
Insect Behavioral Change and the Potential Contributions of Neuroinflammation—A Call for Future Research


Abstract
:1. Introduction
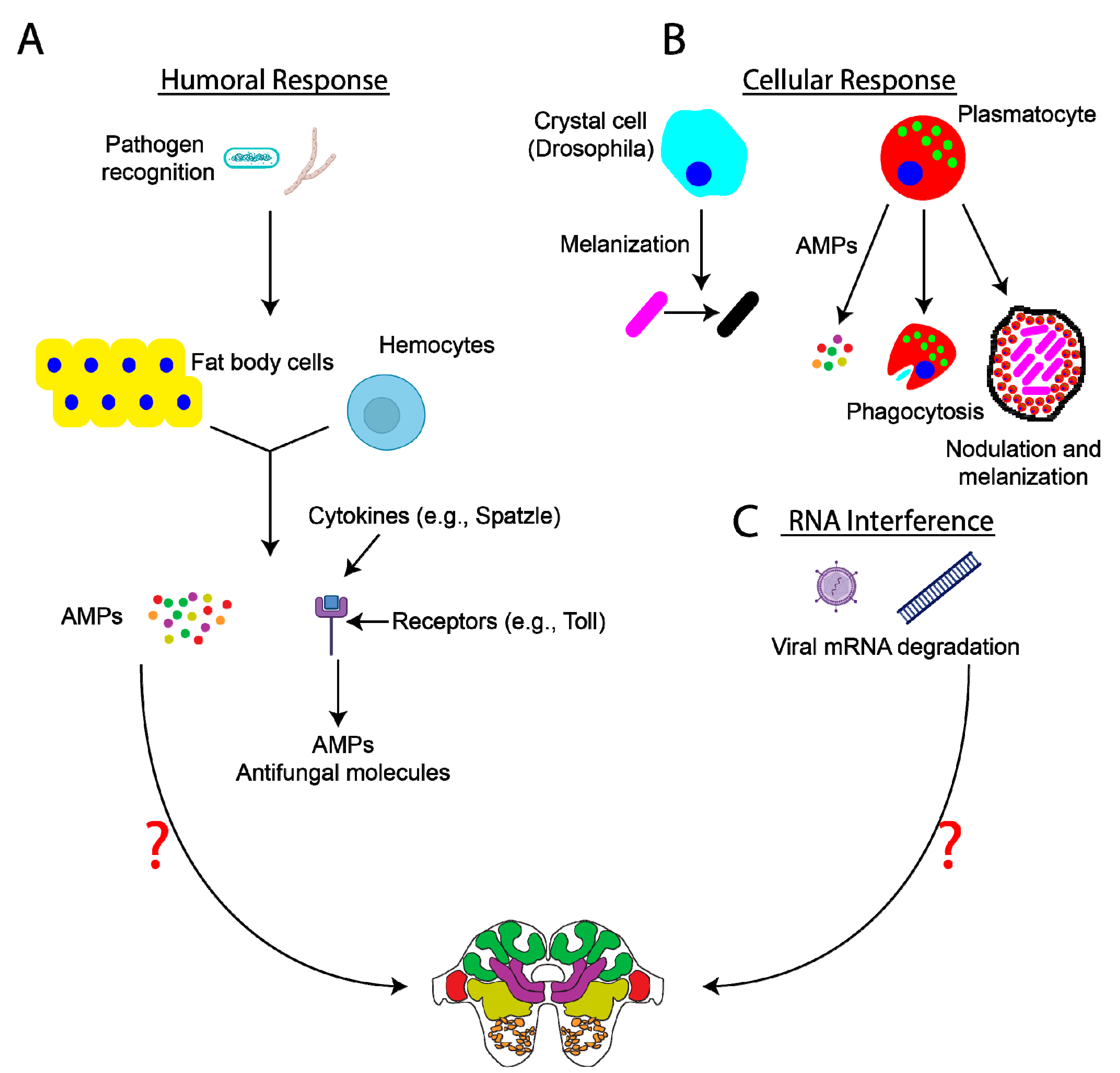
2. Insect Immunity—An Overview

3. Host Behavioral Change Associated with Microbial Infection
3.1. Viruses
3.2. Bacteria
3.3. Fungi
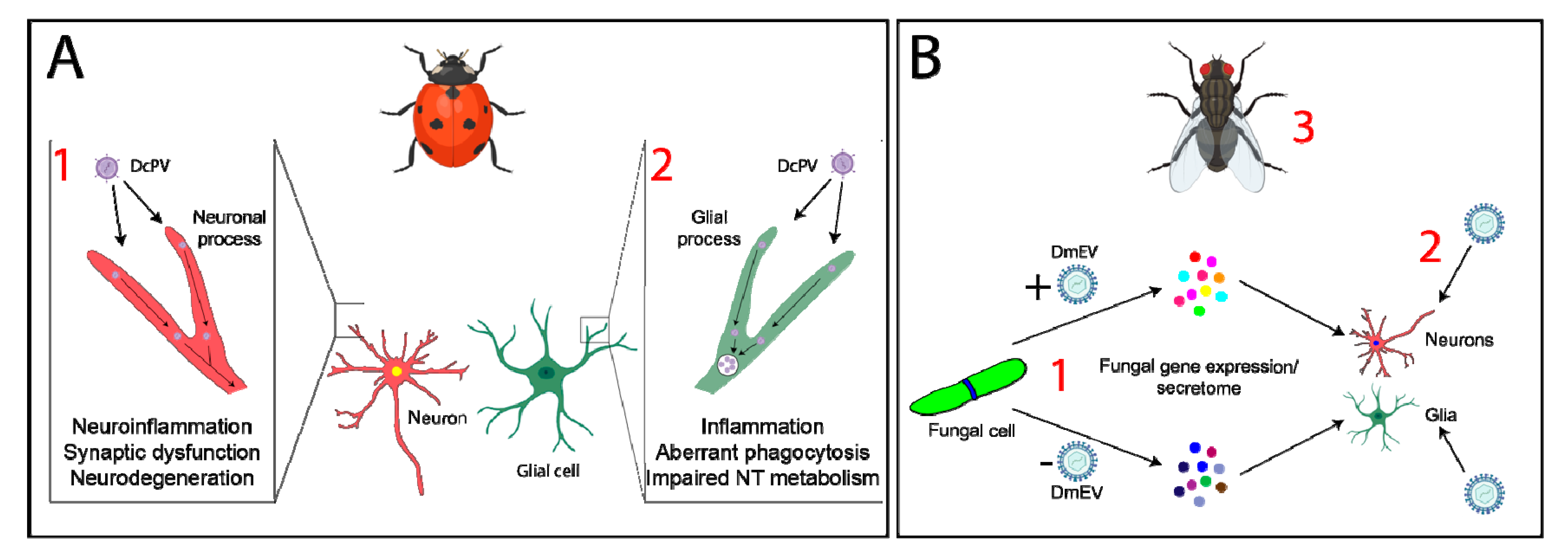
4. Parasitoids and Parasites—Neuromodulation, Molecular Mimicry, and Neuroinflammation
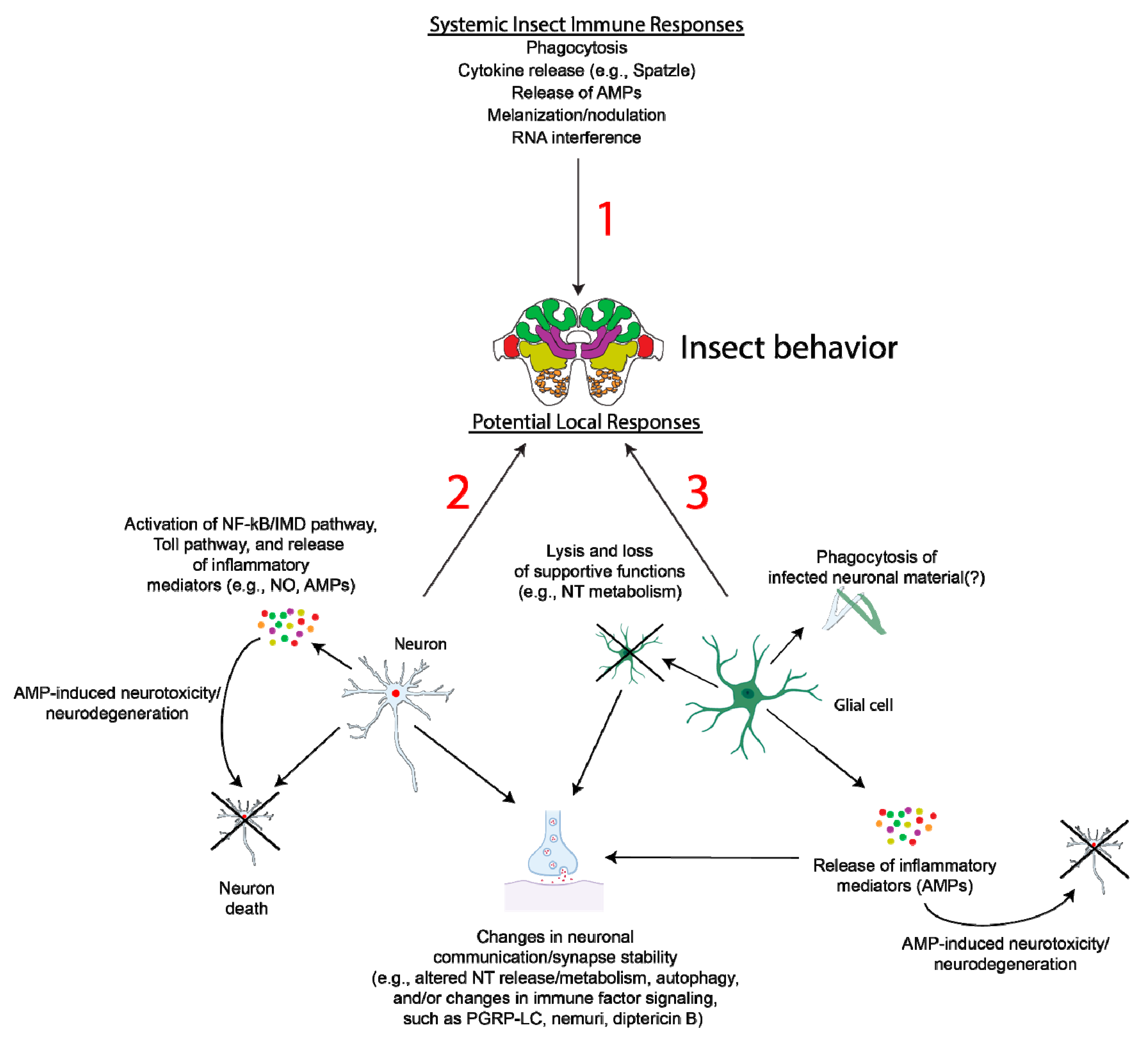
5. Neuroinflammation and Glia

6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Bekker, C.; Ohm, R.A.; Evans, H.C.; Brachmann, A.; Hughes, D.P. Ant-Infecting Ophiocordyceps Genomes Reveal a High Diversity of Potential Behavioral Manipulation Genes and a Possible Major Role for Enterotoxins. Sci. Rep. 2017, 7, 12508. [Google Scholar] [CrossRef] [PubMed]
- Kobmoo, N.; Wichadakul, D.; Arnamnart, N.; Vega, R.C.R.D.L.; Luangsa-Ard, J.-J.; Giraud, T. A Genome Scan of Diversifying Selection in Ophiocordyceps Zombie-Ant Fungi Suggests a Role for Enterotoxins in Coevolution and Host Specificity. Mol. Ecol. 2018, 27, 3582–3598. [Google Scholar] [CrossRef]
- Gaskell, E.A.; Smith, J.E.; Pinney, J.W.; Westhead, D.R.; McConkey, G.A. A Unique Dual Activity Amino Acid Hydroxylase in Toxoplasma Gondii. PLoS ONE 2009, 4, e4801. [Google Scholar] [CrossRef] [Green Version]
- Kamita, S.G.; Nagasaka, K.; Chua, J.W.; Shimada, T.; Mita, K.; Kobayashi, M.; Maeda, S.; Hammock, B.D. A Baculovirus-Encoded Protein Tyrosine Phosphatase Gene Induces Enhanced Locomotory Activity in a Lepidopteran Host. Proc. Natl. Acad. Sci. USA 2005, 102, 2584. [Google Scholar] [CrossRef] [Green Version]
- Katsuma, S.; Koyano, Y.; Kang, W.; Kokusho, R.; Kamita, S.G.; Shimada, T. The Baculovirus Uses a Captured Host Phosphatase to Induce Enhanced Locomotory Activity in Host Caterpillars. PLoS Pathog. 2012, 8, e1002644. [Google Scholar] [CrossRef] [Green Version]
- Hoover, K.; Grove, M.; Gardner, M.; Hughes, D.P.; McNeil, J.; Slavicek, J. A Gene for an Extended Phenotype. Science 2011, 333, 1401. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; van Houte, S.; Drees, G.F.; van Oers, M.M.; Ros, V.I.D. Parasitic Manipulation of Host Behaviour: Baculovirus SeMNPV EGT Facilitates Tree-Top Disease in Spodoptera Exigua Larvae by Extending the Time to Death. Insects 2015, 6, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawtin, R.E.; Zarkowska, T.; Arnold, K.; Thomas, C.J.; Gooday, G.W.; King, L.A.; Kuzio, J.A.; Possee, R.D. Liquefaction of Autographa Californica Nucleopolyhedrovirus-Infected Insects Is Dependent on the Integrity of Virus-Encoded Chitinase and Cathepsin Genes. Virology 1997, 238, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Adamo, S.A. Parasites: Evolution’s Neurobiologists. J. Exp. Biol. 2013, 216, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; De Fine Licht, H.H.; et al. Psychoactive Plant- and Mushroom-Associated Alkaloids from Two Behavior Modifying Cicada Pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef]
- de Bekker, C.; Ohm, R.A.; Loreto, R.G.; Sebastian, A.; Albert, I.; Merrow, M.; Brachmann, A.; Hughes, D.P. Gene Expression during Zombie Ant Biting Behavior Reflects the Complexity Underlying Fungal Parasitic Behavioral Manipulation. BMC Genom. 2015, 16, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bekker, C.; Will, I.; Hughes, D.P.; Brachmann, A.; Merrow, M. Daily Rhythms and Enrichment Patterns in the Transcriptome of the Behavior-Manipulating Parasite Ophiocordyceps Kimflemingiae. PLoS ONE 2017, 12, e0187170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron, D.G.; Ponton, F.; Marché, L.; Galeotti, N.; Renault, L.; Demey-Thomas, E.; Poncet, J.; Brown, S.P.; Jouin, P.; Thomas, F. ‘Suicide’ of Crickets Harbouring Hairworms: A Proteomics Investigation. Insect Mol. Biol. 2006, 15, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Biron, D.G.; Marché, L.; Ponton, F.; Loxdale, H.D.; Galéotti, N.; Renault, L.; Joly, C.; Thomas, F. Behavioural Manipulation in a Grasshopper Harbouring Hairworm: A Proteomics Approach. Proc. Biol. Sci. 2005, 272, 2117–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.P.; Brodeur, J.; Thomas, F. Host Manipulation by Parasites; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-964223-6. [Google Scholar]
- Kaiser, M.; Arvidson, R.; Zarivach, R.; Adams, M.E.; Libersat, F. Molecular Cross-Talk in a Unique Parasitoid Manipulation Strategy. Insect Biochem. Mol. Biol. 2019, 106, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Libersat, F. The Role of the Cerebral Ganglia in the Venom-Induced Behavioral Manipulation of Cockroaches Stung by the Parasitoid Jewel Wasp. J. Exp. Biol. 2015, 218, 1022–1027. [Google Scholar] [CrossRef] [Green Version]
- Loreto, R.G.; Hughes, D.P. The Metabolic Alteration and Apparent Preservation of the Zombie Ant Brain. J. Insect Physiol. 2019, 103918. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.C.; Korzan, W.J.; Carpenter, R.E.; Kuris, A.M.; Lafferty, K.D.; Summers, C.H.; Øverli, Ø. Parasite Manipulation of Brain Monoamines in California Killifish (Fundulus parvipinnis) by the Trematode Euhaplorchis Californiensis. Proc. Biol. Sci. 2009, 276, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.C.; Øverli, Ø. Brain-Encysting Trematodes and Altered Monoamine Activity in Naturally Infected Killifish Fundulus parvipinnis. J. Fish Biol. 2012, 81, 2213–2222. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Nakajima, K. Role of the Immune System in the Development of the Central Nervous System. Front. Neurosci. 2019, 13, 916. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Dissing-Olesen, L.; Stevens, B. New Insights on the Role of Microglia in Synaptic Pruning in Health and Disease. Curr. Opin. Neurobiol. 2016, 36, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottahedin, A.; Ardalan, M.; Chumak, T.; Riebe, I.; Ek, J.; Mallard, C. Effect of Neuroinflammation on Synaptic Organization and Function in the Developing Brain: Implications for Neurodevelopmental and Neurodegenerative Disorders. Front. Cell Neurosci. 2017, 11, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, L.M. Immune Proteins in Brain Development and Synaptic Plasticity. Neuron 2009, 64, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatz, C.J. MHC Class I: An Unexpected Role in Neuronal Plasticity. Neuron 2009, 64, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Stephan, A.H.; Barres, B.A.; Stevens, B. The Complement System: An Unexpected Role in Synaptic Pruning during Development and Disease. Annu. Rev. Neurosci. 2012, 35, 369–389. [Google Scholar] [CrossRef] [Green Version]
- Yirmiya, R.; Goshen, I. Immune Modulation of Learning, Memory, Neural Plasticity and Neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Lucin, K.M.; Wyss-Coray, T. Immune Activation in Brain Aging and Neurodegeneration: Too Much or Too Little? Neuron 2009, 64, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Lye, S.H.; Chtarbanova, S. Drosophila as a Model to Study Brain Innate Immunity in Health and Disease. Int. J. Mol. Sci. 2018, 19, 3922. [Google Scholar] [CrossRef] [Green Version]
- Nainu, F.; Salim, E.; Asri, R.M.; Hori, A.; Kuraishi, T. Neurodegenerative Disorders and Sterile Inflammation: Lessons from a Drosophila Model. J. Biochem. 2019, 166, 213–221. [Google Scholar] [CrossRef]
- Rittschof, C.C.; Schirmeier, S. Insect Models of Central Nervous System Energy Metabolism and Its Links to Behavior. Glia 2018, 66, 1160–1175. [Google Scholar] [CrossRef]
- Fuentes-Medel, Y.; Logan, M.A.; Ashley, J.; Ataman, B.; Budnik, V.; Freeman, M.R. Glia and Muscle Sculpt Neuromuscular Arbors by Engulfing Destabilized Synaptic Boutons and Shed Presynaptic Debris. PLoS Biol. 2009, 7, e1000184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.R. Drosophila Central Nervous System Glia. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurant, E. Keeping the CNS Clear: Glial Phagocytic Functions in Drosophila. Glia 2011, 59, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Purice, M.D.; Ray, A.; Münzel, E.J.; Pope, B.J.; Park, D.J.; Speese, S.D.; Logan, M.A. A Novel Drosophila Injury Model Reveals Severed Axons Are Cleared through a Draper/MMP-1 Signaling Cascade. eLife 2017, 6, e23611. [Google Scholar] [CrossRef] [PubMed]
- Dushay, M. Insect Infection and Immunity. Evolution, Ecology, and Mechanisms. Jens Rolff and Stuart Reynolds, Editors. Integr. Comp. Biol. 2010, 50, 908–909. [Google Scholar] [CrossRef]
- Hillyer, J.F. Insect Immunology and Hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Siva-Jothy, M.T.; Moret, Y.; Rolff, J. Insect Immunity: An Evolutionary Ecology Perspective. In Advances in Insect Physiology; Simpson, S.J., Ed.; Academic Press: New York, NY, USA, 2005; Volume 32, pp. 1–48. [Google Scholar]
- Cremer, S.; Armitage, S.A.O.; Schmid-Hempel, P. Social Immunity. Curr. Biol. 2007, 17, R693–R702. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C. Cellular and Molecular Mechanisms of Insect Immunity. Insect Physiol. Ecol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, P.O. Hemocytes of Insects: Their Morphology and Function. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2008; pp. 1787–1790. ISBN 978-1-4020-6359-6. [Google Scholar]
- Flajnik, M.F.; Kasahara, M. Origin and Evolution of the Adaptive Immune System: Genetic Events and Selective Pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.; Eleftherianos, I. Memory and Specificity in the Insect Immune System: Current Perspectives and Future Challenges. Front. Immunol. 2017, 8, 539. [Google Scholar] [CrossRef] [PubMed]
- Ferro, K.; Peuß, R.; Yang, W.; Rosenstiel, P.; Schulenburg, H.; Kurtz, J. Experimental Evolution of Immunological Specificity. Proc. Natl. Acad. Sci. USA 2019, 116, 20598–20604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadd, B.M.; Schmid-Hempel, P. Insect Immunity Shows Specificity in Protection upon Secondary Pathogen Exposure. Curr. Biol. 2006, 16, 1206–1210. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, G.; Farrell, G.; Kavanagh, K. Immune Priming: The Secret Weapon of the Insect World. Virulence 2020, 11, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Adamo, S.A. Bidirectional connections between the immune system and the nervous system in insects. In Insect Immunology; Beckage, N.E., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 129–149. ISBN 978-0-12-373976-6. [Google Scholar]
- Herniou, E.A.; Olszewski, J.A.; Cory, J.S.; O’Reilly, D.R. The Genome Sequence and Evolution of Baculoviruses. Annu. Rev. Entomol. 2003, 48, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, R. Insektentötende Pilze mit Besonderer Berücksichtigung der “Nonne”; P. Weber: Frankfurt, Germany, 1891. [Google Scholar]
- Goulson, D. Wipfelkrankheit: Modification of Host Behaviour during Baculoviral Infection. Oecologia 1997, 109, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. Host Manipulation by Parasites: Cases, Patterns, and Remaining Doubts. Front. Ecol. Evol. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, V.; Elkinton, J.S. Rainfall Effects on Transmission of Gypsy Moth (Lepidoptera: Lymantriidae) Nuclear Polyhedrosis Virus. Environ. Entomol. 1995, 24, 1144–1149. [Google Scholar] [CrossRef]
- van Houte, S.; van Oers, M.M.; Han, Y.; Vlak, J.M.; Ros, V.I.D. Baculovirus Infection Triggers a Positive Phototactic Response in Caterpillars to Induce ‘Tree-Top’ Disease. Biol. Lett. 2014, 10, 20140680. [Google Scholar] [CrossRef]
- Adamo, S.A.; Kovalko, I.; Easy, R.H.; Stoltz, D. A Viral Aphrodisiac in the Cricket Gryllus Texensis. J. Exp. Biol. 2014, 217, 1970–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, S.A. The Effects of the Stress Response on Immune Function in Invertebrates: An Evolutionary Perspective on an Ancient Connection. Horm. Behav. 2012, 62, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Aubert, A. Sickness and Behaviour in Animals: A Motivational Perspective. Neurosci. Biobehav. Rev. 1999, 23, 1029–1036. [Google Scholar] [CrossRef]
- Dantzer, R. Cytokine-Induced Sickness Behaviour: A Neuroimmune Response to Activation of Innate Immunity. Eur. J. Pharm. 2004, 500, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; Kelley, K.W. Stress and Immunity: An Integrated View of Relationships between the Brain and the Immune System. Life Sci. 1989, 44, 1995–2008. [Google Scholar] [CrossRef]
- Hart, B.L. Biological Basis of the Behavior of Sick Animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef]
- Sullivan, K.; Fairn, E.; Adamo, S.A. Sickness Behaviour in the Cricket Gryllus Texensis: Comparison with Animals across Phyla. Behav. Process. 2016, 128, 134–143. [Google Scholar] [CrossRef]
- Raina, A.K.; Adams, J.R. Gonad-Specific Virus of Corn Earworm. Nature 1995, 374, 770. [Google Scholar] [CrossRef]
- Burand, J.P.; Tan, W.; Kim, W.; Nojima, S.; Roelofs, W. Infection with the Insect Virus Hz-2v Alters Mating Behavior and Pheromone Production in Female Helicoverpa Zea Moths. J. Insect Sci. 2005, 5. [Google Scholar] [CrossRef] [Green Version]
- Maure, F.; Brodeur, J.; Ponlet, N.; Doyon, J.; Firlej, A.; Elguero, É.; Thomas, F. The Cost of a Bodyguard. Biol. Lett. 2011, 7, 843–846. [Google Scholar] [CrossRef]
- Dheilly, N.M.; Maure, F.; Ravallec, M.; Galinier, R.; Doyon, J.; Duval, D.; Leger, L.; Volkoff, A.-N.; Missé, D.; Nidelet, S.; et al. Who Is the Puppet Master? Replication of a Parasitic Wasp-Associated Virus Correlates with Host Behaviour Manipulation. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20142773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnoff, S.B.; Watson, D.W.; Gibson, D.M.; Kwan, E.C. Behavioral Effects of the Entomopathogenic Fungus, Entomophthora Muscae on Its Host Musca Domestica: Postural Changes in Dying Hosts and Gated Pattern of Mortality. J. Insect Physiol. 1995, 41, 895–903. [Google Scholar] [CrossRef]
- Goldstein, B. An Empusa Disease of Drosophila. Mycologia 1927, 19, 97–109. [Google Scholar] [CrossRef]
- Elya, C.; Lok, T.C.; Spencer, Q.E.; McCausland, H.; Martinez, C.C.; Eisen, M. Robust Manipulation of the Behavior of Drosophila Melanogaster by a Fungal Pathogen in the Laboratory. eLife 2018, 7, e34414. [Google Scholar] [CrossRef] [PubMed]
- Coyle, M.C.; Elya, C.N.; Bronski, M.J.; Eisen, M.B. Entomophthovirus: An Insect-Derived Iflavirus That Infects a Behavior Manipulating Fungal Pathogen of Dipterans. bioRxiv 2018, 371526. [Google Scholar] [CrossRef] [Green Version]
- Araújo, J.P.M.; Hughes, D.P. The Fungal Spore: Its Organization and Role in the Ecosystem. In The Fungal Community: Its Organization and Role in the Ecosystem; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 359–367. [Google Scholar]
- Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hywel-Jones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. The Life of a Dead Ant: The Expression of an Adaptive Extended Phenotype. Am. Nat. 2009, 174, 424–433. [Google Scholar] [CrossRef]
- de Bekker, C. Ophiocordyceps–Ant Interactions as an Integrative Model to Understand the Molecular Basis of Parasitic Behavioral Manipulation. Curr. Opin. Insect Sci. 2019. [Google Scholar] [CrossRef]
- de Bekker, C.; Quevillon, L.E.; Smith, P.B.; Fleming, K.R.; Ghosh, D.; Patterson, A.D.; Hughes, D.P. Species-Specific Ant Brain Manipulation by a Specialized Fungal Parasite. BMC Evol. Biol. 2014, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.P.; Andersen, S.B.; Hywel-Jones, N.L.; Himaman, W.; Billen, J.; Boomsma, J.J. Behavioral Mechanisms and Morphological Symptoms of Zombie Ants Dying from Fungal Infection. BMC Ecol. 2011, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreto, R.G.; Araújo, J.P.M.; Kepler, R.M.; Fleming, K.R.; Moreau, C.S.; Hughes, D.P. Evidence for Convergent Evolution of Host Parasitic Manipulation in Response to Environmental Conditions. Evolution 2018, 72, 2144–2155. [Google Scholar] [CrossRef]
- Fredericksen, M.A.; Zhang, Y.; Hazen, M.L.; Loreto, R.G.; Mangold, C.A.; Chen, D.Z.; Hughes, D.P. Three-Dimensional Visualization and a Deep-Learning Model Reveal Complex Fungal Parasite Networks in Behaviorally Manipulated Ants. Proc. Natl. Acad. Sci. USA 2017, 201711673. [Google Scholar] [CrossRef] [Green Version]
- Mangold, C.A.; Ishler, M.J.; Loreto, R.G.; Hazen, M.L.; Hughes, D.P. Zombie Ant Death Grip Due to Hypercontracted Mandibular Muscles. J. Exp. Biol. 2019, 222, jeb200683. [Google Scholar] [CrossRef] [Green Version]
- Will, I.; Das, B.; Trinh, T.; Brachmann, A.; Ohm, R.A.; de Bekker, C. Genetic Underpinnings of Host Manipulation by Ophiocordyceps as Revealed by Comparative Transcriptomics. G3 Genes Genomes Genet 2020, 10, 2275–2296. [Google Scholar] [CrossRef]
- Małagocka, J.; Grell, M.N.; Lange, L.; Eilenberg, J.; Jensen, A.B. Transcriptome of an Entomophthoralean Fungus (Pandora Formicae) Shows Molecular Machinery Adjusted for Successful Host Exploitation and Transmission. J. Invertebr. Pathol. 2015, 128, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Małagocka, J.; Jensen, A.B.; Eilenberg, J. Pandora Formicae, a Specialist Ant Pathogenic Fungus: New Insights into Biology and Taxonomy. J. Invertebr. Pathol. 2017, 143, 108–114. [Google Scholar] [CrossRef]
- Boer, P. Observations of Summit Disease in Formica Rufa LINNAEUS, 1761 (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 63–66. [Google Scholar]
- Marikovsky, P.I. On Some Features of Behavior of the Ants Formica Rufa L. Infected with Fungous Disease. Ins. Soc. 1962, 9, 173–179. [Google Scholar] [CrossRef]
- Hughes, D.P.; Araújo, J.P.M.; Loreto, R.G.; Quevillon, L.; de Bekker, C.; Evans, H.C. From So Simple a Beginning: The Evolution of Behavioral Manipulation by Fungi. Adv. Genet. 2016, 94, 437–469. [Google Scholar] [CrossRef]
- Soper, R.S.; Delyzer, A.J.; Smith, L.F.R. The Genus Massospora Entomopathogenic for Cicadas. Part. II. Biology of Massospora Levispora and Its Host Okanagana Rimosa, with Notes on Massospora Cicadina on the Periodical Cicadas. Ann. Entomol. Soc. Am. 1976, 69, 89–95. [Google Scholar] [CrossRef]
- Cooley, J.R.; Marshall, D.C.; Hill, K.B.R. A Specialized Fungal Parasite (Massospora cicadina) Hijacks the Sexual Signals of Periodical Cicadas (Hemiptera: Cicadidae: Magicicada). Sci. Rep. 2018, 8, 1432. [Google Scholar] [CrossRef]
- Nair, K.S.S.; McEwen, F.L. Strongwellsea Castrans (Phycomycetes: Entomophthoraceae), a Fungal Parasite of the Adult Cabbage Maggot, Hylemya Brassicae (Diptera: Anthomyiidae). J. Invertebr. Pathol. 1973, 22, 442–449. [Google Scholar] [CrossRef]
- Haspel, G.; Rosenberg, L.A.; Libersat, F. Direct Injection of Venom by a Predatory Wasp into Cockroach Brain. J. Neurobiol. 2003, 56, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, F.X. Ampulex compressa (Fabr.), a cockroach-hunting wasp introduced from New Caledonia into Hawaii. Proc. Hawaii. Entomol. Soc. 1942, 11, 221–233. [Google Scholar]
- Libersat, F.; Delago, A.; Gal, R. Manipulation of Host Behavior by Parasitic Insects and Insect Parasites. Annu. Rev. Entomol. 2008, 54, 189–207. [Google Scholar] [CrossRef]
- Weisel-Eichler, A.; Libersat, F. Are Monoaminergic Systems Involved in the Lethargy Induced by a Parasitoid Wasp in the Cockroach Prey? J. Comp. Physiol. A 2002, 188, 315–324. [Google Scholar] [CrossRef]
- Eberhard, W.G. Spider Manipulation by a Wasp Larva. Nature 2000, 406, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, W.G. The Natural History and Behavior of Hymenoepimecis Argyraphaga (Hymenoptera: Ichneumonidae) a Parasitoid of Plesiometa Argyra (Araneae: Tetragnathidae). J. Hymenopt. Res. 2000, 9, 220–240. [Google Scholar]
- Eberhard, W.G. Recovery of Spiders from the Effects of Parasitic Wasps: Implications for Fine-Tuned Mechanisms of Manipulation. Anim. Behav. 2010, 79, 375–383. [Google Scholar] [CrossRef]
- Eberhard, W.G. The Polysphinctine Wasps Acrotaphus Tibialis, Eruga ca. Gutfreundi, and Hymenoepimecis Tedfordi (Hymenoptera, Ichneumonidae, Pimplinae) Induce Their Host Spiders to Build Modified Webs. Ann. Entomol. Soc. Am. 2013, 106, 652–660. [Google Scholar] [CrossRef]
- Thomas, F.; Schmidt-Rhaesa, A.; Martin, G.; Manu, C.; Durand, P.; Renaud, F. Do Hairworms (Nematomorpha) Manipulate the Water Seeking Behaviour of Their Terrestrial Hosts? J. Evol. Biol. 2002, 15, 356–361. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Morris, A.K. Altered Behavior of Parasitized Killifish Increases Susceptibility to Predation by Bird Final Hosts. Ecology 1996, 77, 1390–1397. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.E. Euhaplorchis calif or niensis n,g., n.sp., Heterophyidae, Trematoda, with notes on its life-cycle. Trans. Am. Microsc. Soc. 1950, 69, 194–209. [Google Scholar] [CrossRef]
- Helluy, S. Host-parasite relations of the trematode Microphallus papillorobustus (Rankin, 1940). I. Penetration of cercariae and relationship of the metacercariae with the nervous tissue of Gammarus, intermediate hosts. Ann. Parasitol. Hum. Comp. 1982, 57, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helluy, S. Host-parasite relations of the trematode Microphallus papillorobustus (Rankin 1940). III Factors involved in the behavioral changes of the Gammarus, intermediate hosts and predator tests. Ann. Parasitol. Hum. Comp. 1984, 59, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Helluy, S.; Thomas, F. Effects of Microphallus Papillorobustus (Platyhelminthes: Trematoda) on Serotonergic Immunoreactivity and Neuronal Architecture in the Brain of Gammarus Insensibilis (Crustacea: Amphipoda). Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 563–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helluy, S.; Thomas, F. Parasitic Manipulation and Neuroinflammation: Evidence from the System Microphallus Papillorobustus (Trematoda)—Gammarus (Crustacea). Parasit Vectors 2010, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Berdoy, M.; Webster, J.P.; Macdonald, D.W. Fatal Attraction in Rats Infected with Toxoplasma Gondii. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 1591–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martynowicz, J.; Augusto, L.; Wek, R.C.; Boehm, S.L.; Sullivan, W.J. Guanabenz Reverses a Key Behavioral Change Caused by Latent Toxoplasmosis in Mice by Reducing Neuroinflammation. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Berenreiterová, M.; Flegr, J.; Kuběna, A.A.; Němec, P. The Distribution of Toxoplasma Gondii Cysts in the Brain of a Mouse with Latent Toxoplasmosis: Implications for the Behavioral Manipulation Hypothesis. PLoS ONE 2011, 6, e28925. [Google Scholar] [CrossRef] [PubMed]
- Prandovszky, E.; Gaskell, E.; Martin, H.; Dubey, J.P.; Webster, J.P.; McConkey, G.A. The Neurotropic Parasite Toxoplasma Gondii Increases Dopamine Metabolism. PLoS ONE 2011, 6, e23866. [Google Scholar] [CrossRef]
- Stibbs, H.H. Changes in Brain Concentrations of Catecholamines and Indoleamines in Toxoplasma Gondii Infected Mice. Ann. Trop. Med. Parasitol. 1985, 79, 153–157. [Google Scholar] [CrossRef]
- Skallová, A.; Kodym, P.; Frynta, D.; Flegr, J. The Role of Dopamine in Toxoplasma-Induced Behavioural Alterations in Mice: An Ethological and Ethopharmacological Study. Parasitology 2006, 133, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Flegr, J.; Markoš, A. Masterpiece of Epigenetic Engineering—How Toxoplasma Gondii Reprogrammes Host Brains to Change Fear to Sexual Attraction. Mol. Ecol. 2014, 23, 5934–5936. [Google Scholar] [CrossRef]
- Lim, A.; Kumar, V.; Dass, S.A.H.; Vyas, A. Toxoplasma Gondii Infection Enhances Testicular Steroidogenesis in Rats. Mol. Ecol. 2013, 22, 102–110. [Google Scholar] [CrossRef]
- Dass, S.A.H.; Vyas, A. Toxoplasma Gondii Infection Reduces Predator Aversion in Rats through Epigenetic Modulation in the Host Medial Amygdala. Mol. Ecol. 2014, 23, 6114–6122. [Google Scholar] [CrossRef] [PubMed]
- Yong, E. Is This Fungus Using a Virus to Control an Animal’s Mind?—The Atlantic. Available online: https://www.theatlantic.com/science/archive/2018/07/is-this-fungus-using-a-virus-to-mind-control-an-animal/565982/ (accessed on 13 October 2018).
- Rossi, M.; Castiglioni, P.; Hartley, M.-A.; Eren, R.O.; Prével, F.; Desponds, C.; Utzschneider, D.T.; Zehn, D.; Cusi, M.G.; Kuhlmann, F.M.; et al. Type I Interferons Induced by Endogenous or Exogenous Viral Infections Promote Metastasis and Relapse of Leishmaniasis. Proc. Natl. Acad. Sci. USA 2017, 114, 4987–4992. [Google Scholar] [CrossRef] [Green Version]
- Adaui, V.; Lye, L.-F.; Akopyants, N.S.; Zimic, M.; Llanos-Cuentas, A.; Garcia, L.; Maes, I.; De Doncker, S.; Dobson, D.E.; Arevalo, J.; et al. Association of the Endobiont Double-Stranded RNA Virus LRV1 With Treatment Failure for Human Leishmaniasis Caused by Leishmania Braziliensis in Peru and Bolivia. J. Infect. Dis. 2016, 213, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Bourreau, E.; Ginouves, M.; Prévot, G.; Hartley, M.-A.; Gangneux, J.-P.; Robert-Gangneux, F.; Dufour, J.; Sainte-Marie, D.; Bertolotti, A.; Pratlong, F.; et al. Presence of Leishmania RNA Virus 1 in Leishmania Guyanensis Increases the Risk of First-Line Treatment Failure and Symptomatic Relapse. J. Infect. Dis. 2016, 213, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Grybchuk, D.; Akopyants, N.S.; Kostygov, A.Y.; Konovalovas, A.; Lye, L.-F.; Dobson, D.E.; Zangger, H.; Fasel, N.; Butenko, A.; Frolov, A.O.; et al. Viral Discovery and Diversity in Trypanosomatid Protozoa with a Focus on Relatives of the Human Parasite Leishmania. Proc. Natl. Acad. Sci. USA 2018, 115, E506–E515. [Google Scholar] [CrossRef] [Green Version]
- Riegler, M.; O’Neill, S.L. The Genus Wolbachia. In The Prokaryotes; Dr, M.D.P., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 547–561. ISBN 978-0-387-25495-1. [Google Scholar]
- Miller, W.J. Bugs in Transition: The Dynamic World of Wolbachia in Insects. PLoS Genet. 2013, 9, e1004069. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Albertson, R.; Casper-Lindley, C.; Cao, J.; Tram, U.; Sullivan, W. Symmetric and Asymmetric Mitotic Segregation Patterns Influence Wolbachia Distribution in Host Somatic Tissue. J. Cell Sci. 2009, 122, 4570–4583. [Google Scholar] [CrossRef] [Green Version]
- Albertson, R.; Tan, V.; Leads, R.R.; Reyes, M.; Sullivan, W.; Casper-Lindley, C. Mapping Wolbachia Distributions in the Adult Drosophila Brain. Cell. Microbiol. 2013, 15, 1527–1544. [Google Scholar] [CrossRef] [PubMed]
- Casper-Lindley, C.; Kimura, S.; Saxton, D.S.; Essaw, Y.; Simpson, I.; Tan, V.; Sullivan, W. Rapid Fluorescence-Based Screening for Wolbachia Endosymbionts in Drosophila Germ Line and Somatic Tissues. Appl. Environ. Microbiol. 2011, 77, 4788–4794. [Google Scholar] [CrossRef] [Green Version]
- Strunov, A.; Schneider, D.I.; Albertson, R.; Miller, W.J. Restricted Distribution and Lateralization of Mutualistic Wolbachia in the Drosophila Brain. Cell. Microbiol. 2016, 19, e12639. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Sehgal, A.; Williams, J.A.; Wang, Y.-F. Wolbachia Affects Sleep Behavior in Drosophila Melanogaster. J. Insect Physiol. 2018, 107, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Evans, O.; Caragata, E.P.; McMeniman, C.J.; Woolfit, M.; Green, D.C.; Williams, C.R.; Franklin, C.E.; O’Neill, S.L.; McGraw, E.A. Increased Locomotor Activity and Metabolism of Aedes Aegypti Infected with a Life-Shortening Strain of Wolbachia Pipientis. J. Exp. Biol. 2009, 212, 1436–1441. [Google Scholar] [CrossRef] [Green Version]
- Rohrscheib, C.E.; Bondy, E.; Josh, P.; Riegler, M.; Eyles, D.; van Swinderen, B.; Weible, M.W.; Brownlie, J.C. Wolbachia Influences the Production of Octopamine and Affects Drosophila Male Aggression. Appl. Environ. Microbiol. 2015, 81, 4573–4580. [Google Scholar] [CrossRef] [Green Version]
- Vale, P.F.; Jardine, M.D. Sex-Specific Behavioural Symptoms of Viral Gut Infection and Wolbachia in Drosophila Melanogaster. J. Insect Physiol. 2015, 82, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Templé, N.; Richard, F.-J. Intra-Cellular Bacterial Infections Affect Learning and Memory Capacities of an Invertebrate. Front. Zool. 2015, 12, 36. [Google Scholar] [CrossRef] [Green Version]
- Farahani, H.K.; Ashouri, A.; Goldansaz, S.H.; Shapiro, M.S.; Pierre, J.-S.; Baaren, J. van Decrease of Memory Retention in a Parasitic Wasp: An Effect of Host Manipulation by Wolbachia? Insect Sci. 2017, 24, 569–583. [Google Scholar] [CrossRef]
- Peng, Y.; Nielsen, J.E.; Cunningham, J.P.; McGraw, E.A. Wolbachia Infection Alters Olfactory-Cued Locomotion in Drosophila Spp. Appl. Environ. Microbiol. 2008, 74, 3943–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Wang, Y. Infection of Wolbachia May Improve the Olfactory Response of Drosophila. Chin. Sci. Bull. 2009, 54, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Crespigny, F.E.C.D.; Pitt, T.D.; Wedell, N. Increased Male Mating Rate in Drosophila Is Associated with Wolbachia Infection. J. Evol. Biol. 2006, 19, 1964–1972. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.J.; Schneider, D. Endosymbiotic Microbes as Adaptive Manipulators of Arthropod Behavior and Natural Driving Sources of Host Speciation; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-177462-1. [Google Scholar]
- Cenit, M.C.; Sanz, Y.; Codoñer-Franch, P. Influence of Gut Microbiota on Neuropsychiatric Disorders. World J. Gastroenterol. 2017, 23, 5486–5498. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Mahony, S.M. The Microbiome-Gut-Brain Axis: From Bowel to Behavior. Neurogastroenterol. Motil. 2011, 23, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P.; Kunze, W.A.; Bienenstock, J. On Communication between Gut Microbes and the Brain. Curr. Opin. Gastroenterol. 2012, 28, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The Gut Microbiome in Health and in Disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef]
- Archie, E.A.; Theis, K.R. Animal Behaviour Meets Microbial Ecology. Anim. Behav. 2011, 82, 425–436. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Gerardo, N.M.; Inouye, D.W.; Medina, M.; Xavier, J.B. Animal Behavior and the Microbiome. Science 2012, 338, 198–199. [Google Scholar] [CrossRef]
- Burgener, N.; Dehnhard, M.; Hofer, H.; East, M.L. Does Anal Gland Scent Signal Identity in the Spotted Hyaena? Anim. Behav. 2009, 77, 707–715. [Google Scholar] [CrossRef]
- Theis, K.R.; Venkataraman, A.; Dycus, J.A.; Koonter, K.D.; Schmitt-Matzen, E.N.; Wagner, A.P.; Holekamp, K.E.; Schmidt, T.M. Symbiotic Bacteria Appear to Mediate Hyena Social Odors. Proc. Natl. Acad. Sci. USA 2013, 110, 19832–19837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesperance, D.N.; Broderick, N.A. Microbiomes as Modulators of Drosophila Melanogaster Homeostasis and Disease. Curr. Opin. Insect Sci. 2020, 39, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Masuzzo, A.; Manière, G.; Viallat-Lieutaud, A.; Avazeri, É.; Zugasti, O.; Grosjean, Y.; Kurz, C.L.; Royet, J. Peptidoglycan-Dependent NF-ΚB Activation in a Small Subset of Brain Octopaminergic Neurons Controls Female Oviposition. eLife 2019, 8, e50559. [Google Scholar] [CrossRef] [PubMed]
- Masuzzo, A.; Montanari, M.; Kurz, L.; Royet, J. How Bacteria Impact Host Nervous System and Behaviors: Lessons from Flies and Worms. Trends Neurosci. 2020, 43, 998–1010. [Google Scholar] [CrossRef]
- Schretter, C.E.; Vielmetter, J.; Bartos, I.; Marka, Z.; Marka, S.; Argade, S.; Mazmanian, S.K. A Gut Microbial Factor Modulates Locomotor Behaviour in Drosophila. Nature 2018, 563, 402–406. [Google Scholar] [CrossRef]
- Araújo, J.P.M.; Hughes, D.P. Chapter One—Diversity of Entomopathogenic Fungi: Which Groups Conquered the Insect Body? In Advances in Genetics; Genetics and Molecular Biology of Entomopathogenic Fungi; Lovett, B., St. Leger, R.J., Eds.; Academic Press: New York, NY, USA, 2016; Volume 94, pp. 1–39. [Google Scholar]
- Lezi, E.; Zhou, T.; Koh, S.; Chuang, M.; Sharma, R.; Pujol, N.; Chisholm, A.D.; Eroglu, C.; Matsunami, H.; Yan, D. An Antimicrobial Peptide and Its Neuronal Receptor Regulate Dendrite Degeneration in Aging and Infection. Neuron 2018, 97, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Chtarbanova, S.; Petersen, A.J.; Ganetzky, B. Dnr1 Mutations Cause Neurodegeneration in Drosophila by Activating the Innate Immune Response in the Brain. Proc. Natl. Acad. Sci. USA 2013, 110, E1752–E1760. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gordesky-Gold, B.; Leney-Greene, M.; Weinbren, N.L.; Tudor, M.; Cherry, S. Inflammation-Induced, STING-Dependent Autophagy Restricts Zika Virus Infection in the Drosophila Brain. Cell Host Microbe 2018, 24, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Moore, J. Parasites and the Behavior of Animals; Oxford University Press: New York, NY, USA, 2002; ISBN 978-0-19-514653-0. [Google Scholar]
- Libersat, F.; Kaiser, M.; Emanuel, S. Mind Control: How Parasites Manipulate Cognitive Functions in Their Insect Hosts. Front. Psychol. 2018, 9, 572. [Google Scholar] [CrossRef]
- Kato, K.; Awasaki, T.; Ito, K. Neuronal Programmed Cell Death Induces Glial Cell Division in the Adult Drosophila Brain. Development 2009, 136, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Katzenberger, R.J.; Loewen, C.A.; Wassarman, D.R.; Petersen, A.J.; Ganetzky, B.; Wassarman, D.A. A Drosophila Model of Closed Head Traumatic Brain Injury. Proc. Natl. Acad. Sci. USA 2013, 110, E4152–E4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzenberger, R.J.; Ganetzky, B.; Wassarman, D.A. Age and Diet Affect Genetically Separable Secondary Injuries That Cause Acute Mortality Following Traumatic Brain Injury in Drosophila. G3 (Bethesda) 2016, 6, 4151–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzenberger, R.J.; Chtarbanova, S.; Rimkus, S.A.; Fischer, J.A.; Kaur, G.; Seppala, J.M.; Swanson, L.C.; Zajac, J.E.; Ganetzky, B.; Wassarman, D.A. Death Following Traumatic Brain Injury in Drosophila Is Associated with Intestinal Barrier Dysfunction. eLife 2015, 4, e04790. [Google Scholar] [CrossRef] [PubMed]
- Balduf, W.V. The Bionomics of Dinocampus Coccinellæ Schrank. Ann. Entomol. Soc. Am. 1926, 19, 465–498. [Google Scholar] [CrossRef]
- Zhan, L.; Xie, Q.; Tibbetts, R.S. Opposing Roles of P38 and JNK in a Drosophila Model of TDP-43 Proteinopathy Reveal Oxidative Stress and Innate Immunity as Pathogenic Components of Neurodegeneration. Hum. Mol. Genet. 2015, 24, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Shieh, S.-Y.; Bonini, N.M. Genes and Pathways Affected by CAG-Repeat RNA-Based Toxicity in Drosophila. Hum. Mol. Genet 2011, 20, 4810–4821. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.K.; Tapadia, M.G. Yorkie Regulates Neurodegeneration Through Canonical Pathway and Innate Immune Response. Mol. Neurobiol. 2018, 55, 1193–1207. [Google Scholar] [CrossRef]
- Harris, N.; Braiser, D.J.; Dickman, D.K.; Fetter, R.D.; Tong, A.; Davis, G.W. The Innate Immune Receptor PGRP-LC Controls Presynaptic Homeostatic Plasticity. Neuron 2015, 88, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.; Fetter, R.D.; Brasier, D.J.; Tong, A.; Davis, G.W. Molecular Interface of Neuronal Innate Immunity, Synaptic Vesicle Stabilization, and Presynaptic Homeostatic Plasticity. Neuron 2018, 100, 1163–1179.e4. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.A.; Sathyanarayanan, S.; Hendricks, J.C.; Sehgal, A. Interaction Between Sleep and the Immune Response in Drosophila: A Role for the NFκB Relish. Sleep 2007, 30, 389–400. [Google Scholar] [CrossRef]
- Kuo, T.-H.; Pike, D.H.; Beizaeipour, Z.; Williams, J.A. Sleep Triggered by an Immune Response in Drosophila Is Regulated by the Circadian Clock and Requires the NFkappaB Relish. BMC Neurosci. 2010, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Toda, H.; Williams, J.A.; Gulledge, M.; Sehgal, A. A Sleep-Inducing Gene, Nemuri, Links Sleep and Immune Function in Drosophila. Science 2019, 363, 509–515. [Google Scholar] [CrossRef]
- Dissel, S.; Seugnet, L.; Thimgan, M.S.; Silverman, N.; Angadi, V.; Thacher, P.V.; Burnham, M.M.; Shaw, P.J. Differential Activation of Immune Factors in Neurons and Glia Contribute to Individual Differences in Resilience/Vulnerability to Sleep Disruption. Brain Behav. Immun. 2015, 47, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barajas-Azpeleta, R.; Wu, J.; Gill, J.; Welte, R.; Seidel, C.; McKinney, S.; Dissel, S.; Si, K. Antimicrobial Peptides Modulate Long-Term Memory. PLoS Genet. 2018, 14, e1007440. [Google Scholar] [CrossRef] [Green Version]
- Rivero, A. Nitric Oxide: An Antiparasitic Molecule of Invertebrates. Trends Parasitol. 2006, 22, 219–225. [Google Scholar] [CrossRef]
- Jacklet, J.W. Nitric Oxide Signaling in Invertebrates. Invertebr. Neurosci. 1997, 3, 1–14. [Google Scholar] [CrossRef]
- Weber, A.N.R.; Tauszig-Delamasure, S.; Hoffmann, J.A.; Lelièvre, E.; Gascan, H.; Ray, K.P.; Morse, M.A.; Imler, J.-L.; Gay, N.J. Binding of the Drosophila Cytokine Spätzle to Toll Is Direct and Establishes Signaling. Nat. Immunol. 2003, 4, 794–800. [Google Scholar] [CrossRef] [PubMed]
- DeLotto, Y.; DeLotto, R. Proteolytic Processing of the Drosophila Spätzle Protein by Easter Generates a Dimeric NGF-like Molecule with Ventralising Activity. Mech. Dev. 1998, 72, 141–148. [Google Scholar] [CrossRef]
- Sutcliffe, B.; Forero, M.G.; Zhu, B.; Robinson, I.M.; Hidalgo, A. Neuron-Type Specific Functions of DNT1, DNT2 and Spz at the Drosophila Neuromuscular Junction. PLoS ONE 2013, 8, e75902. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Pennack, J.A.; McQuilton, P.; Forero, M.G.; Mizuguchi, K.; Sutcliffe, B.; Gu, C.-J.; Fenton, J.C.; Hidalgo, A. Drosophila Neurotrophins Reveal a Common Mechanism for Nervous System Formation. PLoS Biol. 2008, 6, e284. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Guldner, E.; Renaud, F. Differential Parasite (Trematoda) Encapsulation in Gammarus Aequicauda (Amphipoda). J. Parasitol. 2000, 86, 650–654. [Google Scholar] [CrossRef]
- Shen, W.; Ganetzky, B. Autophagy Promotes Synapse Development in Drosophila. J. Cell Biol. 2009, 187, 71–79. [Google Scholar] [CrossRef] [Green Version]
- McDonald, M.J.; Rosbash, M. Microarray Analysis and Organization of Circadian Gene Expression in Drosophila. Cell 2001, 107, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Lazzaro, B.P.; Sceurman, B.K.; Clark, A.G. Genetic Basis of Natural Variation in D. Melanogaster Antibacterial Immunity. Science 2004, 303, 1873–1876. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Terry, E.E.; Fejer, E.; Gamba, D.; Hartmann, N.; Logsdon, J.; Michalski, D.; Rois, L.E.; Scuderi, M.J.; Kunst, M.; et al. Achilles Is a Circadian Clock-Controlled Gene That Regulates Immune Function in Drosophila. Brain Behav. Immun. 2017, 61, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Hartenstein, V. Morphological Diversity and Development of Glia in Drosophila. Glia 2011, 59, 1237–1252. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Urban, J.; Technau, G.M. Distribution, Classification, and Development OfDrosophila Glial Cells in the Late Embryonic and Early Larval Ventral Nerve Cord. Rouxs Arch. Dev. Biol. 1995, 204, 284–307. [Google Scholar] [CrossRef] [PubMed]
- Edenfeld, G.; Stork, T.; Klämbt, C. Neuron-Glia Interaction in the Insect Nervous System. Curr. Opin. Neurobiol. 2005, 15, 34–39. [Google Scholar] [CrossRef]
- Kremer, M.C.; Jung, C.; Batelli, S.; Rubin, G.M.; Gaul, U. The Glia of the Adult Drosophila Nervous System. Glia 2017, 65, 606–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limmer, S.; Weiler, A.; Volkenhoff, A.; Babatz, F.; Klämbt, C. The Drosophila Blood-Brain Barrier: Development and Function of a Glial Endothelium. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, A.J.; Rimkus, S.A.; Wassarman, D.A. ATM Kinase Inhibition in Glial Cells Activates the Innate Immune Response and Causes Neurodegeneration in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, E656–E664. [Google Scholar] [CrossRef] [Green Version]
- Kounatidis, I.; Chtarbanova, S.; Cao, Y.; Hayne, M.; Jayanth, D.; Ganetzky, B.; Ligoxygakis, P. NF-ΚB Immunity in the Brain Determines Fly Lifespan in Healthy Aging and Age-Related Neurodegeneration. Cell Rep. 2017, 19, 836–848. [Google Scholar] [CrossRef]
- Kemp, M.W.; Massey, R.C. The Use of Insect Models to Study Human Pathogens. Drug Discov. Today Dis. Models 2007, 4, 105–110. [Google Scholar] [CrossRef]
- Scully, L.R.; Bidochka, M.J. Developing Insect Models for the Study of Current and Emerging Human Pathogens. FEMS Microbiol. Lett. 2006, 263, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Loreto, R.G.; Elliot, S.L.; Freitas, M.L.R.; Pereira, T.M.; Hughes, D.P. Long-Term Disease Dynamics for a Specialized Parasite of Ant Societies: A Field Study. PLoS ONE 2014, 9, e103516. [Google Scholar] [CrossRef] [Green Version]
- Solá Gracia, E.; de Bekker, C.; Hanks, E.M.; Hughes, D.P. Within the Fortress: A Specialized Parasite Is Not Discriminated against in a Social Insect Society. PLoS ONE 2018, 13, e0193536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroeymeyt, N.; Grasse, A.V.; Crespi, A.; Mersch, D.P.; Cremer, S.; Keller, L. Social Network Plasticity Decreases Disease Transmission in a Eusocial Insect. Science 2018, 362, 941–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.P. On the Origins of Parasite-Extended Phenotypes. Integr. Comp. Biol. 2014, 54, 210–217. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Host | Parasite | Behavior | Mechanism(s) | Refs |
|---|---|---|---|---|
| Lepidoptera, Diptera, Hymenoptera, etc. | Baculoviruses | Tree climbing, enhanced locomotor activity and liquification | Expression of virus-specific genes, egt, ptp, chitinase, and cathepsin | [4,5,6,7,8,48,49,50,51,52,53] |
| Gryllus texensis | IIV-6/CrIV | Decreased egg production and sperm motility; maintained sexual behavior | Decreased expression of the virucidal enzyme phenoloxidase | [54] |
| Helicoverpa zea | HzNv-2 | Persistent calling behavior; increased pheromone production and contacts with males in infected females | Unknown | [61,62] |
| Ladybeetle | Dinocampus coccinellae/ DcPV | Bodyguard behavior/paralysis | Viral replication in host nervous tissue; neuroinflammation | [63,64] |
| House flies/fruit flies | Entomophthora muscae/DmEV | Attach to an elevated surface via the proboscides and raise wings, allowing for fungal spore dispersal | Unknown | [65,66,67,68] |
| Ants (Camponotus) | Ophiocordyceps unilateralis | Impaired motor control, convulsions; travel to areas optimal for fungal growth; circadian-synchronized biting behavior | Fungal production of enterotoxins and neuroactive compounds; altered host gene expression; formation of dense fungal network | [1,11,12,18,69,70,71,72,73,74,75,76,77] |
| Ants (Formica) | Pandora formica | Summit disease and biting behavior | Unknown | [78,79,80,81,82] |
| Periodical cicadas | Massospora cicadina Massospora platypediae Massospora levispora | Irregular flying and crawling behavior; altered sexual behavior | M. cicadina—production of the plant amphetamine cathinone; M. platypediae and M. levispora—production of psilocybin | [10,82,83,84] |
| Hylemya brassicae Hylemya platura | Strongwellsea castrans | Irregular flying and crawling behavior | Unknown | [85] |
| Periplaneta americana | Ampulex compressa | Transient paralysis and self-grooming | Venom containing neuromodulators | [16,17,86,87,88,89] |
| Plesiometa argyra | Hymenoepimecis argyraphaga | Altered web construction | Hypothesized venom-based modulation | [90,91,92,93] |
| Grasshoppers/crickets | Spinochordodes tellinii/ Paragordius tricuspidatus | Water-seeking behavior | Production of mimetic molecules (e.g., Wnt signaling modulators) and proteins involved in neurotransmission and apoptosis | [13,14,94] |
| Killifish | Euhaplorchis californiensis | Rapid swimming and “flashing”, increasing predation by birds | Altered monoamine signaling | [19,20,95,96] |
| Gammarids | Microphallus papillorobustus | Altered responses to environmental stimuli, increasing predation by birds | Altered monoamine signaling and neuroinflammation (i.e., NO synthesis) | [97,98,99,100] |
| Rodents | Toxoplasma gondii | Altered innate fear responses to cat odor | Alterations in dopamine and testosterone synthesis, epigenetic modifications, cyst formation, and neuroinflammation | [3,101,102,103,104,105,106,107,108,109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangold, C.A.; Hughes, D.P. Insect Behavioral Change and the Potential Contributions of Neuroinflammation—A Call for Future Research. Genes 2021, 12, 465. https://doi.org/10.3390/genes12040465
Mangold CA, Hughes DP. Insect Behavioral Change and the Potential Contributions of Neuroinflammation—A Call for Future Research. Genes. 2021; 12(4):465. https://doi.org/10.3390/genes12040465
Chicago/Turabian StyleMangold, Colleen A., and David P. Hughes. 2021. "Insect Behavioral Change and the Potential Contributions of Neuroinflammation—A Call for Future Research" Genes 12, no. 4: 465. https://doi.org/10.3390/genes12040465
APA StyleMangold, C. A., & Hughes, D. P. (2021). Insect Behavioral Change and the Potential Contributions of Neuroinflammation—A Call for Future Research. Genes, 12(4), 465. https://doi.org/10.3390/genes12040465