma. Genes. 2021; 12(2):226. https://doi.org/10.3390/genes12020226
Chicago/Turabian StyleMartina, Matteo, Yury Tikunov, Ezio Portis, and Arnaud G. Bovy. 2021. "The Genetic Basis of Tomato Aroma" Genes 12, no. 2: 226. https://doi.org/10.3390/genes12020226
APA StyleMartina, M., Tikunov, Y., Portis, E., & Bovy, A. G. (2021). The Genetic Basis of Tomato Aroma. Genes, 12(2), 226. https://doi.org/10.3390/genes12020226