
Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Standards
2.2. Animals
2.3. Collection and Processing of the Biological Material
2.4. Isolation and Culture of Rabbit Stem Cells
2.5. Culture of Human Adipose-Derived Stem Cells
2.6. Population Doubling Time
2.7. Detection of Surface and Intracellular Markers Using Flow Cytometry
2.8. Detection of Surface and Intracellular Markers Using Confocal Microscopy
2.9. RT-PCR
2.10. Digital Droplet PCR
2.11. Neurogenic Differentiation
2.12. RT-qPCR
2.13. Fluorescent Assessment of Neurodifferentiation
2.14. Statistical Analysis
3. Results
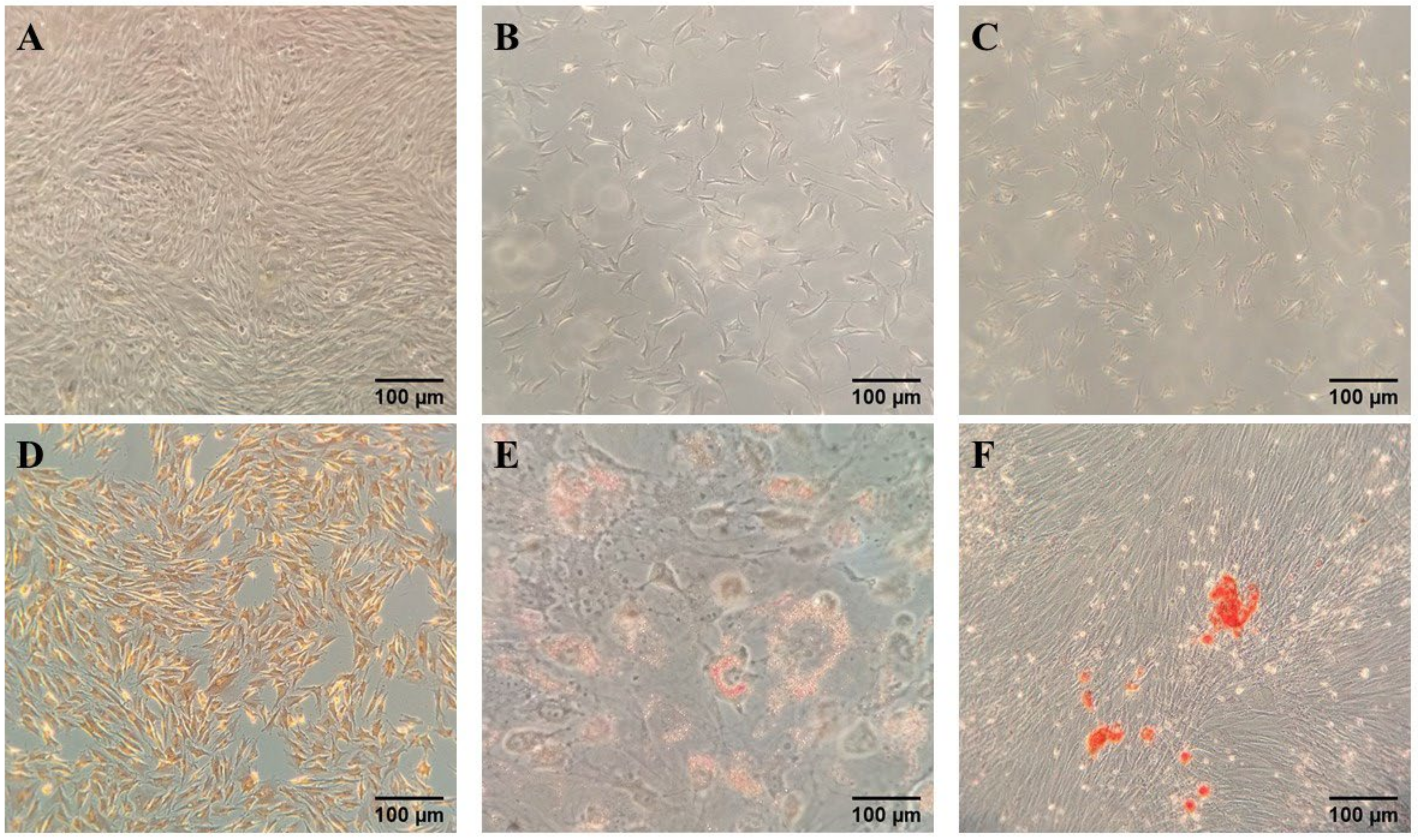
3.1. Morphology and Proliferation of Rabbit AT-MSCs
3.2. Detection of the Expression of Surface and Intracellular Markers Using Flow Cytometry
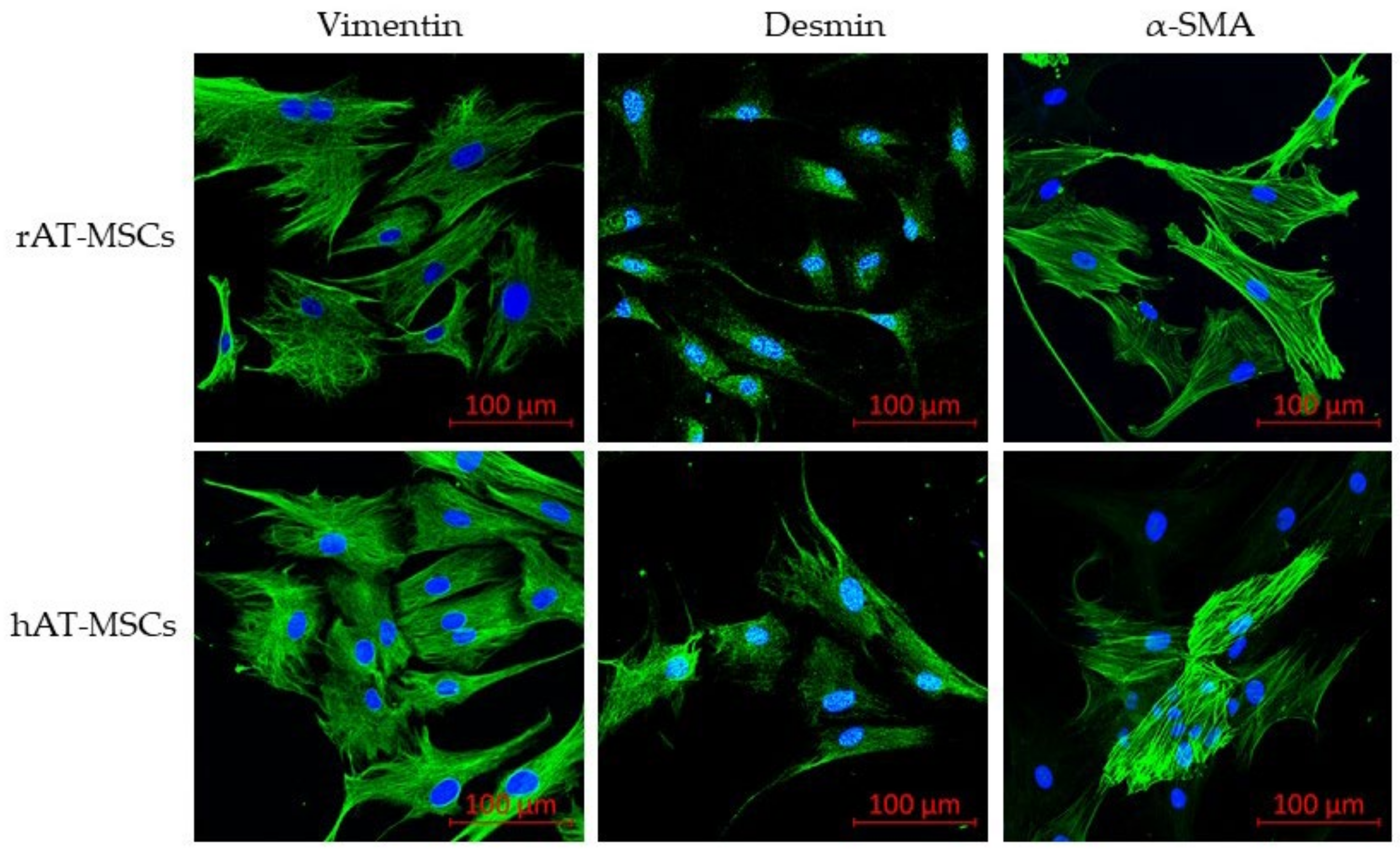
3.3. Detection of Surface and Intracellular Markers Using Confocal Microscopy
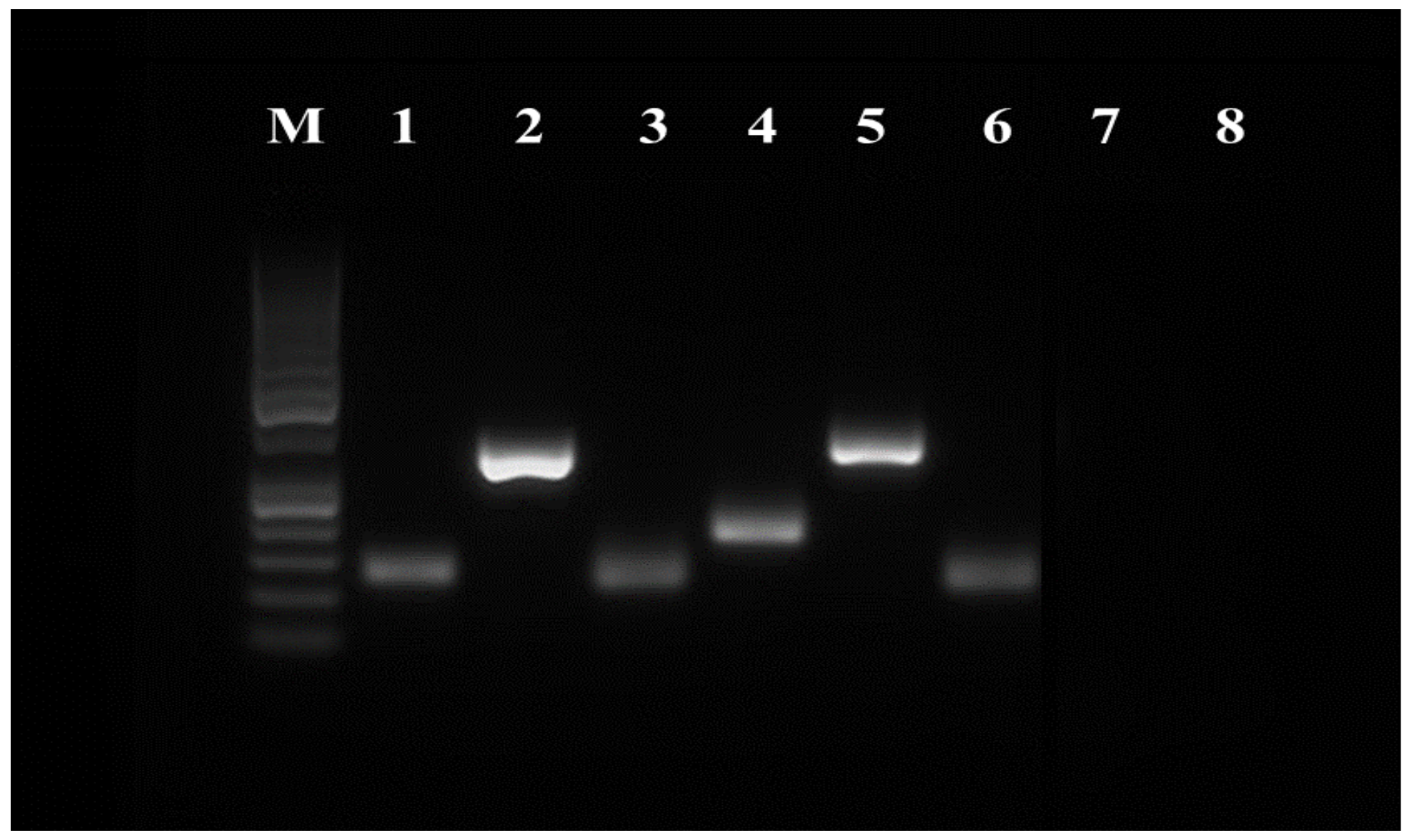
3.4. RT-PCR
3.5. Droplet Digital PCR
3.6. Neurodifferentiation of Rabbit Stem Cells
3.6.1. RT-qPCR
3.6.2. Confocal Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Differentiation Assays

References
- Seo, Y.; Shin, T.-H.; Kim, H.-S. Current Strategies to Enhance Adipose Stem Cell Function: An Update. Int. J. Mol. Sci. 2019, 20, 3827. [Google Scholar] [CrossRef] [Green Version]
- Krueger, T.E.G.; Thorek, D.L.J.; Denmeade, S.R.; Isaacs, J.T.; Brennen, W.N. Concise Review: Mesenchymal Stem Cell-Based Drug Delivery: The Good, the Bad, the Ugly, and the Promise. Stem Cells Transl. Med. 2018, 7, 651–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, T.; Lee, S.; Yoon, H.-Y.; Lee, J.I. Current Status of Canine Umbilical Cord Blood-Derived Mesenchymal Stem Cells in Veterinary Medicine. Stem Cells Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, D.T. Banking of Adipose- and Cord Tissue-Derived Stem Cells: Technical and Regulatory Issues. Adv. Exp. Med. Biol. 2016, 951, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.W.; Choi, J.R.; Wan Safwani, W.K.Z. Biobanking of human mesenchymal stem cells: Future strategy to facilitate clinical applications. In Biobanking and Cryopreservation of Stem Cells; Karimi-Busheri, F., Weinfeld, M., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2016; pp. 99–110. [Google Scholar] [CrossRef]
- Dayem, A.A.; Bin Lee, S.; Kim, K.; Lim, K.M.; Jeon, T.-I.; Seok, J.; Cho, A.S.-G. Production of Mesenchymal Stem Cells Through Stem Cell Reprogramming. Int. J. Mol. Sci. 2019, 20, 1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majka, M.; Sułkowski, M.; Badyra, B.; Musiałek, P. Concise Review: Mesenchymal Stem Cells in Cardiovascular Regeneration: Emerging Research Directions and Clinical Applications. Stem Cells Transl. Med. 2017, 6, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.O.; Gordon-Evans, W.J.; Fredericks, L.P.; Kiefer, K.; Conzemius, M.G.; Griffon, D.J. Comparison of Mesenchymal Stem Cell Surface Markers from Bone Marrow Aspirates and Adipose Stromal Vascular Fraction Sites. Front. Veter. Sci. 2016, 2, 82. [Google Scholar] [CrossRef] [Green Version]
- Strem, B.M.; Hicok, K.C.; Zhu, M.; Wulur, I.; Alfonso, Z.; E Schreiber, R.; Fraser, J.K.; Hedrick, M.H. Multipotential differentiation of adipose tissue-derived stem cells. Keio J. Med. 2005, 54, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, T.J.; Abrahamse, H. Neuronal Differentiation of Adipose Derived Stem Cells: Progress So Far. Int. J. Photoenergy 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-C.; Lee, T.-H.; Huang, Y.-H.; Chang, N.-K.; Lin, Y.-J.; Chien, P.-W.C.; Yang, W.-H.; Lin, M.H.-C. Comparison of Surface Markers between Human and Rabbit Mesenchymal Stem Cells. PLoS ONE 2014, 9, e111390. [Google Scholar] [CrossRef] [Green Version]
- Screven, R.; Kenyon, E.; Myers, M.J.; Yancy, H.F.; Skasko, M.; Boxer, L.; Bigley, E.C., III; Borjesson, D.L.; Zhu, M. Immunophenotype and Gene Expression Profile of Mesenchymal Stem Cells Derived from Canine Adipose Tissue and Bone Marrow. Vet. Immunol. Immunopathol. 2014, 161, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Kocan, B.; Maziarz, A.; Tabarkiewicz, J.; Ochiya, T.; Banaś-Ząbczyk, A. Trophic Activity and Phenotype of Adipose Tissue-Derived Mesenchymal Stem Cells as a Background of Their Regenerative Potential. Stem Cells Int. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- González-Garza, M.T.; Cruz-Vega, D.E.; Cárdenas-Lopez, A.; De La Rosa, R.M.; Moreno-Cuevas, J.E. Comparing stemness gene expression between stem cell subpopulations from peripheral blood and adipose tissue. Am. J. Stem Cells 2018, 7, 38–47. [Google Scholar] [PubMed]
- Bailey, A.M.; Kapur, S.; Katz, A.J. Characterization of Adipose-Derived Stem Cells: An Update. Curr. Stem Cell Res. Ther. 2010, 5, 95–102. [Google Scholar] [CrossRef]
- Mizuno, H.; Tobita, M.; Uysal, A.C. Concise Review: Adipose-Derived Stem Cells as a Novel Tool for Future Regenerative Medicine. Stem Cells 2012, 30, 804–810. [Google Scholar] [CrossRef]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Bajek, A.; Gurtowska, N.; Olkowska, J.; Maj, M.; Kaźmierski, Ł.; Bodnar, M.; Marszałek, A.; Dębski, R.; Drewa, T. Does the Harvesting Technique Affect the Properties of Adipose-Derived Stem Cells?-The Comparative Biological Characterization. J. Cell. Biochem. 2017, 118, 1097–1107. [Google Scholar] [CrossRef]
- Si, Z.; Wang, X.; Sun, C.; Kang, Y.; Xu, J.; Wang, X.; Hui, Y. Adipose-derived stem cells: Sources, potency, and implications for regenerative therapies. Biomed. Pharmacother. 2019, 114, 108765. [Google Scholar] [CrossRef]
- Anwer, A.G.; Gosnell, M.E.; Perinchery, S.M.; Inglis, D.W.; Goldys, E.M. Visible 532 nm laser irradiation of human adipose tissue-derived stem cells: Effect on proliferation rates, mitochondria membrane potential and autofluorescence. Lasers Surg. Med. 2012, 44, 769–778. [Google Scholar] [CrossRef]
- Mvula, B.; Moore, T.J.; Abrahamse, H. Effect of low-level laser irradiation and epidermal growth factor on adult human adipose-derived stem cells. Lasers Med. Sci. 2010, 25, 33–39. [Google Scholar] [CrossRef]
- De Villiers, J.A.; Houreld, N.N.; Abrahamse, H. Influence of Low Intensity Laser Irradiation on Isolated Human Adipose Derived Stem Cells Over 72 Hours and Their Differentiation Potential into Smooth Muscle Cells Using Retinoic Acid. Stem Cell Rev. Rep. 2011, 7, 869–882. [Google Scholar] [CrossRef]
- Muller, S.; Abrahamse, H.; Ndlovu, L.; Houreld, N.N. Fluence and Wavelength of Low Intensity Laser Irradiation Affect Activity and Proliferation of Human Adipose Derived Stem Cells: Peer Reviewed Original Article. Med. Technol. SA 2010, 24, 15–20. [Google Scholar]
- Neuhuber, B.; Gallo, G.; Howard, L.; Kostura, L.; Mackay, A.; Fischer, I. Reevaluation of in vitro differentiation protocols for bone marrow stromal cells: Disruption of actin cytoskeleton induces rapid morphological changes and mimics neuronal phenotype. J. Neurosci. Res. 2004, 77, 192–204. [Google Scholar] [CrossRef]
- Bertani, N.; Malatesta, P.; Volpi, G.; Sonego, P.; Perris, R. Neurogenic potential of human mesenchymal stem cells revisited: Analysis by immunostaining, time-lapse video and microarray. J. Cell Sci. 2005, 118, 3925–3936. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, G.; Lopatina, T.; Kalinina, N.; Rybalkina, E.; Parfyonova, Y.; Tkachuk, V.; Revishchin, A. In vitro neuronal induction of adipose-derived stem cells and their fate after transplantation into injured mouse brain. Curr. Med. Chem. 2012, 19, 5170–5177. [Google Scholar] [CrossRef]
- Cardozo, A.J.; Gómez, D.E.; Argibay, P.F. Neurogenic differentiation of human adipose-derived stem cells: Relevance of different signaling molecules, transcription factors, and key marker genes. Gene 2012, 511, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Intawicha, P.; Ou, Y.-W.; Lo, N.-W.; Zhang, S.-C.; Chen, Y.-Z.; Lin, T.-A.; Su, H.-L.; Guu, H.-F.; Chen, M.-J.; Lee, K.-H.; et al. Characterization of Embryonic Stem Cell Lines Derived from New Zealand White Rabbit Embryos. Cloning Stem Cells 2009, 11, 27–38. [Google Scholar] [CrossRef]
- Calasans-Maia, M.D.; Monteiro, M.L.; Áscoli, F.O.; Granjeiro, J.M. The rabbit as an animal model for experimental surgery. Acta Cir. Bras. 2009, 24, 325–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, B.S.; Bhat, K.M.; Mapara, M. Rabbit as an animal model for experimental research. Dent. Res. J. 2012, 9, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Kovac, M.; Vasicek, J.; Kulikova, B.; Bauer, M.; Curlej, J.; Balazi, A.; Chrenek, P. Different RNA and protein expression of surface markers in rabbit amniotic fluid-derived mesenchymal stem cells. Biotechnol. Prog. 2017, 33, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, B.; Kovac, M.; Bauer, M.; Tomkova, M.; Olexikova, L.; Vasicek, J.; Balazi, A.; Makarevich, A.V.; Chrenek, P. Survivability of rabbit amniotic fluid-derived mesenchymal stem cells post slow-freezing or vitrification. Acta Histochem. 2019, 121, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Vašíček, J.; Kováč, M.; Baláži, A.; Kulíková, B.; Tomková, M.; Olexíková, L.; Čurlej, J.; Bauer, M.; Schnabl, S.; Hilgarth, M.; et al. Combined approach for characterization and quality assessment of rabbit bone marrow-derived mesenchymal stem cells intended for gene banking. New Biotechnol. 2020, 54, 1–12. [Google Scholar] [CrossRef]
- Tomková, M.; Kulíková, B.; Vašíček, J.; Baláži, A.; Makarevič, A.; Chrenek, P. Effect of Different Culture Medium on Cultivation of Adipose Tissue Derived Stem Cells from Two Biological Sources. J. Microbiol. Biotechnol. Food Sci. 2018, 8, 798–801. [Google Scholar] [CrossRef]
- Su, J.; Chen, X.; Huang, Y.; Li, W.; Li, J.; Cao, K.; Cao, G.; Zhang, L.; Li, F.; I Roberts, A.; et al. Phylogenetic distinction of iNOS and IDO function in mesenchymal stem cell-mediated immunosuppression in mammalian species. Cell Death Differ. 2014, 21, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Esfandiari, B.; Soliemani, M.; Kaviani, S.; Parivar, K. Rapid Neural Differentiation of Human Adipose Tissue-Derived Stem Cells Using NGF, Forskolin and BFGF. Biomed. Pharmacol. J. 2016, 9, 39. [Google Scholar] [CrossRef]
- Gayathri, V.; Harikrishnan, V.; Mohanan, P.V. Integration of Rabbit Adipose Derived Mesenchymal Stem Cells to Hydroxyapatite Burr Hole Button Device for Bone Interface Regeneration. Int. J. Biomater. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zomer, H.D.; Roballo, K.C.; Lessa, T.B.; Bressan, F.F.; Gonçalves, N.N.; Meirelles, F.V.; Trentin, A.G.; Ambrósio, C.E. Distinct features of rabbit and human adipose-derived mesenchymal stem cells: Implications for biotechnology and translational research. Stem Cells Cloning Adv. Appl. 2018, 11, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.-S.; El-Ashram, S.; Luo, D.-Z.; Luo, H.-N.; Wang, B.-Y.; Chen, S.-F.; Bai, Y.-S.; Chen, Z.-S.; Liu, C.-Y.; Ji, H.-Q. A Comparative Study of Biological Characteristics and Transcriptome Profiles of Mesenchymal Stem Cells from Different Canine Tissues. Int. J. Mol. Sci. 2019, 20, 1485. [Google Scholar] [CrossRef] [Green Version]
- Arrizabalaga, J.H.; Nollert, M.U. Properties of porcine adipose-derived stem cells and their applications in preclinical models. Adipocyte 2017, 6, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Bourebaba, L.; Michalak, I.; Baouche, M.; Kucharczyk, K.; Marycz, K. Cladophora glomerata methanolic extract promotes chondrogenic gene expression and cartilage phenotype differentiation in equine adipose-derived mesenchymal stromal stem cells affected by metabolic syndrome. Stem Cell Res. Ther. 2019, 10, 1–20. [Google Scholar] [CrossRef]
- Elashry, M.I.; Gegnaw, S.T.; Klymiuk, M.C.; Wenisch, S.; Arnhold, S. Influence of mechanical fluid shear stress on the osteogenic differentiation protocols for Equine adipose tissue-derived mesenchymal stem cells. Acta Histochem. 2019, 121, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, P.; Lombardi, F.; Siragusa, G.; Cifone, M.G.; Cinque, B.; Giuliani, M. Methods of Isolation, Characterization and Expansion of Human Adipose-Derived Stem Cells (ASCs): An Overview. Int. J. Mol. Sci. 2018, 19, 1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.K.; Homrich, S.G.; Rodrigues, C.C.R.; Azzolin, V.F.; Duarte, M.M.M.F.; Pillar, D.M.; Lenz, L.S.; Unfer, T.C.; De Souza, D.V.; Da Cruz, I.B.M. Human adipose-derived stem cells obtained from lipoaspirates are highly susceptible to hydrogen peroxide mediated cytogenotoxicity. Arch. Biosci. Health 2019, 1, 11–28. [Google Scholar] [CrossRef]
- Kawagishi-Hotta, M.; Hasegawa, S.; Igarashi, T.; Yamada, T.; Takahashi, M.; Numata, S.; Kobayashi, T.; Iwata, Y.; Arima, M.; Yamamoto, N.; et al. Enhancement of individual differences in proliferation and differentiation potentials of aged human adipose-derived stem cells. Regen. Ther. 2017, 6, 29–40. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Umbilical Cord Blood, or Adipose Tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Baghban, E.M.R.; Mardpour, S.; Ebrahimi, M. Growth Kinetics and in Vitro Aging of Mesenchymal Stem Cells Isolated from Rat Adipose versus Bone Marrow Tissues. Iran. J. Vet. Surg. 2008, 3, 9–20. [Google Scholar]
- Aliborzi, G.; Vahdati, A.; Mehrabani, D.; Hosseini, S.E.; Tamadon, A. Isolation, Characterization and Growth Kinetic Comparison of Bone Marrow and Adipose Tissue Mesenchymal Stem Cells of Guinea Pig. Int. J. Stem Cells 2016, 9, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lorenzo, M.J.; Royo-Cañas, M.; Alegre-Aguarón, E.; Desportes, P.; Castiella, T.; García-Álvarez, F.; Larrad, L. Phenotype and Chondrogenic Differentiation of Mesenchymal Cells from Adipose Tissue of Different Species. J. Orthop. Res. 2009, 27, 1499–1507. [Google Scholar] [CrossRef]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise Review: The Surface Markers and Identity of Human Mesenchymal Stem Cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Seo, M.J.; Pulin, A.A.; Gregory, C.A.; Ylostalo, J.; Prockop, D.J. The CD34-like protein PODXL and α6-integrin (CD49f) identify early progenitor MSCs with increased clonogenicity and migration to infarcted heart in mice. Blood 2009, 113, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.-R.; Yang, S.-R.; Jung, J.-W.; Kim, H.; Ko, K.; Han, D.W.; Park, S.-B.; Choi, S.W.; Kang, S.-K.; Scholer, H.; et al. CD49f Enhances Multipotency and Maintains Stemness Through the Direct Regulation of OCT4 and SOX2. Stem Cells 2012, 30, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, G. Aldehyde Dehydrogenases: Not Just Markers, but Functional Regulators of Stem Cells. Stem Cells Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Nishikawa, S.; Haraguchi, T.; Arikawa, Y.; Eto, S.; Hiyama, M.; Iseri, T.; Itoh, Y.; Nakaichi, M.; Sakai, Y.; et al. Aldehyde dehydrogenase activity helps identify a subpopulation of murine adipose-derived stem cells with enhanced adipogenic and osteogenic differentiation potential. World J. Stem Cells 2017, 9, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Tsekouras, A.; Mantas, D.; Tsilimigras, I.D.; Moris, D.; Kontos, M.; Zografos, C.G. Comparison of the Viability and Yield of Adipose-Derived Stem Cells (ASCs) from Different Donor Areas. In Vivo 2017, 31, 1229–1234. [Google Scholar] [CrossRef] [Green Version]
- Zampar, A.G.; Junior, J.A.F.; Orellana, M.D.; Caruso, S.R.; Fernandes, T.R.; Gomes, R.; Aragon, D.C.; De Santis, G.C.; Covas, D.T. Analysis of Adipose-Derived Stem Cells from Different Donor Areas and Their Influence on Fibroblasts In Vitro. Aesthetic Plast. Surg. 2020, 44, 971–978. [Google Scholar] [CrossRef]
- Mitterberger, M.C.; Lechner, S.; Mattesich, M.; Kaiser, A.; Probst, D.; Wenger, N.; Pierer, G.; Zwerschke, W. DLK1 (PREF1) Is a Negative Regulator of Adipogenesis in CD105+/CD90+/CD34+/CD31−/FABP4− Adipose-Derived Stromal Cells from Subcutaneous Abdominal Fat Pats of Adult Women. Stem Cell Res. 2012, 9, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Vašíček, J.; Shehata, M.; Schnabl, S.; Hilgarth, M.; Hubmann, R.; Jäger, U.; Bauer, M.; Chrenek, P. Critical assessment of the efficiency of CD34 and CD133 antibodies for enrichment of rabbit hematopoietic stem cells. Biotechnol. Prog. 2018, 34, 1278–1289. [Google Scholar] [CrossRef]
- Yaszemski, A.; Rooney, G.E.; Windebank, A.J. Characterization of Rabbit Mesenchymal Stem Cells. In Experimental Biology 2011 Meeting Abstracts; Wiley: Washington, MD, USA, 2011. [Google Scholar]
- Echeverry, D.M.; Rojas, D.M.; Aguilera, C.J.; Veraguas, D.M.; Cabezas, J.G.; Rodríguez-Álvarez, L.; Castro, F.O. Differentiation and multipotential characteristics of mesenchymal stem cells derived from adipose tissue of an endangered wild cat (Leopardus guigna). Austral. J. Veter. Sci. 2019, 51, 17–26. [Google Scholar] [CrossRef]
- Peroni, D.; Scambi, I.; Pasini, A.; Lisi, V.; Bifari, F.; Krampera, M.; Rigotti, G.; Sbarbati, A.; Galiè, M. Stem molecular signature of adipose-derived stromal cells. Exp. Cell Res. 2008, 314, 603–615. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of Molecular Profiles of Human Mesenchymal Stem Cells Derived from Bone Marrow, Umbilical Cord Blood, Placenta and Adipose Tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, J.; Horvath, T.L.; Ballas, C.B.; March, K.L.; Srour, E.F. In vitro clonal analysis of murine pluripotent stem cells isolated from skeletal muscle and adipose stromal cells. Exp. Hematol. 2008, 36, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almalki, S.G.; Agrawal, D.K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, S.; Trentz, O.A.; Reddy, M.S.; Rela, M.; Kandasamy, M.; Sellathamby, S. In vitro transdifferentiation of human adipose tissue-derived stem cells to neural lineage cells—A stage-specific incidence. Adipocyte 2019, 8, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Hasani, S.; Javeri, A.; Asadi, A.; Taha, M.F. Cardiac Differentiation of Adipose Tissue-Derived Stem Cells Is Driven by BMP4 and bFGF but Counteracted by 5-Azacytidine and Valproic Acid. Cell J. 2019, 22, 273–282. [Google Scholar]
- Al-Ghadban, S.; Bunnell, B.A. Adipose Tissue-Derived Stem Cells: Immunomodulatory Effects and Therapeutic Potential. Physiology 2020, 35, 125–133. [Google Scholar] [CrossRef]
- Lu, T.; Pei, W.; Wang, K.; Zhang, S.; Chen, F.; Wu, Y.; Guan, W. In Vitro Culture and Biological Properties of Broiler Adipose-derived Stem Cells. Exp. Ther. Med. 2018, 16, 2399–2407. [Google Scholar] [CrossRef] [Green Version]
- Kruminis-Kaszkiel, E.; Osowski, A.; Bejer-Oleńska, E.; Dziekoński, M.; Wojtkiewicz, J. Differentiation of Human Mesenchymal Stem Cells from Wharton’s Jelly Towards Neural Stem Cells Using a Feasible and Repeatable Protocol. Cells 2020, 9, 739. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Medina, L.V.; Pasantes-Morales, H.; Aguilera-Castrejon, A.; Picones, A.; Lara-Figueroa, C.O.; Luis, E.; Montesinos, J.J.; Cortés-Morales, V.A.; Ruiz, M.P.D.L.R.; Hernández-Estévez, E.; et al. Neuronal Transdifferentiation Potential of Human Mesenchymal Stem Cells from Neonatal and Adult Sources by a Small Molecule Cocktail. Stem Cells Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Urrutia, D.N.; Caviedes, P.; Mardones, R.; Minguell, J.J.; Vega-Letter, A.M.; Jofre, C.M. Comparative study of the neural differentiation capacity of mesenchymal stromal cells from different tissue sources: An approach for their use in neural regeneration therapies. PLoS ONE 2019, 14, e0213032. [Google Scholar] [CrossRef]
- Kozlowska, U.; Krawczenko, A.; Futoma, K.; Jurek, T.; Rorat, M.; Patrzalek, D.; Klimczak, A. Similarities and differences between mesenchymal stem/progenitor cells derived from various human tissues. World J. Stem Cells 2019, 11, 347–374. [Google Scholar] [CrossRef] [PubMed]
- Musina, R.A.; Bekchanova, E.S.; Sukhikh, G.T. Comparison of Mesenchymal Stem Cells Obtained from Different Human Tissues. Bull. Exp. Biol. Med. 2005, 139, 504–509. [Google Scholar] [CrossRef]
- Forghani, A.; Koduru, S.V.; Chen, C.; Leberfinger, A.N.; Ravnic, D.J.; Hayes, D.J. Differentiation of Adipose Tissue–Derived CD34+/CD31− Cells into Endothelial Cells In Vitro. Regen. Eng. Transl. Med. 2020, 6, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Baptista, L.S. Adipose stromal/stem cells in regenerative medicine: Potentials and limitations. World J. Stem Cells 2020, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lisini, D.; Nava, S.; Pogliani, S.; Avanzini, M.; Lenta, E.; Bedini, G.; Mantelli, M.; Pecciarini, L.; Croce, S.; Boncoraglio, G.; et al. Adipose tissue-derived mesenchymal stromal cells for clinical application: An efficient isolation approach. Curr. Res. Transl. Med. 2019, 67, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Adolfsson, E.; Helenius, G.; Friberg, Ö.; Samano, N.; Frøbert, O.; Johansson, K. Bone marrow- and adipose tissue-derived mesenchymal stem cells from donors with coronary artery disease; growth, yield, gene expression and the effect of oxygen concentration. Scand. J. Clin. Lab. Investig. 2020, 80, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Patrikoski, M.; Mannerström, B.; Miettinen, S. Perspectives for Clinical Translation of Adipose Stromal/Stem Cells. Stem Cells Int. 2019, 2019, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.-L.; Ahmad, T.S.; Selvaratnam, L.; Kamarul, T. Isolation, characterization and the multi-lineage differentiation potential of rabbit bone marrow-derived mesenchymal stem cells. J. Anat. 2013, 222, 437–450. [Google Scholar] [CrossRef]
- Cong, Z.; Lin, J.-B.; Jiang, H.-X.; Lin, W.-B.; Gao, J.-T.; Gao, M.-M.; Wu, B.-W.; Feng, B.; Cheng, N.-N.; Huang, G.; et al. Comparison and Optimization: Different Medium and a Novel Scheme for Rabbit Bone Marrow Mesenchymal Stem Cells Culture. Res. Square 2020. [Google Scholar] [CrossRef]
- Borghesi, J.; Mario, L.C.; Carreira, A.C.O.; Miglino, M.A.; Favaron, P.O. Phenotype and multipotency of rabbit (Oryctolagus cuniculus) amniotic stem cells. Stem Cell Res. Ther. 2017, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Kováč, M.; Kulíková, B.; Vašíček, J.; Chrenek, P. Cryopreservation of Amniotic Fluid Stem Cells Derived from Zobor Rabbit. Slovak J. Anim. Sci. 2016, 2016, 62–67. [Google Scholar]
- Slamečka, J.J.; Chrenek, P. Rabbit Amniotic Fluid as a Potential Alternative Source of Broadly Multipotent Stem Cells. Slovak J. Anim. Sci. 2013, 2013, 11–15. [Google Scholar]
- Jiang, S.; Zhang, S. Differentiation of cardiomyocytes from amniotic fluid-derived mesenchymal stem cells by combined induction with transforming growth factor β1 and 5-azacytidine. Mol. Med. Rep. 2017, 16, 5887–5893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, T.; Chelluri, L.K. Standardization and quality assessment for clinical grade mesenchymal stem cells from human adipose tissue. Hematol. Transfus. Cell Ther. 2019, 41, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bajek, A.; Olkowska, J.; Walentowicz-Sadłecka, M.; Sadłecki, P.; Grabiec, M.; Porowinska, D.; Drewa, T.; Roszkowski, K. Human Adipose-Derived and Amniotic Fluid-Derived Stem Cells: A Preliminary In Vitro Study Comparing Myogenic Differentiation Capability. Med. Sci. Monit. 2018, 24, 1733–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Dekoninck, P.; Toelen, J.; Roubliova, X.; Carter, S.; Pozzobon, M.; Russo, F.M.; Richter, J.; Vandersloten, P.-J.; Verbeken, E.; De Coppi, P.; et al. The use of human amniotic fluid stem cells as an adjunct to promote pulmonary development in a rabbit model for congenital diaphragmatic hernia. Prenat. Diagn. 2015, 35, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Roubelakis, M.G.; Pappa, K.I.; Bitsika, V.; Zagoura, D.; Vlahou, A.; Papadaki, H.A.; Antsaklis, A.; Anagnou, N.P. Molecular and Proteomic Characterization of Human Mesenchymal Stem Cells Derived from Amniotic Fluid: Comparison to Bone Marrow Mesenchymal Stem Cells. Stem Cells Dev. 2007, 16, 931–952. [Google Scholar] [CrossRef] [Green Version]
- Fei, X.; Jiang, S.; Zhang, S.; Li, Y.; Ge, J.; He, B.; Goldstein, S.; Ruiz, G. Isolation, Culture, and Identification of Amniotic Fluid-Derived Mesenchymal Stem Cells. Cell Biophys. 2013, 67, 689–694. [Google Scholar] [CrossRef]
- Janz, F.D.L.; Debes, A.D.A.; Cavaglieri, R.D.C.; Duarte, S.A.; Romão, C.M.; Morón, A.F.; Zugaib, M.; Bydlowski, S.P. Evaluation of Distinct Freezing Methods and Cryoprotectants for Human Amniotic Fluid Stem Cells Cryopreservation. J. Biomed. Biotechnol. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Perin, L.; Giuliani, S.; Jin, D.; Sedrakyan, S.; Carraro, G.; Habibian, R.; Warburton, D.; Atala, A.; De Filippo, R.E. Renal differentiation of amniotic fluid stem cells. Cell Prolif. 2007, 40, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh-Ghalehaziz, S.; Farahzadi, R.; Fathi, E.; Pashaiasl, A.M. A Mini Overview of Isolation, Characterization and Application of Amniotic Fluid Stem Cells. Int. J. Stem Cells 2015, 8, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, Y.; Kim, H.; Hwang, K.J.; Kwon, H.C.; Kim, S.K.; Cho, D.J.; Kang, S.G.; You, J. Human amniotic fluid-derived stem cells have characteristics of multipotent stem cells. Cell Prolif. 2007, 40, 75–90. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Host | Reactivity | Clone | Conjugate | Company |
|---|---|---|---|---|---|
| CD29 | mouse IgG1 | rabbit | P4G11 | FITC | Merck |
| CD34 | mouse IgG1 | human | QBEnd-10 | FITC | Thermo Fisher Scientific |
| CD44 | mouse IgG1 | rabbit | W4/86 | - | Bio-Rad |
| CD45 | mouse IgG1 | rabbit | L12/201 | - | Bio-Rad |
| CD49f | rat IgG2a | rabbit | GoH3 | AF647 | Biolegend |
| CD73 | rat IgG1 | mouse | TY/11.8 | PE-Cy7 | eBioscience |
| CD73 * | mouse IgG1 | human | AD2 | FITC | eBioscience |
| CD90 | mouse IgG1 | rat | OX-7 | PE-Cy7 | BD Biosciences |
| CD90 * | mouse IgG1 | human | 5E10 | FITC | BD Biosciences |
| CD105 | mouse IgG1 | rabbit | SN6 | FITC | GeneTex |
| CD105 * | mouse IgG1 | human | 266 | FITC | BD Biosciences |
| Vimentin | mouse IgG2a | human | Vim 3B4 | - | Dako Cytomation |
| α-SMA | mouse IgG2a | human | 1A4 | - | Dako Cytomation |
| Desmin | mouse IgG1 | human | D33 | - | Dako Cytomation |
| Marker | Host | Reactivity | Clone | Conjugate | Company |
|---|---|---|---|---|---|
| CD49f | rat IgG1 | rabbit | GoH3 | - | Biolegend |
| Sec. Ab | goat | rat | polyclonal | FITC | Biolegend |
| ALPL | mouse | rabbit | TRA-2-49 | - | Novus Biologicals |
| SOX2 | mouse | human | 245610 | - | R&D Systems |
| Sec. Ab | goat | mouse IgG | polyclonal | FITC | Bio-Rad |
| NANOG | goat | human | polyclonal | - | R&D Systems |
| OCT4 | goat | human | polyclonal | - | R&D Systems |
| Sec. Ab | donkey | goat IgG | polyclonal | FITC | Bio-Rad |
| Gene | Product Size (bp) | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|---|
| CD29 | 287 | 5′-AGAATGTCACCAACCGTAGCA-3′ | 5′-CACAAAGGAGCCAAACCCA-3′ | [35] |
| CD44 | 112 | 5′-TCATCCTGGCATCCCTCTTG-3′ | 5′-CCGTTGCCATTGTTGATCAC-3′ | [11] |
| CD73 | 170 | 5′-CTCCTTTCCTCTCAAATCCAG-3′ | 5′-GTCCACGCCCTTCACTTTC-3′ | [35] |
| CD90 | 293 | 5′-CTGCTGCTGCTCTCACTGTC-3′ | 5′-ACAGAAGCAGCTTTGGGAAA-3′ | [31] |
| CD105 | 109 | 5′-TGACATACAGCACCAGCCAG-3′ | 5′-AGCTCTGACACCTCGTTTGG-3′ | [11] |
| B2M | 118 | 5′-ATTCACGCCCAATGATAAGG-3′ | 5′-ATCCTCAGACCTCCATGCTG-3′ | [31] |
| CD34 | 206 | 5′-TTTCCTCATGAACCGTCGCA-3′ | 5′-CGTGTTGTCTTGCGGAATGG-3′ | [31] |
| CD45 | 262 | 5′-TACTCTGCCTCCCGTTG-3′ | 5′-GCTGAGTGTCTGCGTGTC-3′ | [35] |
| ST3GAL2 (SSEA-4) | 126 | 5′-CTGGGAGAATAACCGGTACG-3′ | 5′-GCTCAGTTGCCTCGGTAGAC-3′ | [33] |
| ALPL (MSCA-1) | 137 | 5′-CCCTCATGTGATGGCTTACG-3′ | 5′-CTCAGAACAGGACGCTCAGG-3′ | [33] |
| NANOG | 122 | 5′-GCCAGTCGTGGAGTAACCAT-3′ | 5′-CTGCATGGAGGACTGTAGCA-3′ | [31] |
| OCT4 | 149 | 5′-GAGGAGTCCCAGGACATGAA-3′ | 5′-GTGGTTTGGCTGAACACCTT-3′ | [31] |
| SOX2 | 152 | 5′-CAGCTCGCAGACCTACATGA-3′ | 5′-TGGAGTGGGAGGAAGAGGTA-3′ | [31] |
| ALDH | 135 | 5′-CTGGGAAAAGCAACCTGAAG-3′ | 5′-AACACTGGCCCTGATGGTAG-3′ | NM_001082013.1 1 |
| Gene | Product Size (bp) | Forward Primer | Reverse Primer | Reference | |
|---|---|---|---|---|---|
| rMSCs | ENO2 | 128 | 5′- ACACACTCAAGGGGGTCATC -3′ | 5′- GTCGATGGCTTCCTTTACCA -3′ | XM_002712914.3 1 |
| MAP2 | 161 | 5′- CTCACCATGTTCCTGGAGGT -3′ | 5′- GGAGGAGACGTTGCTGAGTC -3′ | XM_017343068.1 1 | |
| B2M | 118 | 5′-CTCCTTTCCTCTCAAATCCAG-3′ | 5′-GTCCACGCCCTTCACTTTC-3′ | [31] | |
| hMSCs | hENO2 | 238 | 5′- GGAGAACAGTGAAGCCTTGG -3′ | 5′- GGTCAAATGGGTCCTCAATG -3′ | [37] |
| hMAP2 | 97 | 5′- AGTTCCAGCAGCGTGATG -3′ | 5′- CATTCTCTCTTCAGCCTTCTC -3′ | [37] | |
| hACT | 125 | 5′- CCTGGCGTCGTCATTAGTG -3′ | 5′-TCAGTCCTGTCCATAATTAGTCC-3′ | [37] |
| Percentage of Positive Cells % | ||||
|---|---|---|---|---|
| rAT-MSCs | rBM-MSCs | rAF-MSCs | hAT-MSCs | |
| CD29 | 92.12 ± 6.65 | 89.50 ± 8.03 [33] | 96.0 ± 5.7 [32] | 98.98 ± 0.59 |
| CD34 | 1.42 ± 0.67 | 0.78 ± 0.44 [33] | 0.37 ± 0.2 [32] | 0.77 |
| CD44 | 97.15 ± 1.45 | 89.08 ± 8.44 [33] | 93.7 ± 2.3 [32] | NT |
| CD45 | 1.13 ± 0.40 | 4.52 ± 2.99 [33] | 1.65 ± 1.1 [32] | NT |
| CD49f | 98.92 ± 0.86 | 79.32 ± 12.63 | 96.68 ± 1.61 | 77.90 ± 10.18 |
| CD73 | 4.10 ± 0.02 | 3.54 ± 1.83 [33] | 7.93 ± 5.0 [32] | NT |
| CD73 * | 60.50 ± 7.37 | 73.75 ± 21.16 | 27.23 ± 16.54 | 98.58 ± 0.32 |
| CD90 | 10.21 ± 0.07 | 8.74 ± 4.39 [33] | 15.6 ± 4.0 [32] | NT |
| CD90 * | 95.97 ± 3.17 | 98.23 ± 2.09 | 70.77 ± 16.55 | 98.97 ± 0.47 |
| CD105 | 4.60 ± 0.86 | 2.03 ± 1.73 [33] | 0.56 ± 0.4 [32] | NT |
| CD105 * | 38.83 ± 4.30 | 73.45 ± 10.96 | 11.50 ± 3.30 | 94.44 ± 1.93 |
| Vimentin | 85.08 ± 11.43 | 97.84 ± 3.80 [33] | 91.9 ± 4.7 [32] | NT |
| α-SMA | 86.26 ± 10.15 | 98.75 ± 1.19 [33] | 89.0 ± 9.0 [32] | NT |
| Desmin | 75.29 ± 17.09 | 50.12 ± 11.37 [33] | 85.1 ± 9.9 [32] | NT |
| ALDH | 74.63 ± 13.61 | 70.60 ± 21.38 | 31.13 ± 8.11 | 75.56 ± 5.32 |
| Percentage of Positive Droplets % | |||
|---|---|---|---|
| rAT-MSCs | rBM-MSCs | rAF-MSCs | |
| CD29 | 90.3 ± 6.7 | 94.6 ± 5.4 | 89.1 ± 7.7 |
| CD44 | 99.5 ± 0.8 | 89.9 ± 8.7 | 89.6 ± 12.6 |
| CD45 | 0.0 ± 0.0 | 0.1 ± 0.0 | 0.0 ± 0.0 |
| CD73 | 60.7 ± 25.5 | 42.1 ± 15.4 | 25.7 ± 16.8 |
| CD90 | 99.9 ± 0.0 | 47.9 ± 8.6 | 58.3 ± 7.4 |
| CD105 | 55.8 ± 22.2 | 50.4 ± 28.0 | 16.1 ± 10.5 |
| Adipose Tissue | Bone Marrow | Amniotic Fluid | |
|---|---|---|---|
| rMSCs | PDT: approx. 2 days - Phenotype: CD29+; CD44+; CD73±; CD90±; CD105±; CD34±; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipo-genic, osteogenic, chondrogenic, neurogenic [17,34,39,58,60,75,76,77,78,79] | - PDT: approx. 5 days - Phenotype: CD29+; CD44+; CD73+; CD90+; CD105+; CD14-; CD34¬-; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipogenic, osteogenic, chondrogenic, neurogenic [31,33,60,80,81] | - PDT: approx. 3 days - Phenotype: CD29+; CD44+; CD73±; CD90±; CD105±; CD34 ¬-; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipogenic, osteogenic, chondrogenic, neurogenic, cardiomyocytes [31,32,33,82,83,84,85] |
| hMSCs | - PDT: approx. 2 days - Phenotype: CD29+; CD44+; CD73+; CD90+; CD105+; CD14-; CD31-; CD34¬-; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipo-genic, osteogenic, chondrogenic, neurogenic, cardiomyocytes, endothelial cells [39,47,63,66,67,68,72,86,87] | - PDT: approx. 6 days - Phenotype: CD29+; CD44+; CD73+; CD90+; CD105+; CD14-; CD34¬-; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipogenic, osteogenic, chondrogenic, neurogenic [47,63,72,80,88] | - PDT: approx. 2 days - Phenotype: CD29+; CD44+; CD73+; CD90+; CD105+; CD14-; CD34 ¬-; CD45- - SOX2±; OCT4±; NANOG± - Differentiation potential: adipogenic, osteogenic, chondrogenic, neurogenic, hepatocyte, epithelilal lung lineages, kidney lineage [40,87,89,90,91,92,93,94,95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirpáková, M.; Vašíček, J.; Svoradová, A.; Baláži, A.; Tomka, M.; Bauer, M.; Makarevich, A.; Chrenek, P. Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells. Genes 2021, 12, 431. https://doi.org/10.3390/genes12030431
Tirpáková M, Vašíček J, Svoradová A, Baláži A, Tomka M, Bauer M, Makarevich A, Chrenek P. Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells. Genes. 2021; 12(3):431. https://doi.org/10.3390/genes12030431
Chicago/Turabian StyleTirpáková, Mária, Jaromír Vašíček, Andrea Svoradová, Andrej Baláži, Marián Tomka, Miroslav Bauer, Alexander Makarevich, and Peter Chrenek. 2021. "Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells" Genes 12, no. 3: 431. https://doi.org/10.3390/genes12030431
APA StyleTirpáková, M., Vašíček, J., Svoradová, A., Baláži, A., Tomka, M., Bauer, M., Makarevich, A., & Chrenek, P. (2021). Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells. Genes, 12(3), 431. https://doi.org/10.3390/genes12030431