
Discovery of a Major QTL Controlling Trichome IV Density in Tomato Using K-Seq Genotyping

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Glandular Type IV Trichome Density Characterization
2.3. DNA Extraction and Plant Genotyping by K-Seq
2.4. Genetic Mapping, QTL and QTL-Seq Analysis
3. Results
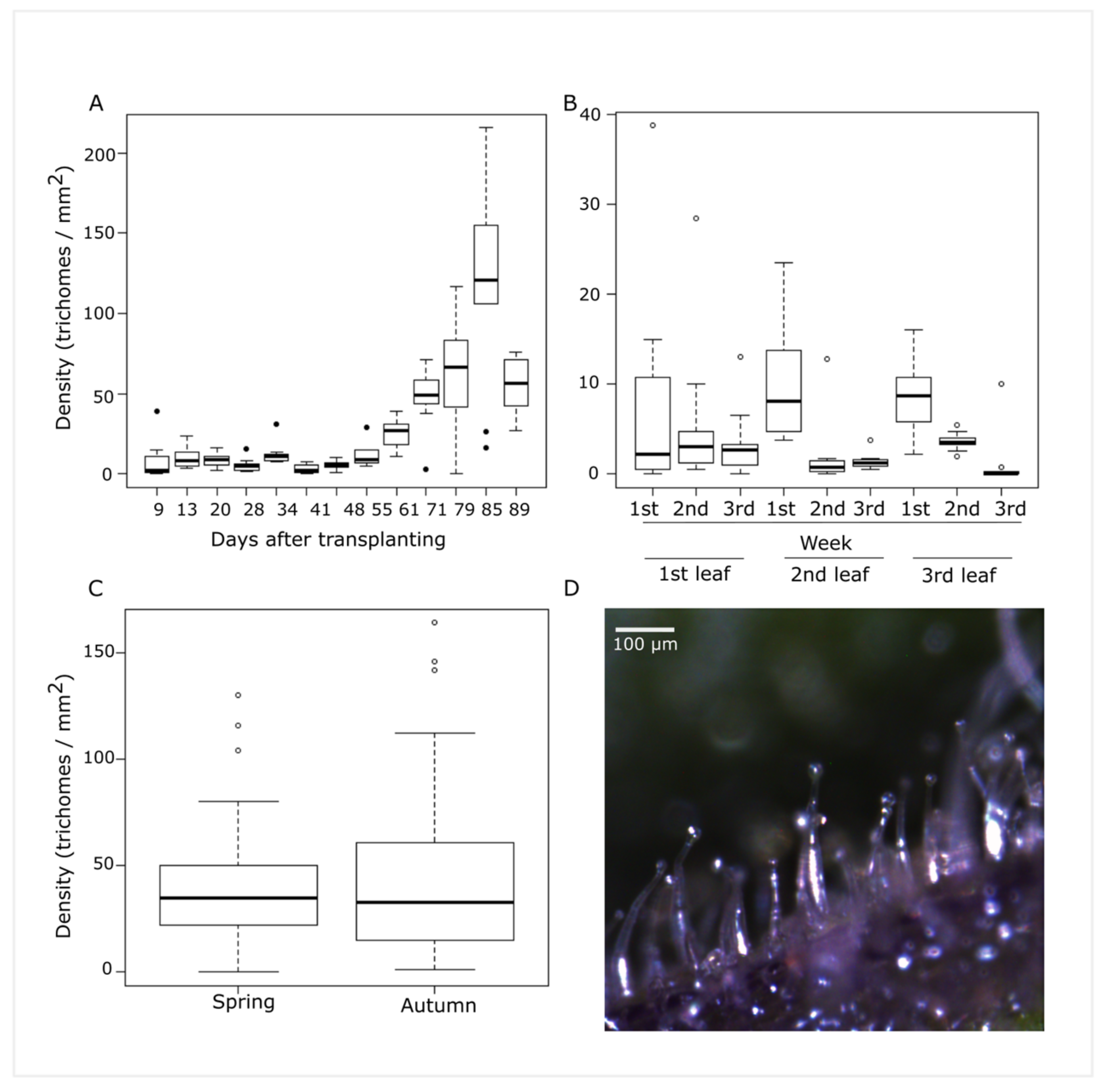
3.1. Type IV Trichome Density Characterization of S. pimpinellifolium Accession BGV016047 with a High Density of Type IV Trichomes
3.2. Trichome Type IV Density on Segregating Populations
3.3. QTL and QTLseq Analysis in BGV016047 × LA2278 Population
3.4. QTL Validation Using S. lycopersicum var. lycopersicum as Genetic Background
3.5. Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woodcock, B.A.; Garratt, M.P.D.; Powney, G.D.; Shaw, R.F.; Osborne, J.L.; Soroka, J.; Lindström, S.A.M.; Stanley, D.; Ouvrard, P.; Edwards, M.E.; et al. Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nat. Commun. 2019, 10, 1481. [Google Scholar] [CrossRef] [Green Version]
- Glas, J.J.; Schimmel, B.C.J.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [Green Version]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [Green Version]
- Peter, A.J.; Shanower, T.G.; Romeis, J. The role of plant trichomes in insect resistance: A selective review. Phytophaga 1995, 7, 41–63. [Google Scholar]
- Liedl, B.E.; Lawson, D.M.; White, K.K.; Shapiro, J.A.; Cohen, D.E.; Carson, W.G.; Trumble, J.T.; Mutschler, M.A. Acylsugars of Wild Tomato Lycopersicon pennellii Alters Settling and Reduces Oviposition of Bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 1995, 88, 742–748. [Google Scholar] [CrossRef]
- Peiffer, M.; Tooker, J.F.; Luthe, D.S.; Felton, G.W. Plants on early alert: Glandular trichomes as sensors for insect herbivores. New Phytol. 2009, 184, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Leong, B.J.; Lybrand, D.B.; Lou, Y.-R.; Fan, P.; Schilmiller, A.L.; Last, R.L. Evolution of metabolic novelty: A trichome-expressed invertase creates specialized metabolic diversity in wild tomato. Sci. Adv. 2019, 5, eaaw3754. [Google Scholar] [CrossRef] [Green Version]
- McDowell, E.T.; Kapteyn, J.; Schmidt, A.; Li, C.; Kang, J.-H.; Descour, A.; Shi, F.; Larson, M.; Schilmiller, A.; An, L.; et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol. 2011, 155, 524–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-S.; Kwon, M.; Cho, J.-H.; Im, J.-S.; Park, Y.-E.; Hong, S.-Y.; Hwang, I.-T.; Kang, J.-H. Characterization of trichome morphology and aphid resistance in cultivated and wild species of potato. Horticult. Environ. Biotechnol. 2017, 58, 450–457. [Google Scholar] [CrossRef]
- Luckwill, L.C. The Genus Lycopersicon: An Historical, Biological, and Taxonomic Survery of the Wild and Cultivated Tomatoes; Aberdeen University Press: Aberdeen, UK, 1943. [Google Scholar]
- Channarayappa, C.; Shivashankar, G.; Muniyappa, V.; Frist, R.H. Resistance of Lycopersicon species to Bemisia tabaci, a tomato leaf curl virus vector. Can. J. Bot. 1992, 70, 2184–2192. [Google Scholar] [CrossRef]
- Alba, J.M.; Montserrat, M.; Fernández-Muñoz, R. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Exp. Appl. Acarol. 2009, 47, 35–47. [Google Scholar] [CrossRef]
- Maluf, W.R.; Campos, G.A.; das Graças Cardoso, M. Relationships between trichome types and spider mite (Tetranychus evansi) repellence in tomatoes with respect to foliar zingiberene contents. Euphytica 2001, 121, 73–80. [Google Scholar] [CrossRef]
- Oriani, M.A.G.; Vendramim, J.D. Influence of trichomes on attractiveness and ovipositional preference of Bemisia tabaci (Genn.) B biotype (Hemiptera: Aleyrodidae) on tomato genotypes. Neotrop. Entomol. 2010, 39, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Firdaus, S. Identification of Whitefly Resistance in Tomato and Hot Pepper. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2012. [Google Scholar]
- Rakha, M.; Hanson, P.; Ramasamy, S. Identification of resistance to Bemisia tabaci Genn. in closely related wild relatives of cultivated tomato based on trichome type analysis and choice and no-choice assays. Genet. Resour. Crop Evol. 2017, 64, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.C.; da Silva, A.A.; Neiva, I.P.; Oliveira, I.R.C.; De Castro, E.M.; Francis, D.M.; Maluf, W.R. Inheritance of type IV glandular trichome density and its association with whitefly resistance from Solanum galapagense accession LA1401. Euphytica 2017, 213, 52. [Google Scholar] [CrossRef]
- Gurr, G.M.; McGrath, D. Foliar pubescence and resistance to potato moth, Phthorimaea operculella, in Lycopersicon hirsutum. Entomol. Exp. Appl. 2002, 103, 35–41. [Google Scholar] [CrossRef]
- Fan, P.; Leong, B.J.; Last, R.L. Tip of the trichome: Evolution of acylsugar metabolic diversity in Solanaceae. Curr. Opin. Plant Biol. 2019, 49, 8–16. [Google Scholar] [CrossRef]
- Guo, Z.; Weston, P.A.; Snyder, J.C. Repellency to two-spotted spider mite, Tetranychus urticae Koch, as related to leaf surface chemistry ofLycopersicon hirsutum accessions. J. Chem. Ecol. 1993, 19, 2965–2979. [Google Scholar] [CrossRef] [PubMed]
- Blauth, S.L.; Churchill, G.A.; Mutschler, M.A. Identification of quantitative trait loci associated with acylsugar accumulation using intraspecific populations of the wild tomato, Lycopersicon pennellii. Theor. Appl. Genet. 1998, 96, 458–467. [Google Scholar] [CrossRef]
- Lemke, C.A.; Mutschler, M.A. Inheritance of glandular trichomes in crosses between Lycopersicon esculentum and Lycopersicon pennellii. J. Am. Soc. Hortic. Sci. 1984, 109, 592–596. [Google Scholar]
- Momotaz, A.; Scott, J.W.; Schuster, D.J. Identification of quantitative trait loci conferring resistance to Bemisia tabaci in an F2 population of Solanum lycopersicum × Solanum habrochaites accession LA1777. J. Am. Soc. Hortic. Sci. 2010, 135, 134–142. [Google Scholar] [CrossRef]
- Firdaus, S.; van Heusden, A.W.; Hidayati, N.; Supena, E.D.J.; Mumm, R.; de Vos, R.C.H.; Visser, R.G.F.; Vosman, B. Identification and QTL mapping of whitefly resistance components in Solanum galapagense. Theor. Appl. Genet. 2013, 126, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, M.A.; Doerge, R.W.; Liu, S.C.; Kuai, J.P.; Liedl, B.E.; Shapiro, J.A. QTL analysis of pest resistance in the wild tomato Lycopersicon pennellii: QTLs controlling acylsugar level and composition. Theor. Appl. Genet. 1996, 92, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Leckie, B.M.; De Jong, D.M.; Mutschler, M.A. Quantitative trait loci increasing acylsugars in tomato breeding lines and their impacts on silverleaf whiteflies. Mol. Breed. 2012, 30, 1621–1634. [Google Scholar] [CrossRef]
- Fernández-Muñoz, R.; Salinas, M.; Álvarez, M.; Cuartero, J. Inheritance of resistance to two-spotted spider mite and glandular leaf trichomes in wild tomato Lycopersicon pimpinellifolium (Jusl.) Mill. J. Am. Soc. Hortic. Sci. 2003, 128, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Firdaus, S.; van Heusden, A.W.; Hidayati, N.; Supena, E.D.J.; Visser, R.G.F.; Vosman, B. Resistance to Bemisia tabaci in tomato wild relatives. Euphytica 2012, 187, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Salinas, M.; Capel, C.; Alba, J.M.; Mora, B.; Cuartero, J.; Fernández-Muñoz, R.; Lozano, R.; Capel, J. Genetic mapping of two QTL from the wild tomato Solanum pimpinellifolium L. controlling resistance against two-spotted spider mite (Tetranychus urticae Koch). Theor. Appl. Genet. 2013, 126, 83–92. [Google Scholar] [CrossRef]
- Vendemiatti, E.; Zsögön, A.; Silva, G.F.F.E.; de Jesus, F.A.; Cutri, L.; Figueiredo, C.R.F.; Tanaka, F.A.O.; Nogueira, F.T.S.; Peres, L.E.P. Loss of type-IV glandular trichomes is a heterochronic trait in tomato and can be reverted by promoting juvenility. Plant Sci. 2017, 259, 35–47. [Google Scholar] [CrossRef]
- Gurr, G.M.; McGrath, D. Effect of plant variety, plant age and photoperiod on glandular pubescence and host-plant resistance to potato moth (Phthorimaea operculella) in Lycopersicon spp. Ann. Appl. Biol. 2001, 138, 221–230. [Google Scholar] [CrossRef]
- Saeidi, Z.; Mallik, B.; Kulkarni, R.S. Inheritance of glandular trichomes and two-spotted spider mite resistance in cross Lycopersicon esculentum “‘Nandi’”and L. pennellii “‘LA2963′”. Euphytica 2007, 154, 231–238. [Google Scholar] [CrossRef]
- Wilkens, R.T.; Shea, G.O.; Halbreich, S.; Stamp, N.E. Resource availability and the trichome defenses of tomato plants. Oecologia 1996, 106, 181–191. [Google Scholar] [CrossRef]
- Weston, P.A.; Johnson, D.A.; Burton, H.T.; Snyder, J.C. Trichome secretion composition, trichome densities, and spider mite resistance of ten accessions of Lycopersicon hirsutum. J. Am. Soc. Hortic. Sci. 1989, 144, 492–498. [Google Scholar]
- Ziarsolo, P.; Hasing, T.; Hilario, R.; Garcia-Carpintero, V.; Blanca, J.; Bombarely, A.; Cañizares, J. K-seq, an affordable, reliable, and open Klenow NGS-based genotyping technology. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mata-Nicolás, E.; Montero-Pau, J.; Gimeno-Paez, E.; Garcia-Carpintero, V.; Ziarsolo, P.; Menda, N.; Mueller, L.A.; Blanca, J.; Cañizares, J.; van der Knaap, E.; et al. Exploiting the diversity of tomato: The development of a phenotypically and genetically detailed germplasm collection. Hortic. Res. 2020, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Maynard, D.N.; Hochmuth, G.J. Knott’s Handbook for Vegetable Growers; John Wiley & Sons: Hoboken, NJ, USA, 2006; ISBN 9780471738282. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 14 October 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Shearer, L.A.; Anderson, L.K.; de Jong, H.; Smit, S.; Goicoechea, J.L.; Roe, B.A.; Hua, A.; Giovannoni, J.J.; Stack, S.M. Fluorescence in situ hybridization and optical mapping to correct scaffold arrangement in the tomato genome. G3 2014, 4, 1395–1405. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. [Google Scholar]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Miao, C.; Fang, J.; Li, D.; Liang, P.; Zhang, X.; Yang, J.; Schnable, J.C.; Tang, H. Genotype-Corrector: Improved genotype calls for genetic mapping in F2 and RIL populations. Sci. Rep. 2018, 8, 10088. [Google Scholar] [CrossRef]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.; Butler, D. R Package ASMap: Efficient Genetic Linkage Map and Diagnosis. J. Stat. Software. 2017, 79, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Delignette-Muller, M.L.; Dutang, C. fitdistrplus: An R Package for Fitting Distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, A.M.; Real, R.; Munoz, A.R.; Brown, J.A. New measures for assessing model equilibrium and prediction mismatch in species distribution models. Divers. Distrib. 2013, 19, 1333–1338. [Google Scholar] [CrossRef]
- Mansfeld, B.N.; Grumet, R. QTLseqr: An R Package for Bulk Segregant Analysis with Next-Generation Sequencing. Plant Genome 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yekutieli, D.; Benjamini, Y. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Potikha, T.; Delmer, D.P. A mutant of Arabidopsis thaliana displaying altered patterns of cellulose deposition. Plant J. 1995, 7, 453–460. [Google Scholar] [CrossRef]
- Gao, Y.; He, C.; Zhang, D.; Liu, X.; Xu, Z.; Tian, Y.; Liu, X.-H.; Zang, S.; Pauly, M.; Zhou, Y.; et al. Two Trichome Birefringence-Like Proteins Mediate Xylan Acetylation, Which Is Essential for Leaf Blight Resistance in Rice. Plant Physiol. 2017, 173, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Ji, W.; McKnight, T.D. Candidate Gene Networks for Acylsugar Metabolism and Plant Defense in Wild Tomato Solanum pennellii. Plant Cell 2020, 32, 81–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.C.; Sussex, I.M. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by gibberellins and photoperiod in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1996, 111, 1321–1328. [Google Scholar] [CrossRef] [Green Version]
- Rakha, M.; Zekeya, N.; Sevgan, S.; Musembi, M.; Ramasamy, S.; Hanson, P. Screening recently identified whitefly/spider mite-resistant wild tomato accessions for resistance to Tuta absoluta. Plant Breed. 2017, 136, 562–568. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-López, M.J.; Moriones, E.; Fernández-Muñoz, R. An Acylsucrose-Producing Tomato Line Derived from the Wild Species Solanum pimpinellifolium Decreases Fitness of the Whitefly Trialeurodes vaporariorum. Insects 2020, 11. [Google Scholar] [CrossRef]
- Leckie, B.M.; De Jong, D.M.; Mutschler, M.A. Quantitative trait loci regulating sugar moiety of acylsugars in tomato. Mol. Breed. 2013, 31, 957–970. [Google Scholar] [CrossRef]
- Smeda, J.R.; Schilmiller, A.L.; Last, R.L.; Mutschler, M.A. Introgression of acylsugar chemistry QTL modifies the composition and structure of acylsugars produced by high-accumulating tomato lines. Mol. Breed. 2016, 36, 160. [Google Scholar] [CrossRef]
- Smeda, J.R.; Schilmiller, A.L.; Anderson, T.; Ben-Mahmoud, S.; Ullman, D.E.; Chappell, T.M.; Kessler, A.; Mutschler, M.A. Combination of Acylglucose QTL reveals additive and epistatic genetic interactions and impacts insect oviposition and virus infection. Mol. Breed. 2017, 38, 3. [Google Scholar] [CrossRef]
- Freitas, J.A.; Maluf, W.R.; das Graças Cardoso, M.; Gomes, L.A.A.; Bearzotti, E. Inheritance of foliar zingiberene contents and their relationship to trichome densities and whitefly resistance in tomatoes. Euphytica 2002, 127, 275–287. [Google Scholar] [CrossRef]
- Vosman, B.; Kashaninia, A.; Van’t Westende, W.; Meijer-Dekens, F.; van Eekelen, H.; Visser, R.G.F.; de Vos, R.C.H.; Voorrips, R.E. QTL mapping of insect resistance components of Solanum galapagense. Theor. Appl. Genet. 2019, 132, 531–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliepaard, C.; Bas, N.; van Heusden, S.; Kos, J.; Pet, G.; Verkerk, R.; Vrieunk, R.; Zabel, P.; Lindhout, P. Mapping of QTLs for glandular trichome densities and Trialeurodes vaporariorum (greenhouse whitefly) resistance in an F 2 from Lycopersicon esculentum× Lycopersicon hirsutum f. glabratum. Heredity 1995, 75, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Fambrini, M.; Pugliesi, C. The Dynamic Genetic-Hormonal Regulatory Network Controlling the Trichome Development in Leaves. Plants 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Doroshkov, A.V.; Konstantinov, D.K.; Afonnikov, D.A.; Gunbin, K.V. The evolution of gene regulatory networks controlling Arabidopsis thaliana L. trichome development. BMC Plant Biol. 2019, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Kirik, V.; Lee, M.M.; Wester, K.; Herrmann, U.; Zheng, Z.; Oppenheimer, D.; Schiefelbein, J.; Hulskamp, M. Functional diversification of MYB23 and GL1 genes in trichome morphogenesis and initiation. Development 2005, 132, 1477–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, L.; Xia, K.; Chen, J.-G.; Wang, S. Functional characterization of TRICHOMELESS2, a new single-repeat R3 MYB transcription factor in the regulation of trichome patterning in Arabidopsis. BMC Plant Biol. 2011, 11, 176. [Google Scholar] [CrossRef] [Green Version]
- Ó’Maoiléidigh, D.S.; Wuest, S.E.; Rae, L.; Raganelli, A.; Ryan, P.T.; Kwaśniewska, K.; Das, P.; Lohan, A.J.; Loftus, B.; Graciet, E.; et al. Control of reproductive floral organ identity specification in Arabidopsis by the C function regulator AGAMOUS. Plant Cell 2013, 25, 2482–2503. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. SNPs | No. Bin Markers | Length | Avg. Spacing | Max. Spacing | |
|---|---|---|---|---|---|
| SL2.50ch01 | 23,537 | 1248 | 153.4 | 0.1 | 3.7 |
| SL2.50ch02 | 10,368 | 662 | 130.3 | 0.2 | 1.8 |
| SL2.50ch03 | 8300 | 479 | 165.2 | 0.3 | 8.5 |
| SL2.50ch04 | 9482 | 534 | 123.9 | 0.2 | 1.3 |
| SL2.50ch05 | 15,374 | 856 | 121.4 | 0.1 | 7.9 |
| SL2.50ch06 | 8375 | 552 | 123.7 | 0.2 | 11.2 |
| SL2.50ch07 | 17,919 | 770 | 110.9 | 0.1 | 1.9 |
| SL2.50ch08 | 7555 | 563 | 164.7 | 0.3 | 13.3 |
| SL2.50ch09 | 11,342 | 725 | 125.9 | 0.2 | 3.7 |
| SL2.50ch10 | 14,855 | 788 | 119.9 | 0.2 | 5.2 |
| SL2.50ch11 | 9925 | 567 | 101.8 | 0.2 | 2.8 |
| SL2.50ch12 | 10,294 | 541 | 159.2 | 0.3 | 6.2 |
| Overall | 147,326 | 8285 | 1600.4 | 0.2 | 13.3 |
| QTL Mapping (SLC Family) | |||||
| Phenotype | Chr | Start | Max | End | LOD |
| Trichome density 6th week | SL2.50ch09 | 2,055,839 | 2,542,786 | 4,250,325 | 6.24 |
| SL2.50ch11 | 54,298,873 | 56,297,460 | 56,297,460 | 5.62 | |
| Trichome density 7th week | SL2.50ch09 | 2,078,180 | 3,756,370 | 4,250,325 | 6.33 |
| SL2.50ch11 | 55,093,420 | 56,297,460 | 56,297,460 | 3.69 | |
| Trichome density 8th week | SL2.50ch09 | 1,966,640 | 2,309,105 | 4,371,756 | 5.36 |
| SL2.50ch11 | 55,548,327 | 56,297,460 | 56,297,460 | 3.83 | |
| Association (SLC Family) | |||||
| Phenotype | Chromosome | Start | Max | End | Adj. pval. |
| Trichome density 6th week | SL2.50ch09 | 2,309,105 | 2,542,786 | 2,545,171 | 0.0090 |
| Trichome density 7th week | SL2.50ch09 | 2,078,180 | 3,727,914 | 4,969,831 | 7.7·10−6 |
| Trichome density 8th week | SL2.50ch06 | 8008 | 154,414 | 615,838 | 0.0002 |
| SL2.50ch09 | 2,069,769 | 2,492,691 | 5,693,896 | 4.6·10−5 | |
| QTLseq | |||||
| Mapping Population | Chromosome | Start | Max | End | ΔSNP |
| SLC family | SL2.50ch09 | 3,042,382 | 3,966,023 | 4,560,795 | 0.57 |
| SLL family | SL2.50ch05 | 9,189,958 | 33,460,771 | 59,293,229 | 0.52 |
| SL2.50ch06 | 23,628 | 23,628 | 943,353 | 0.46 | |
| SL2.50ch09 | 2,105,569 | 2,865,099 | 4,800,975 | 0.56 | |
| QTL Mapping | |||||||
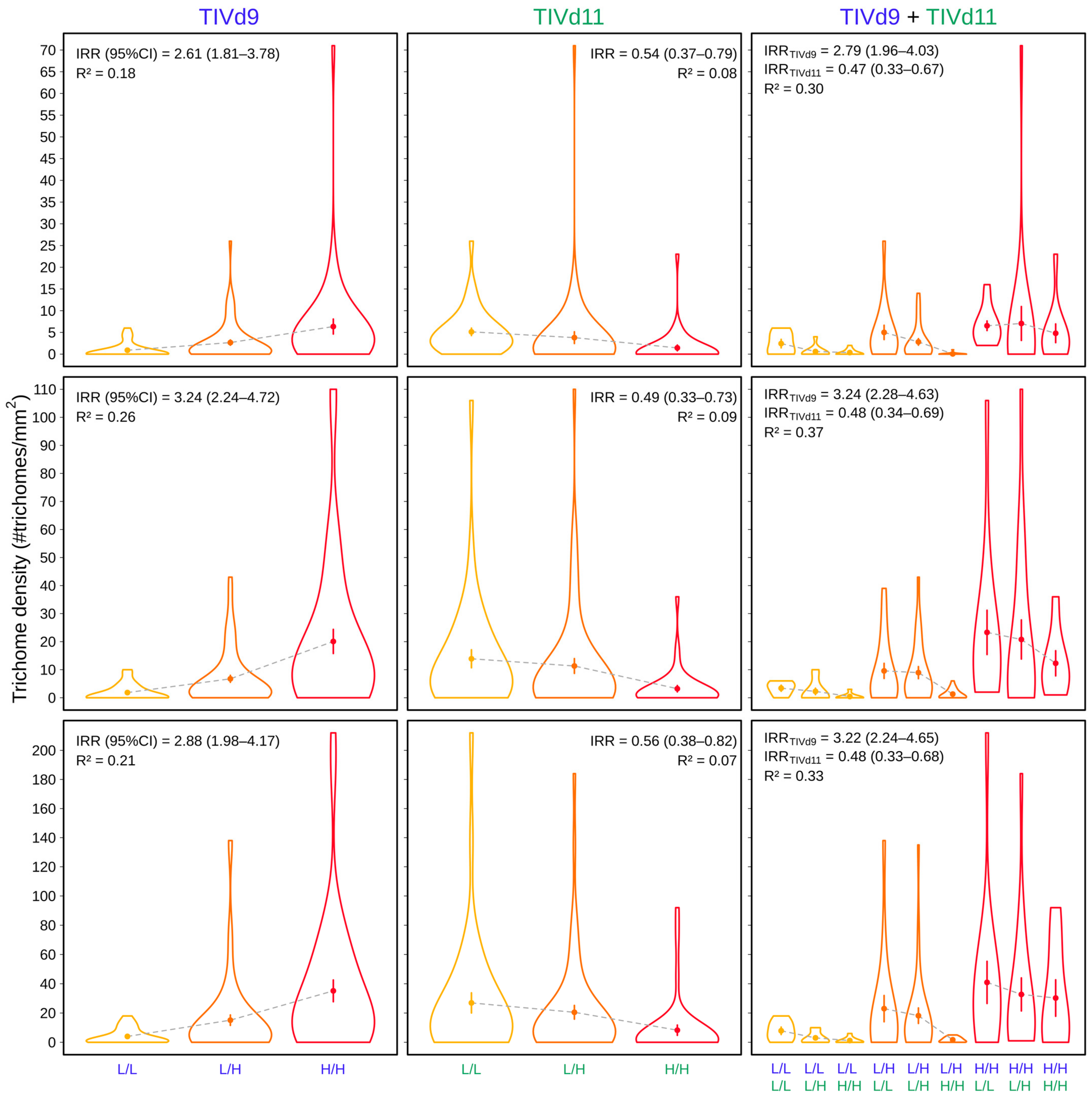
| Phenotype | Model | GOF | R2 | D2 | Variable | IRR (95% CI) | Sig |
| TD 6th w | TIVd9 | 0.361 | 0.18 | 0.16 | SL2.50ch09_2542786 | 2.61 (1.81–3.78) | *** |
| TIVd11 | 0.348 | 0.08 | 0.07 | SL2.50ch11_56297460 | 0.54 (0.37–0.78) | *** | |
| TIVd9 + TIVd11 | 0.386 | 0.30 | 0.26 | SL2.50ch09_2542786 | 2.79 (1.96–4.03) | *** | |
| SL2.50ch11_56297460 | 0.47 (0.33–0.67) | *** | |||||
| TD 7th w | TIVd9 | 0.145 | 0.26 | 0.21 | SL2.50ch09_3756370 | 3.24 (2.24–4.72) | *** |
| TIVd11 | 0.144 | 0.09 | 0.08 | SL2.50ch11_56297460 | 0.49 (0.33–0.73) | *** | |
| TIVd9 + TIVd11 | 0.149 | 0.37 | 0.29 | SL2.50ch09_3756370 | 3.24 (2.28–4.63) | *** | |
| SL2.50ch11_56297460 | 0.48 (0.34–0.69) | *** | |||||
| TD 8th w | TIVd9 | 0.084 | 0.21 | 0.17 | SL2.50ch09_2309105 | 2.88 (1.98–4.17) | *** |
| TIVd11 | 0.076 | 0.07 | 0.06 | SL2.50ch11_56297460 | 0.56 (0.38–0.82) | *** | |
| TIVd9 + TIVd11 | 0.091 | 0.33 | 0.26 | SL2.50ch09_2309105 | 3.22 (2.24–4.65) | *** | |
| SL2.50ch11_56297460 | 0.48 (0.33–0.68) | *** | |||||
| Association | |||||||
| Phenotype | Model | GOF | R2 | D2 | Variable | IRR (95% CI) | Sig |
| TD 6th w | TIVd9 | 0.361 | 0.18 | 0.16 | SL2.50ch09_2542786 | 2.61 (1.81–3.78) | *** |
| TD 7th w | TIVd9 | 0.147 | 0.25 | 0.20 | SL2.50ch09_3727914 | 3.19 (2.21–4.64) | *** |
| TD 8th w | TIVd6 | 0.090 | 0.22 | 0.17 | SL2.50ch06_154414 | 2.47 (1.79–3.41) | *** |
| TIVd9 | 0.085 | 0.22 | 0.18 | SL2.50ch09_2492691 | 2.94 (2.03–4.26) | *** | |
| TIVd6 + TIVd9 | 0.085 | 0.32 | 0.25 | SL2.50ch06_154414 | 1.86 (1.33–2.60) | *** | |
| SL2.50ch09_2492691 | 2.16 (1.44–3.22) | *** | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata-Nicolás, E.; Montero-Pau, J.; Gimeno-Paez, E.; García-Pérez, A.; Ziarsolo, P.; Blanca, J.; van der Knaap, E.; Díez, M.J.; Cañizares, J. Discovery of a Major QTL Controlling Trichome IV Density in Tomato Using K-Seq Genotyping. Genes 2021, 12, 243. https://doi.org/10.3390/genes12020243
Mata-Nicolás E, Montero-Pau J, Gimeno-Paez E, García-Pérez A, Ziarsolo P, Blanca J, van der Knaap E, Díez MJ, Cañizares J. Discovery of a Major QTL Controlling Trichome IV Density in Tomato Using K-Seq Genotyping. Genes. 2021; 12(2):243. https://doi.org/10.3390/genes12020243
Chicago/Turabian StyleMata-Nicolás, Estefanía, Javier Montero-Pau, Esther Gimeno-Paez, Ana García-Pérez, Peio Ziarsolo, José Blanca, Esther van der Knaap, María José Díez, and Joaquín Cañizares. 2021. "Discovery of a Major QTL Controlling Trichome IV Density in Tomato Using K-Seq Genotyping" Genes 12, no. 2: 243. https://doi.org/10.3390/genes12020243
APA StyleMata-Nicolás, E., Montero-Pau, J., Gimeno-Paez, E., García-Pérez, A., Ziarsolo, P., Blanca, J., van der Knaap, E., Díez, M. J., & Cañizares, J. (2021). Discovery of a Major QTL Controlling Trichome IV Density in Tomato Using K-Seq Genotyping. Genes, 12(2), 243. https://doi.org/10.3390/genes12020243