
Transcriptomic Diversity in the Livers of South African Sardines Participating in the Annual Sardine Run

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Specimen Collection
2.3. Nucleic Acid Extraction, Genomic Library Preparation, and Sequencing
2.4. Transcriptome Assembly, Functional Annotation, and Variant Calling
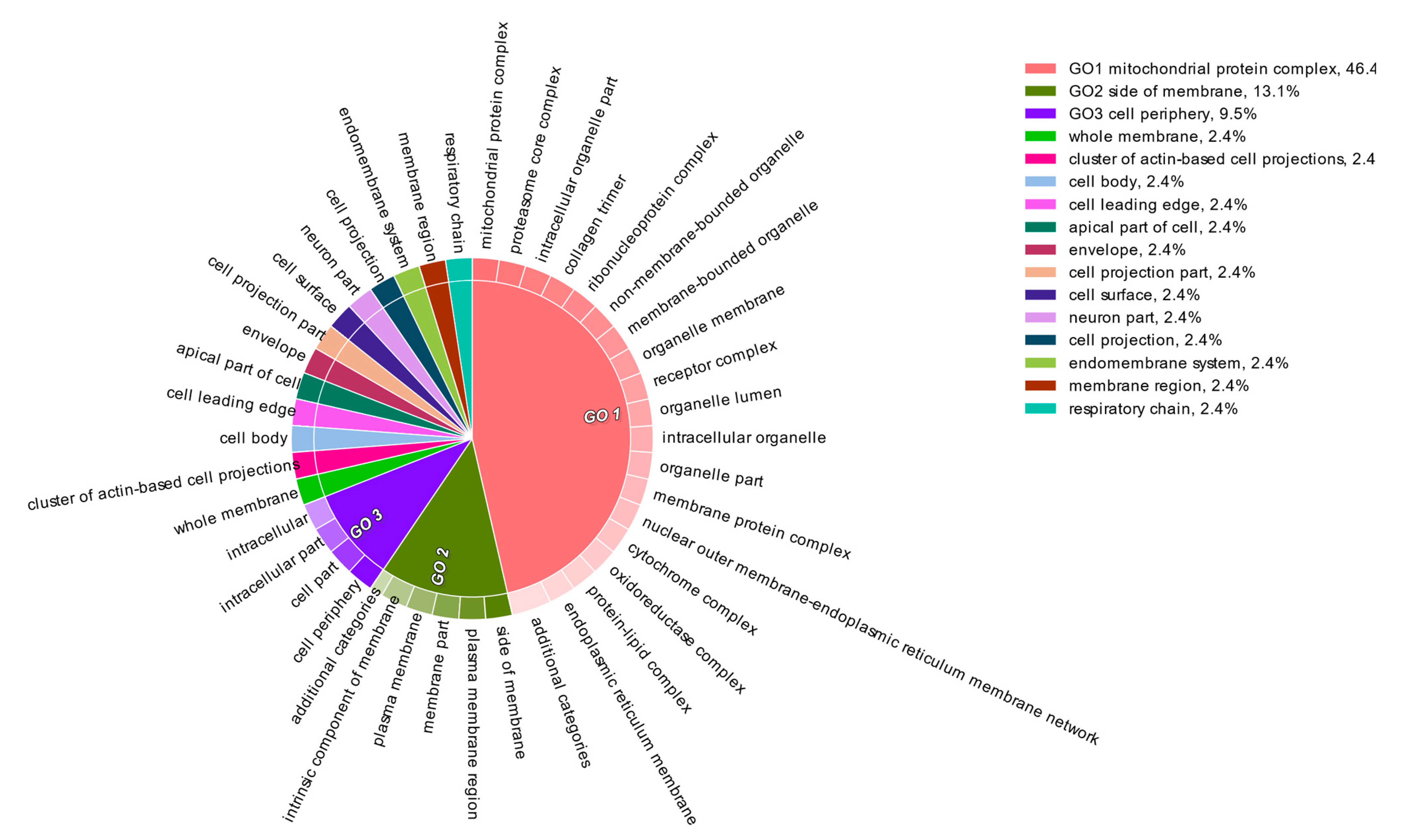
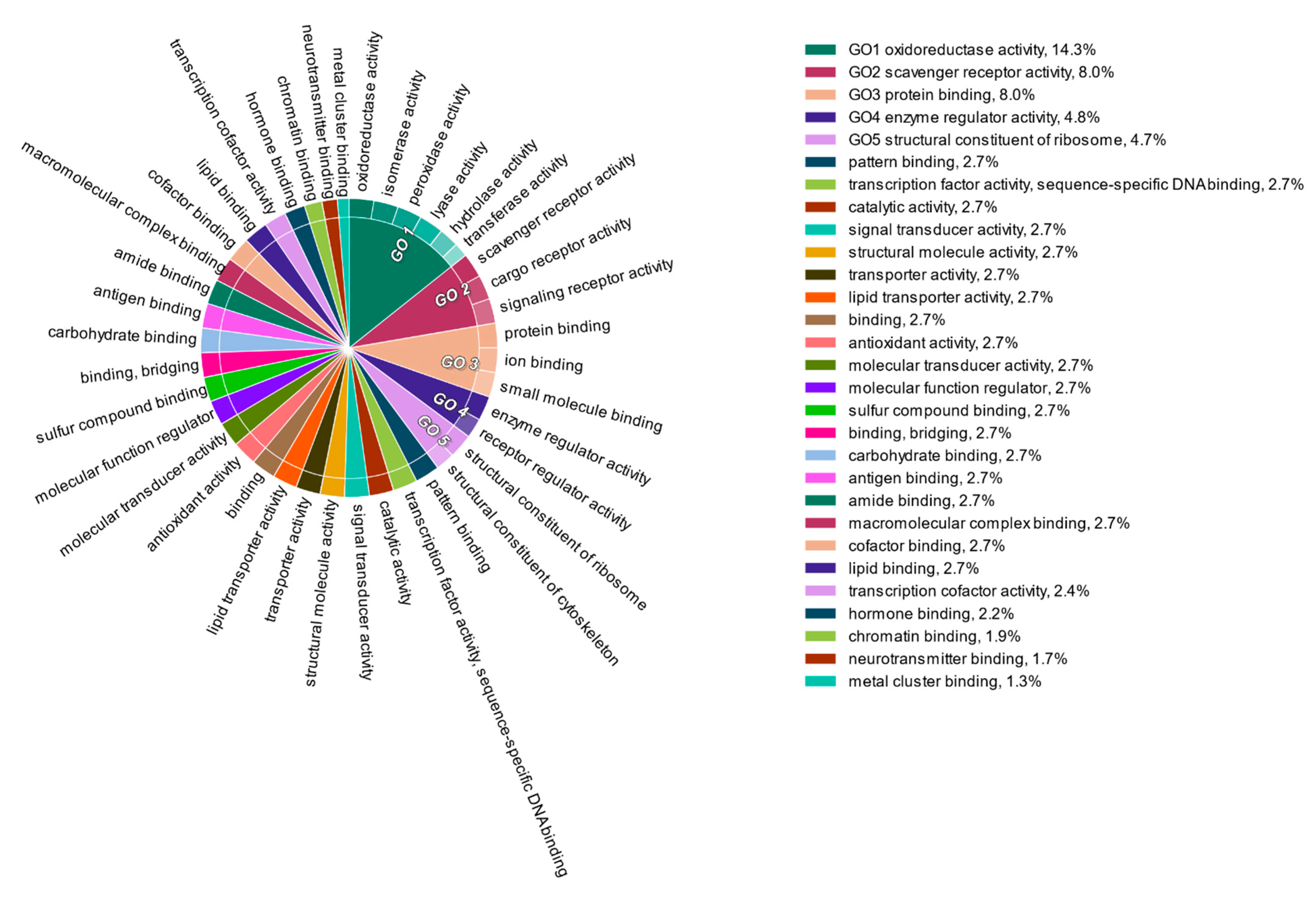
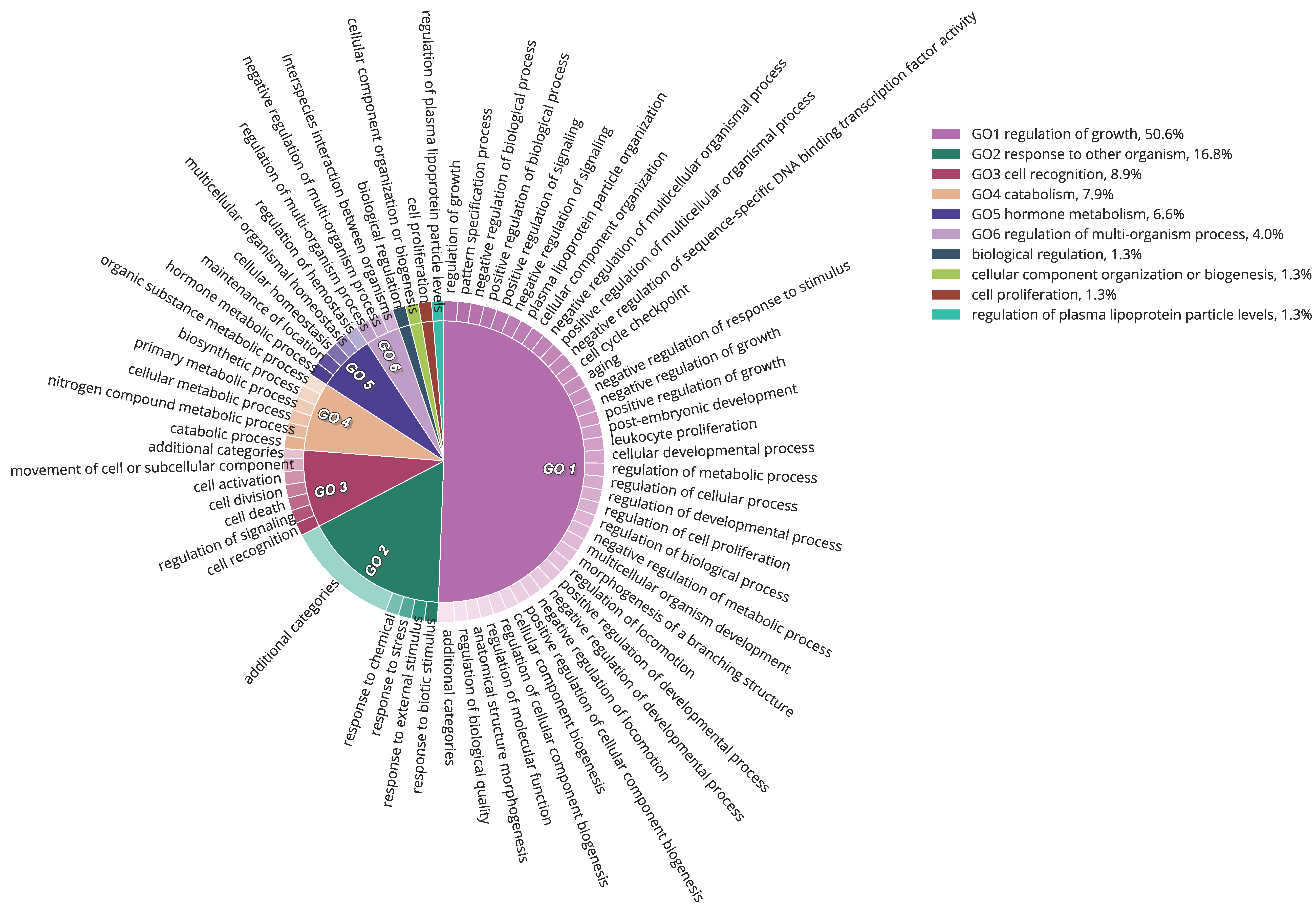
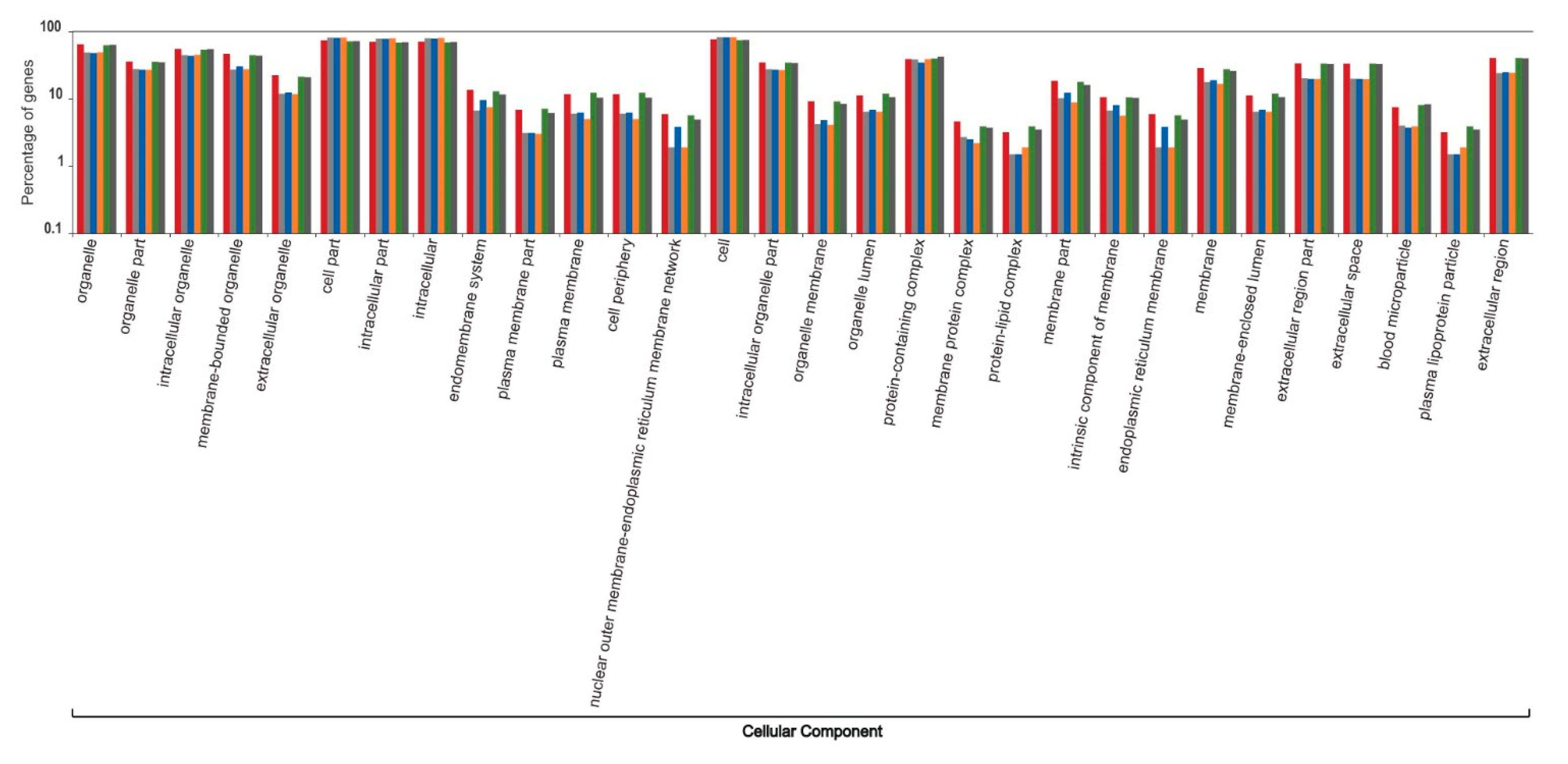
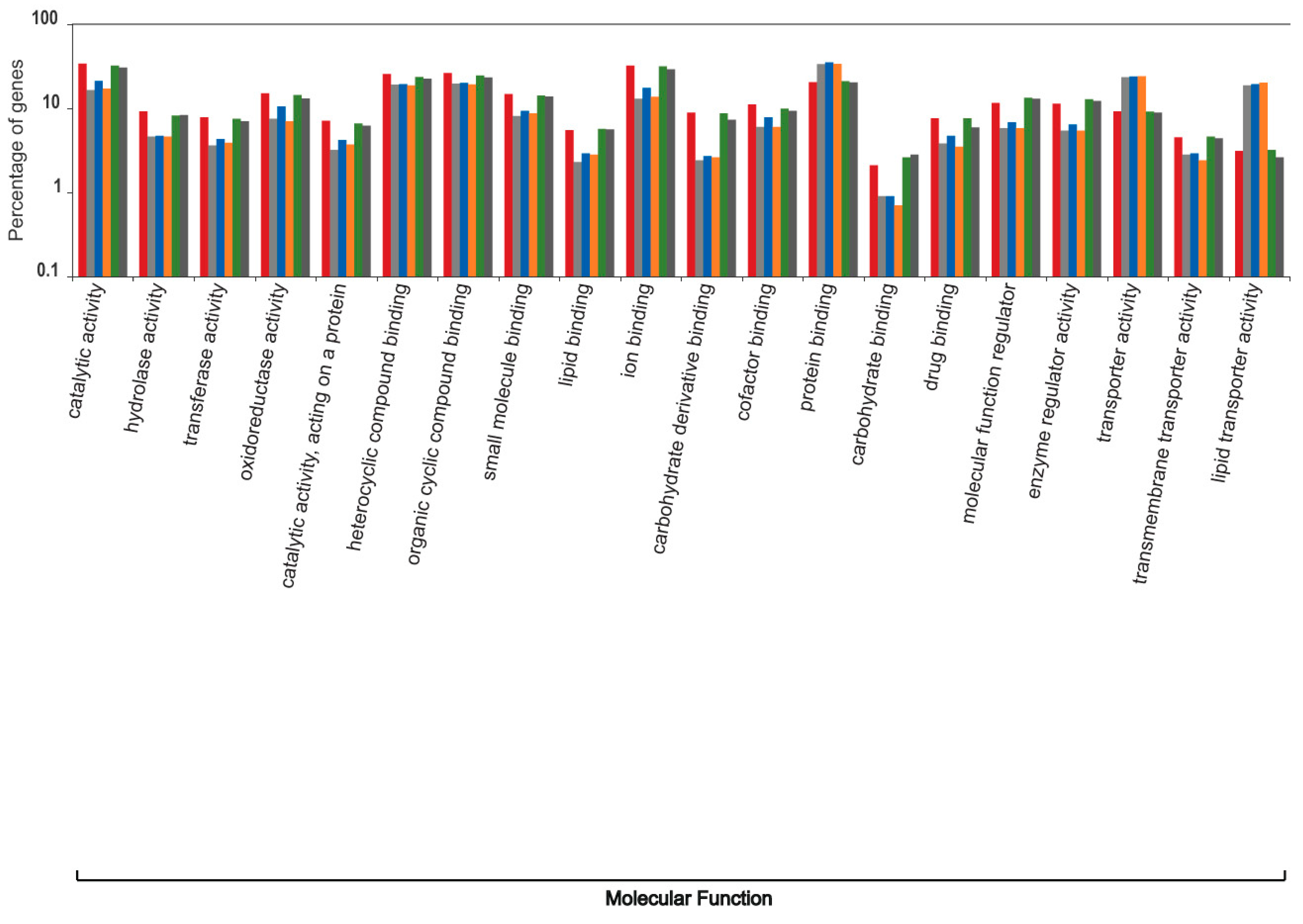
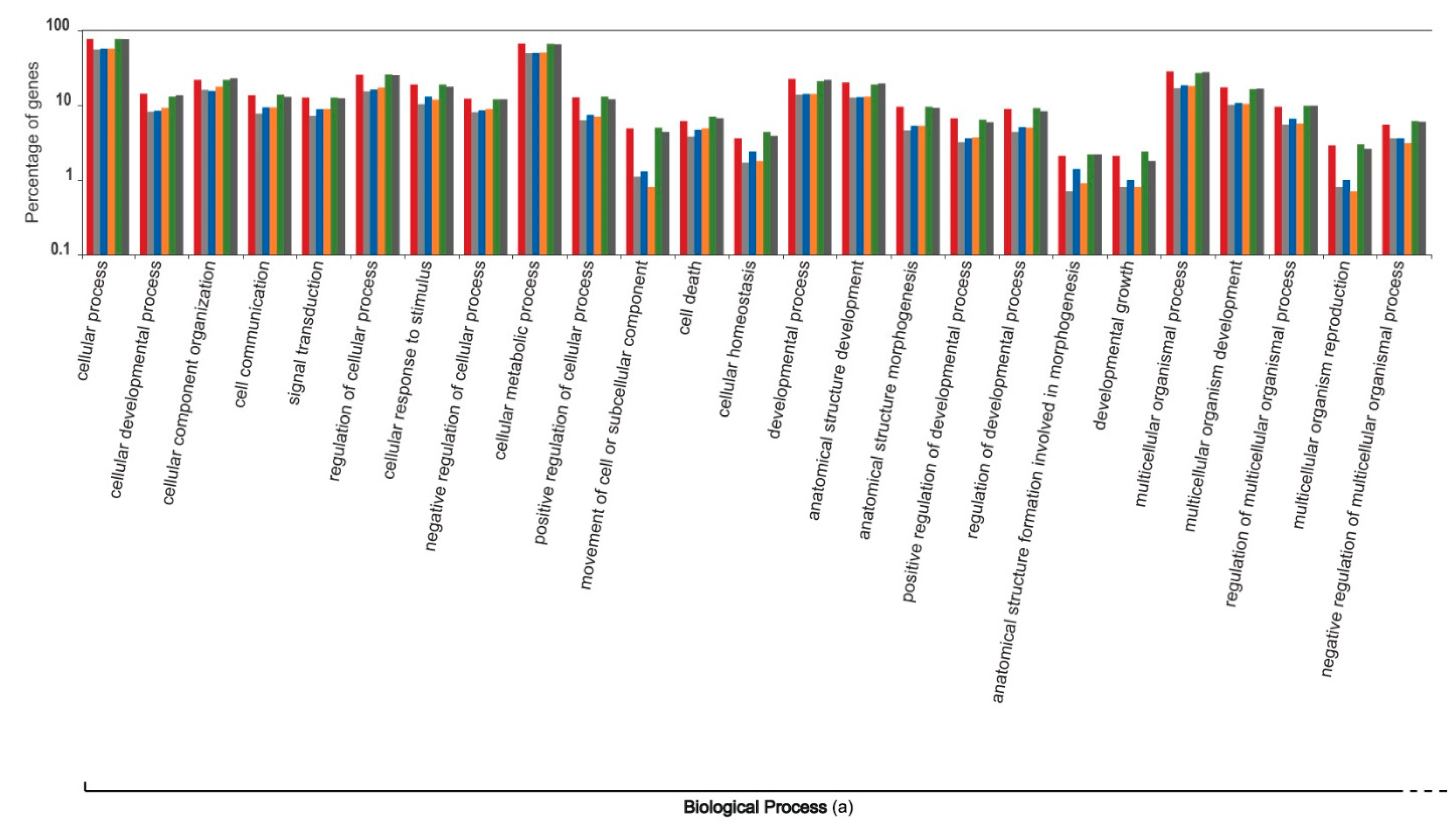
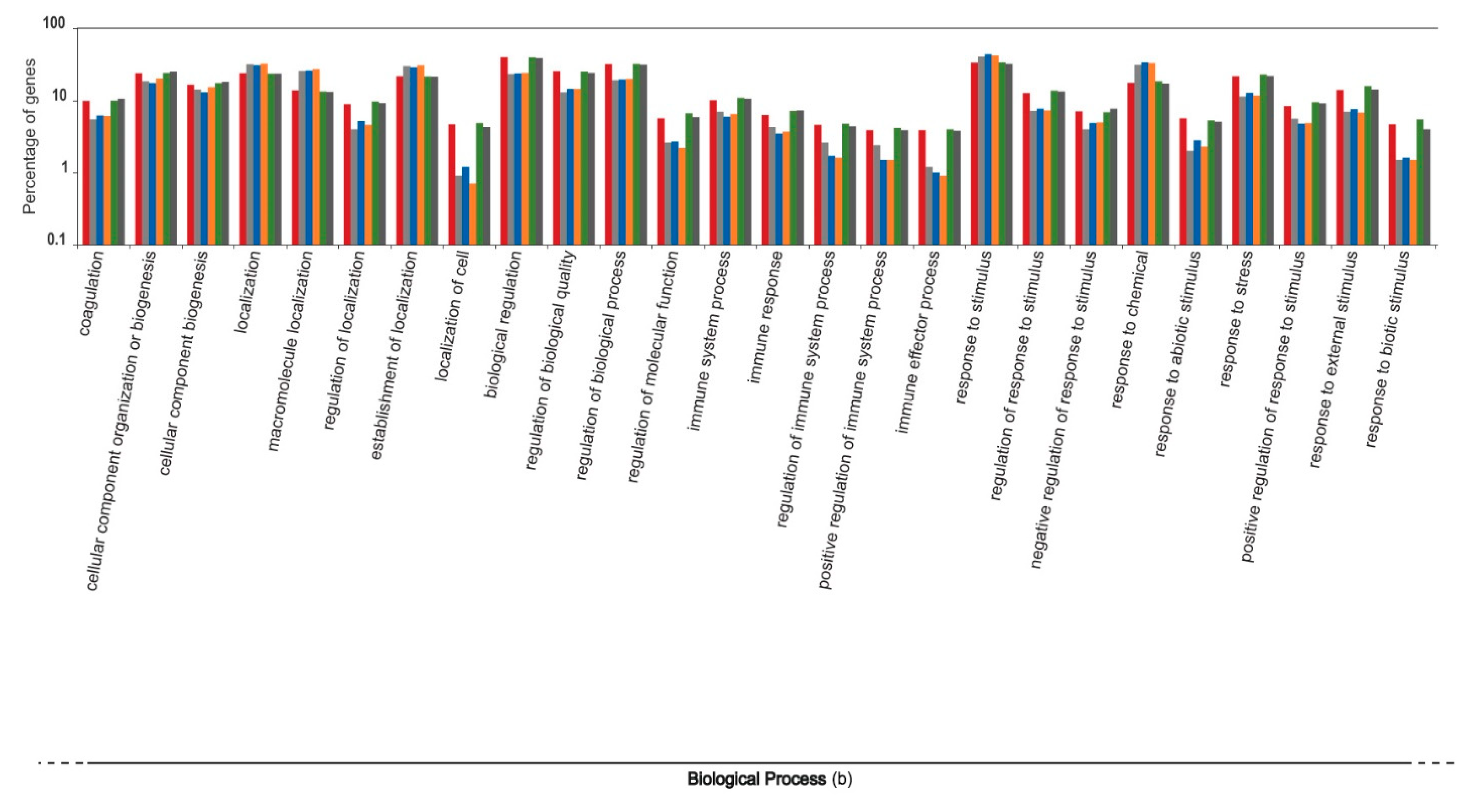
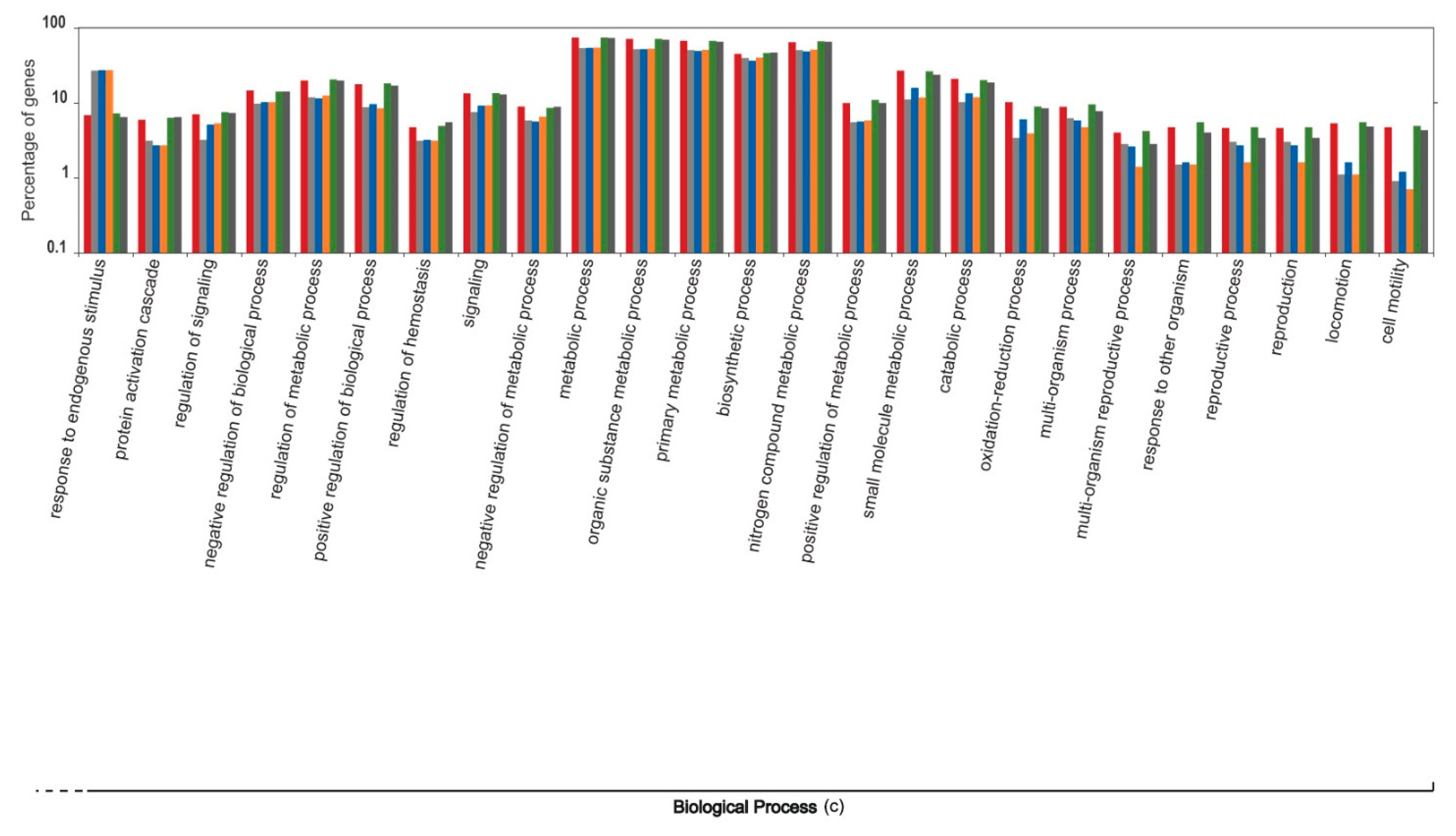
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grantham, H.S.; Game, E.T.; Lombard, A.T.; Hobday, A.J.; Richardson, A.J.; Beckley, L.E.; Pressey, R.L.; Huggett, J.A.; Coetzee, J.C.; van der Lingen, C.D.; et al. Accommodating dynamic oceanographic processes and pelagic biodiversity in marine conservation planning. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensor, E.; Couzin, I.D.; James, R.; Krause, J. Modelling density-dependent fish shoal distributions in the laboratory and field. Oikos 2005, 110, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Webster, M.M.; Goldsmith, J.; Ward, A.J.W.; Hart, P.J.B. Habitat-specific chemical cues influence association preferences and shoal cohesion in fish. Behav. Ecol. Sociobiol. 2007, 62, 273–280. [Google Scholar] [CrossRef]
- Cattelan, S.; Griggio, M. Within-shoal phenotypic homogeneity overrides familiarity in a social fish. Behav. Ecol. Sociobiol. 2020, 74. [Google Scholar] [CrossRef]
- van der Lingen, C.D.; Coetzee, J.C.; Hutchings, L. Overview of the KwaZulu-Natal sardine run. Afr. J. Mar. Sci. 2010, 32, 271–277. [Google Scholar] [CrossRef]
- Hutchings, L.; Morris, T.; van der Lingen, C.D.; Lamberth, S.J.; Connell, A.D.; Taljaard, S.; van Niekerk, L. Ecosystem considerations of the kwaZulu-Natal sardine run. Afr. J. Mar. Sci. 2010, 32, 413–421. [Google Scholar] [CrossRef]
- Myeza, J.; Mason, R.B.; Peddemors, V.M. Socio-economic implications of the kwaZulu-Natal sardine run for local indigenous communities. Afr. J. Mar. Sci. 2010, 32, 399–404. [Google Scholar] [CrossRef]
- Andrew, S.C.; Primmer, C.R.; Debes, P.V.; Erkinaro, J.; Verta, J.P. The Atlantic salmon whole blood transcriptome and how it relates to major locus maturation genotypes and other tissues. Mar. Genom. 2020, 100809. [Google Scholar] [CrossRef]
- Ryu, T.; Kim, J.G.; Lee, J.; Yu, O.H.; Yum, S.; Kim, D.; Woo, S. First transcriptome assembly of a newly dis-covered vent mussel, Gigantidas vrijenhoeki, at Onnuri Vent Field on the northern Central Indian Ridge. Mar. Genom. 2020, 100819. [Google Scholar] [CrossRef]
- Pinosio, S.; Fratini, S.; Cannicci, S.; Oliva, M.; Pretti, C.; Baratti, M. De novo transcriptome assembly for Pach-ygrapsus marmoratus, an intertidal brachyuran crab. Mar. Genom. 2020, 100792. [Google Scholar] [CrossRef]
- Gnocchi, K.G.; Boldrini-França, J.; Gomes, L.C.; Chippari-Gomes, A.R. De novo assembly and annotation of the transcriptome of Astyanax lacustris liver unveil candidate genes to monitor response to environmental stress. Mar. Genom. 2020, 54, 100784. [Google Scholar] [CrossRef]
- Emami-Khoyi, A.; Parbhu, S.P.; Ross, J.G.; Murphy, E.C.; Bothwell, J.; Monsanto, D.M.; van Vuuren, B.J.; Teske, P.R.; Paterson, A.M. De novo transcriptome assembly and annotation of liver and brain tissues of common brushtail possums (Trichosurus vulpecula) in New Zealand: Transcriptome diversity after decades of population control. Genes 2020, 11, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallero, S.; Lombardo, F.; Salvemini, M.; Pizzarelli, A.; Cantacessi, C.; D’Amelio, S. Comparative transcriptomics reveals clues for differences in pathogenicity between Hysterothylacium aduncum, Anisakis simplex sensu stricto and Anisakis pegreff. Genes 2020, 11, 321. [Google Scholar] [CrossRef] [Green Version]
- Tsehay, S.; Ortiz, R.; Johansson, E.; Bekele, E. New transcriptome-based SNP markers for Noug (Guizotia abyssinica) and their conversion to KASP markers for population genetics analyses. Genes 2020, 11, 1373. [Google Scholar] [CrossRef] [PubMed]
- Benítez, R.; Trakooljul, N.; Núñez, Y.; Isabel, B.; Murani, E.; De Mercado, E.; Gómez-Izquierdo, E.; García-Casco, J.; López-Bote, C.; Wimmers, K.; et al. Breed, diet, and interaction effects on adipose tissue transcriptome in iberian and duroc pigs fed different energy sources. Genes 2019, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- van Dyk, J.C.; Pieterse, G.M.; van Vuren, J.H.J. Histological changes in the liver of Oreochromis mossambicus (Cichlidae) after exposure to cadmium and zinc. Ecotoxicol. Environ. Saf. 2007, 66, 432–440. [Google Scholar] [CrossRef]
- Ren, M.; Li, J.; Xue, R.; Wang, Z.; Coll, S.L.; Meng, Q. Liver function and energy metabolism in hepatocellular carcinoma developed in patients with hepatitis B-related cirrhosis. Medicine 2019, 98, e15528. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, D.; Das, S.; Kumar, V. Hypothalamic and liver transcriptome from two crucial life-history stages in a migratory songbird. Exp. Physiol. 2018, 103, 559–569. [Google Scholar] [CrossRef]
- Horton, W.J.; Jensen, M.; Sebastian, A.; Praul, C.A.; Albert, I.; Bartell, P.A. Transcriptome analyses of heart and liver reveal novel pathways for regulating songbird migration. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Swarup, V.; Le, H.; Kumar, V. Transcriptional Signatures in Liver Reveal Metabolic Adaptations to Seasons in Migratory Blackheaded Buntings. Front. Physiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A tissue-mapped axolotl De Novo transcriptome enables identification of limb regeneration factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Stærfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. Busco: Assessing genome assembly and annotation completeness in gene prediction. In Gene prediction; Humana: New York, NY, USA, 2019; pp. 227–245. [Google Scholar]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAM tools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequenc-ing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene Ontology: Tool for the Unification of Biology. Nat. Genet 2000, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimand, J.; Kull, M.; Peterson, H.; Hansen, J.; Vilo, J. G:Profiler-a web-based toolset for functional profiling of gene lists from large-scale experiments. Nucleic Acids Res. 2007, 35, 193–200. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A. WEGO 2.0: AWeb Tool for Analyzing and Plotting Go Annotations, 2018 Update. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsova, I.; Lugmayr, A.; Siira, S.J.; Rackham, O.; Filipovska, A. CirGO: An alternative circular way of visualizing gene ontology terms. BMC Bioinform. 2019, 20, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strange, R.C.; Spiteri, M.A.; Ramachandran, S.; Fryer, A.A. Glutathione-S-transferase family of enzymes. Mutat. Res. Mol. Mech. Mutagen. 2001, 482, 21–26. [Google Scholar] [CrossRef]
- Leaver, M.J.; Wright, J.; Hodgson, P.; Boukouvala, E.; George, S.G. Piscine UDP-glucuronosyltransferase 1B. Aquat. Toxicol. 2007, 84, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzahouane, H.; Barour, C.; Sleimi, N.; Ouali, K. Multi-biomarkers approach to the assessment of the south-eastern Mediterranean Sea health status: Preliminary study on Stramonita haemastoma used as a bioindicator for metal contamination. Chemosphere 2018, 207, 725–741. [Google Scholar] [CrossRef]
- Faheem, M.; Lone, K.P. Oxidative stress and histopathologic biomarkers of exposure to bisphenol-A in the freshwater fish, Ctenopharyngodon idella. Braz. J. Pharm. Sci. 2018, 53, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Samaraweera, A.V.; Sandamalika, W.M.G.; Liyanage, D.S.; Lee, S.; Priyathilaka, T.T.; Lee, J. Molecular char-acterization and functional analysis of glutathione S-transferase kappa 1 (GSTκ1) from the big belly seahorse (Hippocampus abdominalis): Elucidation of its involvement in innate immune responses. Fish Shellfish Immunol. 2019, 92, 356–366. [Google Scholar] [CrossRef]
- Chen, B.; Peng, W.; Xu, J.; Feng, J.; Dong, C.; Xu, P.; Chen, B. Genomic Analysis of Glutathione S-transferases. Pak. J. Zool. 2017, 49, 1437–1448. [Google Scholar] [CrossRef]
- Glisic, B.; Mihaljevic, I.; Popovic, M.; Zaja, R.; Loncar, J.; Fent, K.; Kovacevic, R.; Smital, T. Characterization of glutathione-S-transferases in zebrafish (Danio rerio). Aquat. Toxicol. 2015, 158, 50–62. [Google Scholar] [CrossRef]
- Pérez-López, M.; Anglade, P.; Bec-Ferté, M.P.; Debrauwer, L.; Perdu, E.; Cravedi, J.P.; Rouimi, P. Characterization of hepatic and extrahepatic glutathione S-transferases in rainbow trout (Oncorhynchus mykiss) and their induction by 3,3’,4,4’-tetrachlorobiphenyl. Fish Physiol. Biochem. 2000, 22, 21–32. [Google Scholar] [CrossRef]
- Celius, T.; Walther, B.T. Oogenesis in Atlantic salmon (Salmo salar) occurs by zonagenesis preceding vitellogenesis in vivo and in vitro. J. Endocrinol. 1998, 158, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Conner, S.J.; Hughes, D.C. Analysis of fish ZP1/ZPB homologous genes—Evidence for both genome duplication and species-specific amplification models of evolution. Reproduction 2003, 126, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, Z.; Gao, L.Y.; Colorni, A.; Ucko, M.; Fang, S.; Du, S.J. A transgenic zebrafish model for monitoring xbp1 splicing and endoplasmic reticulum stress in vivo. Mech. Dev. 2015, 137, 33–44. [Google Scholar] [CrossRef]
- Pacitti, D.; Lawan, M.M.; Feldmann, J.; Sweetman, J.; Wang, T.; Martin, S.A.M.; Secombes, C.J. Impact of selenium supplementation on fish antiviral responses: A whole transcriptomic analysis in rainbow trout (Oncorhynchus mykiss) fed supranutritional levels of Sel-Plex®. BMC Genom. 2016, 17, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, I.C.; Hsiao, Y.C.; Sun, H.S.; Chen, T.M.; Lee, S.J. MicroRNAs regulate gene plasticity during cold shock in zebrafish larvae. BMC Genom. 2016, 17, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, G.; Hofweber, R.; Kremer, W.; Kalbitzer, H.R. Structure and function of bacterial cold shock proteins. Cell. Mol. Life Sci. 2007, 64, 1457–1470. [Google Scholar] [CrossRef] [PubMed]
- Chaikam, V.; Karlson, D.T. Comparison of structure, function and regulation of plant cold shock domain proteins to bacterial and animal cold shock domain proteins. BMB Rep. 2010, 43, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, N.K.; Murphy, A.; Costa, M. Arsenic methyltransferase and methylation of inorganic arsenic. Biomolecules 2020, 10, 1351. [Google Scholar] [CrossRef] [PubMed]
- Tierbach, A.; Groh, K.J.; Schönenberger, R.; Schirmer, K.; Suter, M.J.F. Glutathione S-Transferase protein expression in different life stages of zebrafish (Danio rerio). Toxicol. Sci. 2018, 162, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Micheletti, S.J.; Hess, J.E.; Zendt, J.S.; Narum, S.R. Selection at a genomic region of major effect is responsible for evolution of complex life histories in anadromous Steelhead. BMC Evol. Biol. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Jindal, R.; Sinha, R.; Brar, P. Evaluating the protective efficacy of Silybum marianum against deltamethrin in-duced hepatotoxicity in piscine model. Environ. Toxicol. Pharmacol. 2019, 66, 62–68. [Google Scholar] [CrossRef]
- Hygum, T.L.; Fobian, D.; Kamilari, M.; Jørgensen, A.; Schiøtt, M.; Grosell, M.; Møbjerg, N. Comparative investigation of copper tolerance and identification of putative tolerance related genes in tardigrades. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Le Corre, M.; Dussault, C.; Côté, S.D. Weather conditions and variation in timing of spring and fall migrations of migratory caribou. J. Mammal. 2016, 98, gyw177. [Google Scholar] [CrossRef]
- Rakhimberdiev, E.; Duijns, S.; Karagicheva, J.; Camphuysen, C.J.; Dekinga, A.; Dekker, R.; Gavrilov, A.; ten Horn, J.; Jukema, J.; Saveliev, A.; et al. Fuelling conditions at staging sites can mitigate Arctic warming effects in a migratory bird. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.M.; Laughlin, A.J.; Hall, R.J. The response of migratory populations to phenological change: A Migratory Flow Network modelling approach. J. Anim. Ecol. 2016, 85, 648–659. [Google Scholar] [CrossRef]
- Mhlongo, N.; Yemane, D.; Hendricks, M.; van der Lingen, C.D. Have the spawning habitat preferences of an-chovy (Engraulis encrasicolus) and sardine (Sardinops sagax) in the southern Benguela changed in recent years. Fish. Oceanogr. 2015, 24, 1–14. [Google Scholar] [CrossRef]
- Rouault, M.; Pohl, B.; Penven, P. Coastal oceanic climate change and variability from 1982 to 2009 around South Africa. African J. Mar. Sci. 2010, 32, 237–246. [Google Scholar] [CrossRef]
- Mohanty, B.; Mohanty, S.; Sahoo, J. Climate Change: Impacts on Fisheries and Aquaculture. In Climate Change and Variability; Suzanne Simard, Ed.; Intech: Rijeka, Croatia, 2010; pp. 120–138. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Pathway | Occurrences |
|---|---|
| Signal transduction | 3812 |
| Global and overview maps | 3579 |
| Immune system | 1475 |
| Endocrine system | 1473 |
| Transport and catabolism | 1424 |
| Carbohydrate metabolism | 774 |
| Nervous system | 750 |
| Amino acid metabolism | 647 |
| Lipid metabolism | 487 |
| Cellular community—eukaryotes | 468 |
| Folding, sorting, and degradation | 448 |
| Digestive system | 442 |
| Translation | 416 |
| Energy metabolism | 383 |
| Glycan biosynthesis and metabolism | 376 |
| Metabolism of cofactors and vitamins | 351 |
| Replication and repair | 299 |
| Environmental adaptation | 289 |
| Development and regeneration | 284 |
| Circulatory system | 249 |
| Nucleotide metabolism | 226 |
| Xenobiotics biodegradation and metabolism | 210 |
| Cell motility | 210 |
| Transcription | 181 |
| Sensory system | 164 |
| Ageing | 150 |
| Metabolism of other amino acids | 148 |
| Biosynthesis of other secondary metabolites | 125 |
| Excretory system | 121 |
| Metabolism of terpenoids and polyketides | 86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emami-Khoyi, A.; Le Roux, R.; Adair, M.G.; Monsanto, D.M.; Main, D.C.; Parbhu, S.P.; Schnelle, C.M.; van der Lingen, C.D.; Jansen van Vuuren, B.; Teske, P.R. Transcriptomic Diversity in the Livers of South African Sardines Participating in the Annual Sardine Run. Genes 2021, 12, 368. https://doi.org/10.3390/genes12030368
Emami-Khoyi A, Le Roux R, Adair MG, Monsanto DM, Main DC, Parbhu SP, Schnelle CM, van der Lingen CD, Jansen van Vuuren B, Teske PR. Transcriptomic Diversity in the Livers of South African Sardines Participating in the Annual Sardine Run. Genes. 2021; 12(3):368. https://doi.org/10.3390/genes12030368
Chicago/Turabian StyleEmami-Khoyi, Arsalan, Rynhardt Le Roux, Matthew G. Adair, Daniela M. Monsanto, Devon C. Main, Shilpa P. Parbhu, Claudia M. Schnelle, Carl D. van der Lingen, Bettine Jansen van Vuuren, and Peter R. Teske. 2021. "Transcriptomic Diversity in the Livers of South African Sardines Participating in the Annual Sardine Run" Genes 12, no. 3: 368. https://doi.org/10.3390/genes12030368
APA StyleEmami-Khoyi, A., Le Roux, R., Adair, M. G., Monsanto, D. M., Main, D. C., Parbhu, S. P., Schnelle, C. M., van der Lingen, C. D., Jansen van Vuuren, B., & Teske, P. R. (2021). Transcriptomic Diversity in the Livers of South African Sardines Participating in the Annual Sardine Run. Genes, 12(3), 368. https://doi.org/10.3390/genes12030368