
Regulation of Fruit Growth in a Peach Slow Ripening Phenotype

 , and
, and 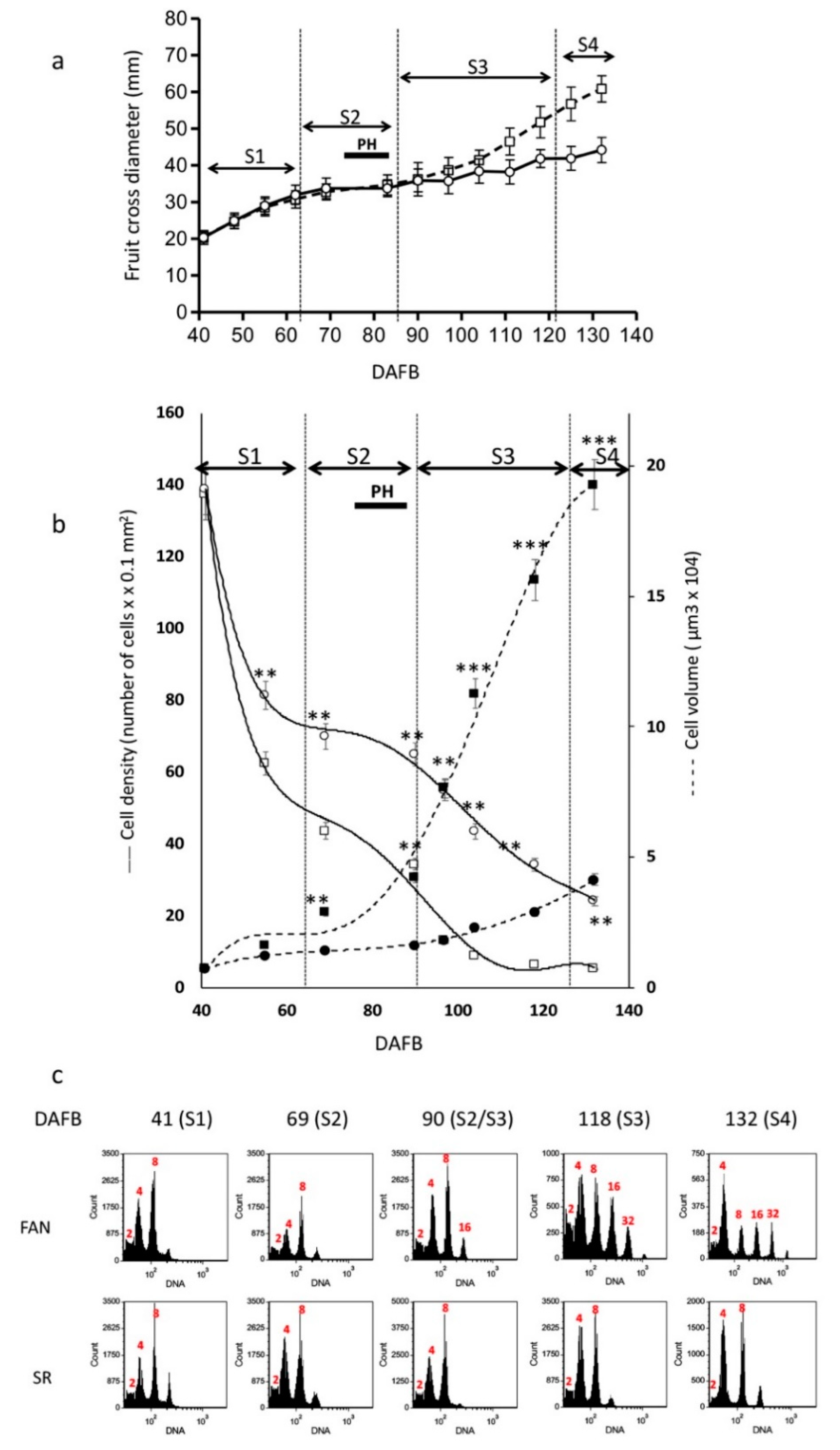{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Samples, and Cytological Analysis
2.2. Ploidy Analysis
2.3. RNA Isolation and qPCR Analysis
2.4. Chromatin Extraction and ChIP Target Analysis
3. Results
3.1. Fruit Growth Pattern in FAN and SR
3.2. Altered Levels of Ploidy Were Observed in SR Mesocarp
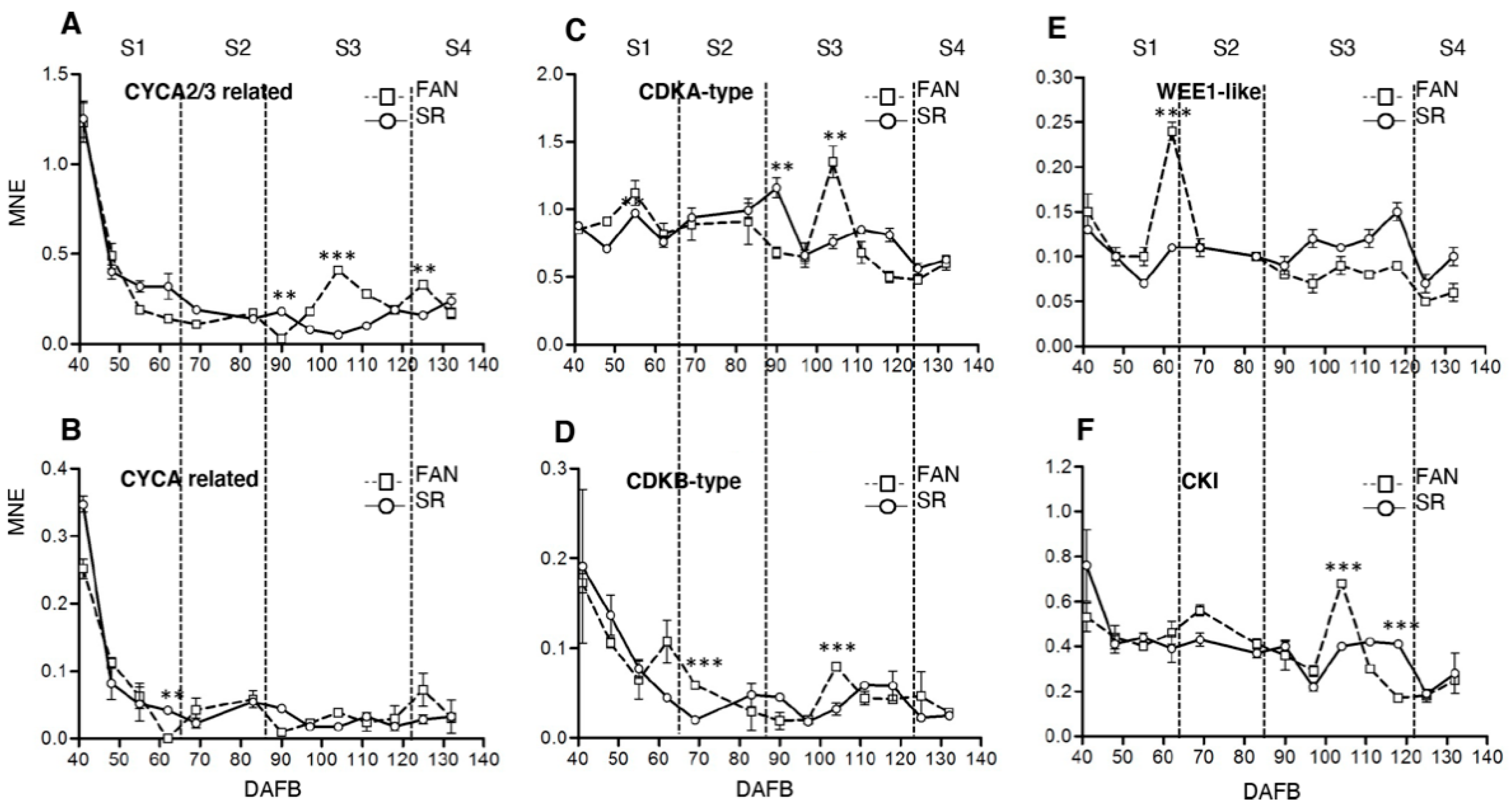
3.3. Expression Level of Cell Cycle and Endoreduplication-Related Genes Is Altered in SR Mesocarp Tissue
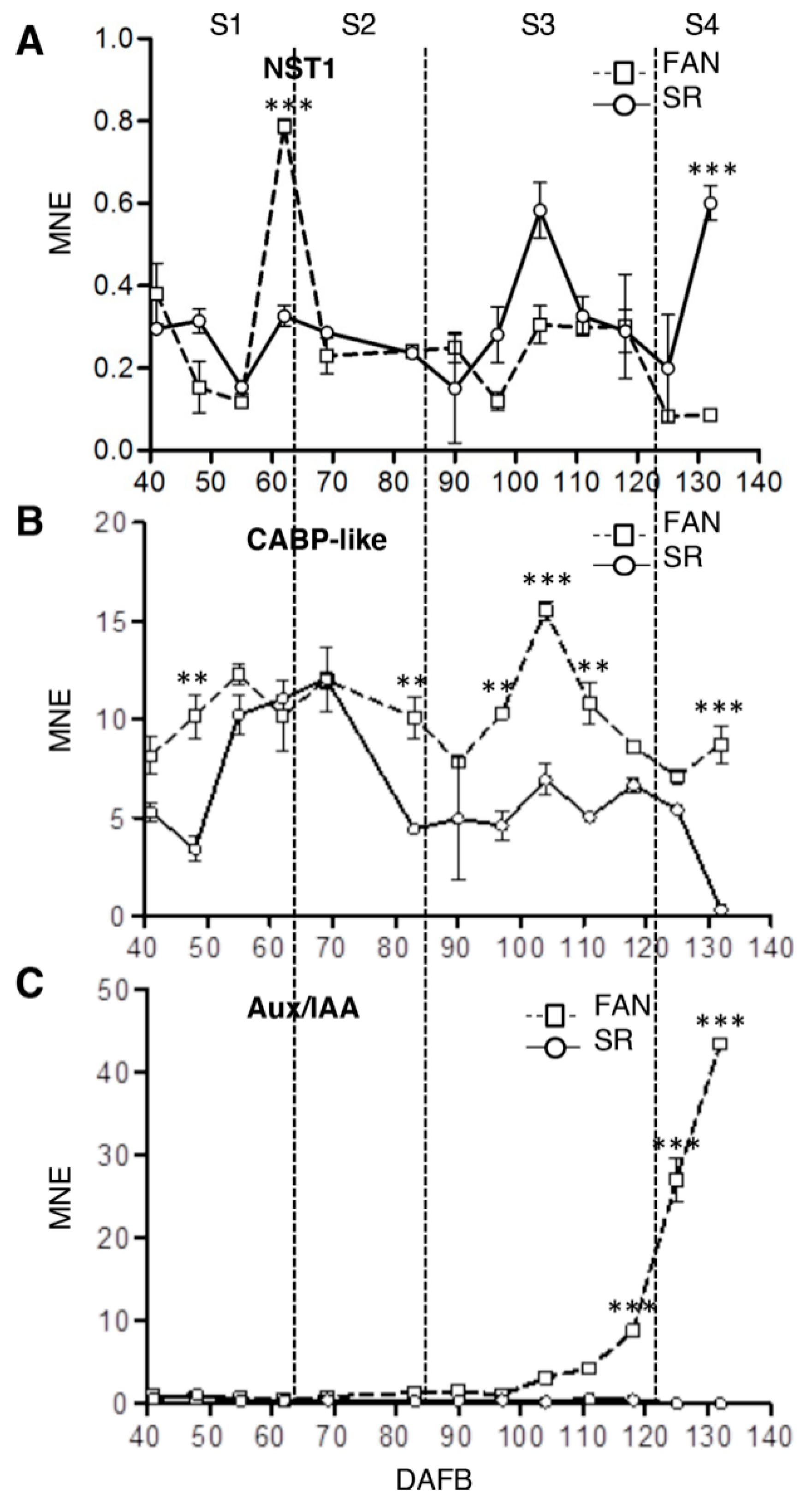
3.4. Altered Expression Level of Developmental Stage and Organ Specific Markers in SR Mesocarp Tissue
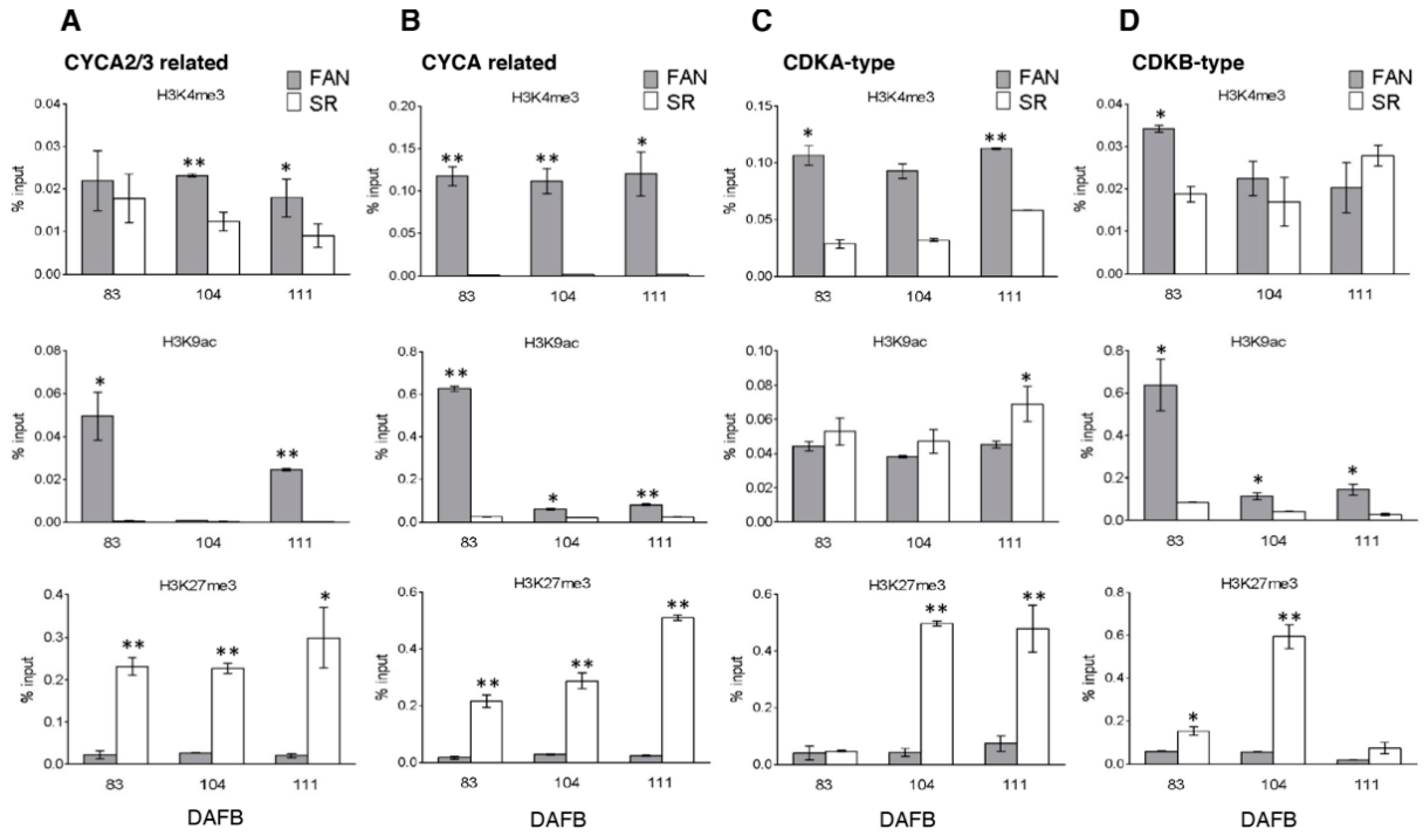
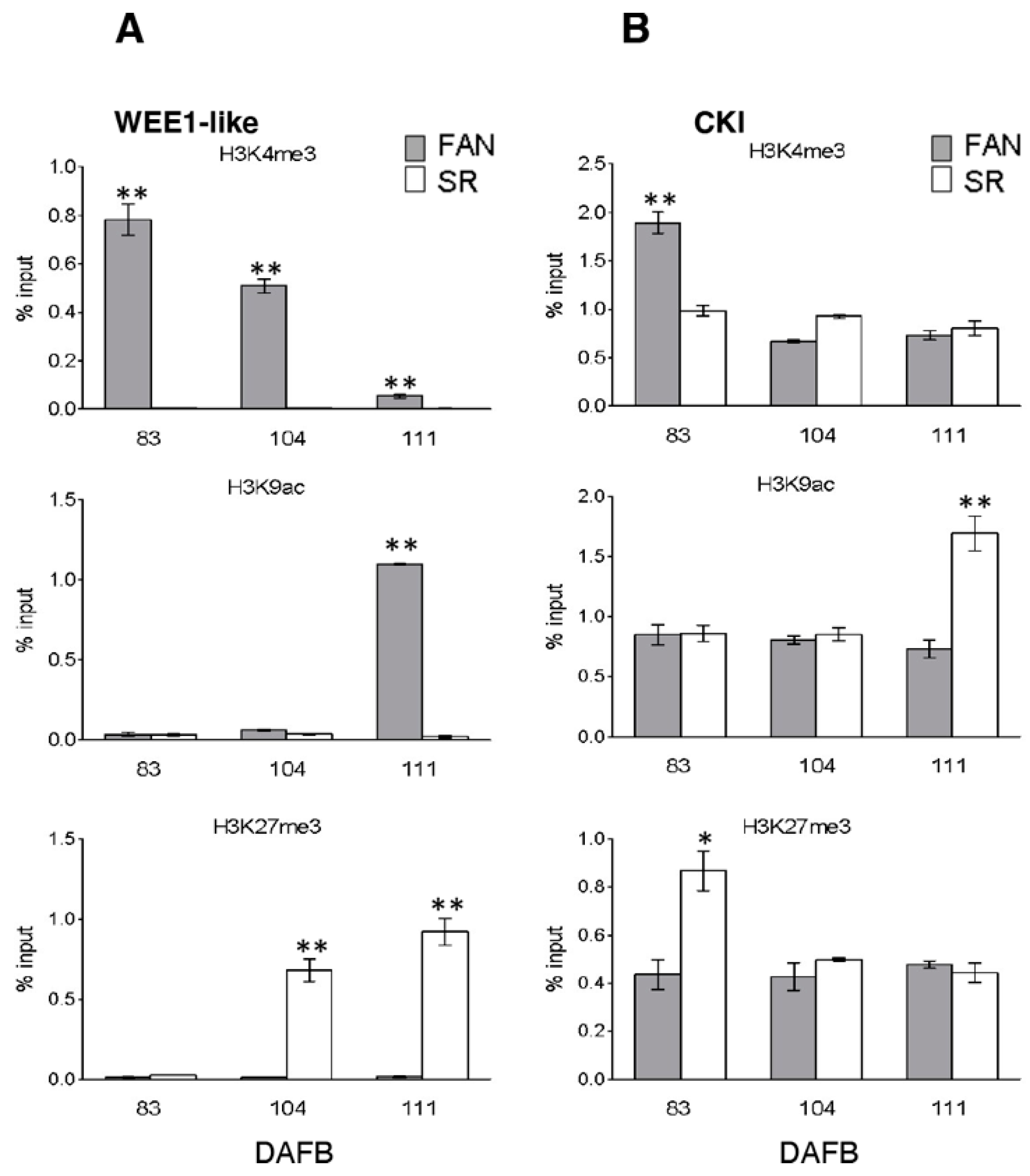
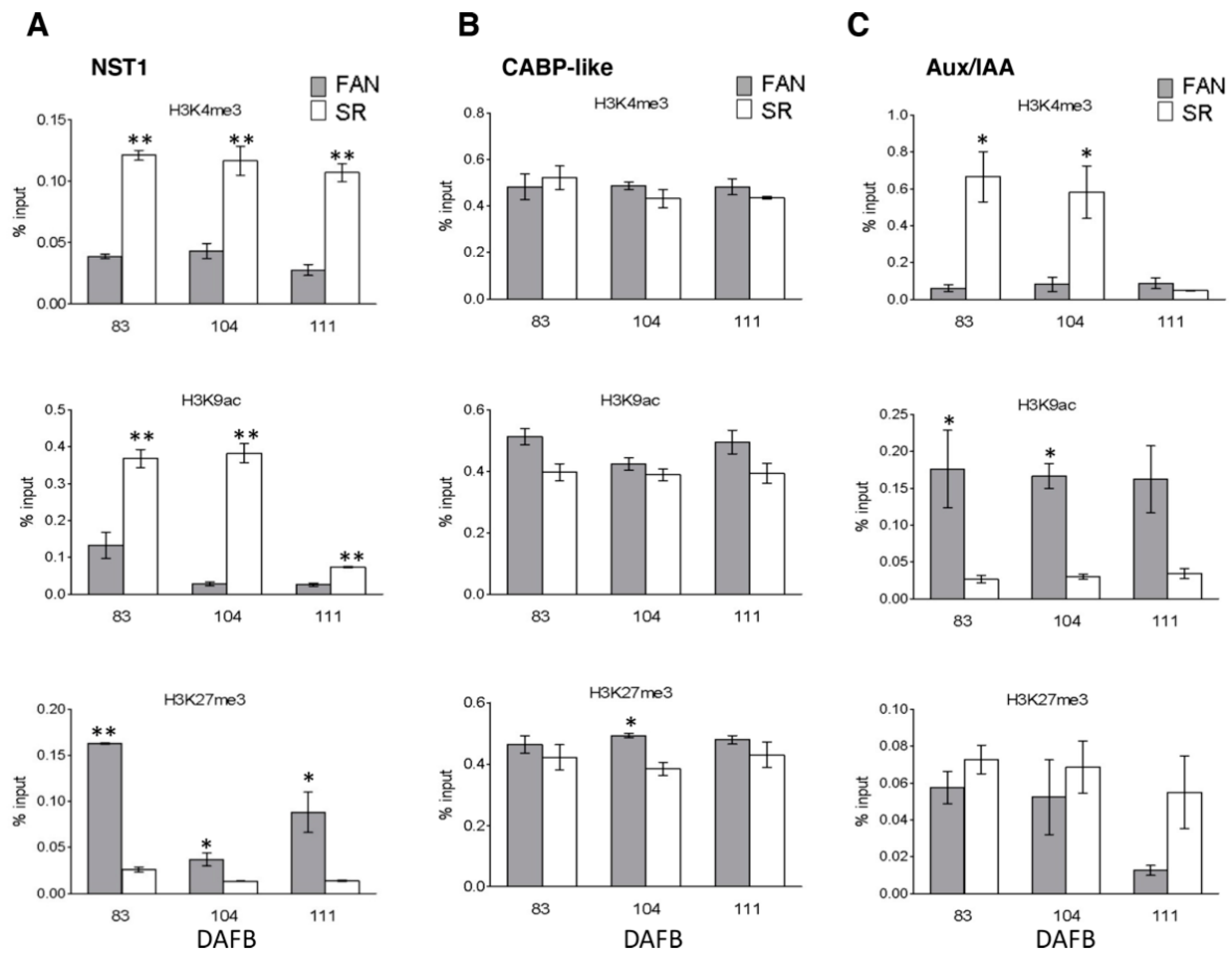
3.5. Differential Enrichments and Depletion of H3K4me3, H3K9ac, and H3K27me3 Modified Histones Were Detectable in Cell Cycle, Endoreduplication-Related and Tissue Marker Gene Loci
4. Discussion
4.1. Endoreduplication Processes Slight Affect Peach Fruits Size
4.2. The Low Ploidy Level in SR Fruit Is Related to an Altered Expression of Genes Controlling the Endocycle Progression
4.3. Is the Altered Transcriptional Control of Cell Cycle and Endocycle Is Related to Uncorrected Fruit Patterning?
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yao, J.-L.; Xu, J.; Tomes, S.; Cui, W.; Luo, Z.; Deng, C.; Ireland, H.S.; Schaffer, R.J.; Gleave, A.P. Ectopic Expression of the PISTILLATA Homologous MdPI Inhibits Fruit Tissue Growth and Changes Fruit Shape in Apple. Plant Direct 2018, 2, e00051. [Google Scholar] [CrossRef]
- Harada, T.; Kurahashi, W.; Yanai, M.; Wakasa, Y.; Satoh, T. Involvement of Cell Proliferation and Cell Enlargement in Increasing the Fruit Size of Malus Species. Sci. Hortic. 2005, 105, 447–456. [Google Scholar] [CrossRef]
- Olmstead, J.W.; Iezzoni, A.F.; Whiting, M.D. Genotypic Differences in Sweet Cherry Fruit Size Are Primarily a Function of Cell Number. J. Am. Soc. Hortic. Sci. 2007, 132, 697–703. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Haji, T.; Miyake, M.; Yaegaki, H. Varietal Differences in Cell Division and Enlargement Periods during Peach (Prunus persica Batsch) Fruit Development. J. Jpn. Soc. Hortic. Sci. 2002, 71, 155–163. [Google Scholar] [CrossRef]
- Scorza, R.; May, L.G.; Purnell, B.; Upchurch, B. Differences in Number and Area of Mesocarp Cells between Small- and Large-Fruited Peach Cultivars. J. Am. Soc. Hortic. Sci. Jashs 1991, 116, 861–864. [Google Scholar] [CrossRef]
- Masia, A.; Zanchin, A.; Rascio, N.; Ramina, A. Some Biochemical and Ultrastructural Aspects of Peach Fruit Development. J. Am. Soc. Hortic. Sci. Jashs 1992, 117, 808–815. [Google Scholar] [CrossRef]
- Bonghi, C.; Ferrarese, L.; Ruperti, B.; Tonutti, P.; Ramina, A. Endo-β-1,4-Glucanases Are Involved in Peach Fruit Growth and Ripening, and Regulated by Ethylene. Physiol. Plant. 1998, 102, 346–352. [Google Scholar] [CrossRef]
- Rodriguez, C.E.; Bustamante, C.A.; Budde, C.O.; Muller, G.L.; Drincovich, M.F.; Lara, M.V. Peach Fruit Development: A Comparative Proteomic Study Between Endocarp and Mesocarp at Very Early Stages Underpins the Main Differential Biochemical Processes Between These Tissues. Front. Plant Sci. 2019, 10, 715. [Google Scholar] [CrossRef] [Green Version]
- Zanchin, A.; Bonghi, C.; Casadoro, G.; Ramina, A.; Rascio, N. Cell Enlargement and Cell Separation During Peach Fruit Development. Int. J. Plant Sci. 1994, 155, 49–56. [Google Scholar] [CrossRef]
- Ohmiya, A. Effects of Auxin on Growth and Ripening of Mesocarp Discs of Peach Fruit. Sci. Hortic. 2000, 84, 309–319. [Google Scholar] [CrossRef]
- Shi, M.; Hu, X.; Wei, Y.; Hou, X.; Yuan, X.; Liu, J.; Liu, Y. Genome-Wide Profiling of Small RNAs and Degradome Revealed Conserved Regulations of MiRNAs on Auxin-Responsive Genes during Fruit Enlargement in Peaches. Int. J. Mol. Sci. 2017, 18, 2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertin, N. Analysis of the Tomato Fruit Growth Response to Temperature and Plant Fruit Load in Relation to Cell Division, Cell Expansion and DNA Endoreduplication. Ann. Bot. 2005, 95, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rewers, M.; Sadowski, J.; Sliwinska, E. Endoreduplication in Cucumber (Cucumis sativus) Seeds during Development, after Processing and Storage, and during Germination. Ann. Appl. Biol. 2009, 155, 431–438. [Google Scholar] [CrossRef]
- Chevalier, C.; Nafati, M.; Mathieu-Rivet, E.; Bourdon, M.; Frangne, N.; Cheniclet, C.; Renaudin, J.-P.; Gevaudant, F.; Hernould, M. Elucidating the Functional Role of Endoreduplication in Tomato Fruit Development. Ann. Bot. 2011, 107, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Bourdon, M.; Frangne, N.; Mathieu-Rivet, E.; Nafati, M.; Cheniclet, C.; Renaudin, J.P.; Chevalier, C. Endoreduplication and growth of fleshy fruits. In Progress in Botany; Springer: Heidelberg, Germany, 2010; Volume 71, pp. 101–132. [Google Scholar]
- Brecht, J.K.; Kader, A.A. Ethylene Production by Fruit of Some Slow-Ripening Nectarine Genotypes. J. Am. Soc. Hortic. Sci. 1984, 109, 763–767. [Google Scholar]
- Brecht, J.K.; Kader, A.A.; Ramming, D.W. Description and Postharvest Physiology of Some Slow-Ripening Nectarine Genotypes. J. Am. Soc. Hortic. Sci. 1984, 109, 596–600. [Google Scholar]
- Ramming, D.W. Genetic Control of a Slow-Ripening Fruit Trait in Nectarine. Can. J. Plant Sci. 1991, 71, 601–603. [Google Scholar] [CrossRef]
- Tataranni, G.; Spada, A.; Pozzi, C.; Bassi, D. AFLP-Based Bulk Segregant Analysis for Tagging the Slow-Ripening Trait in Peach [Prunus persica (L.) Batsch]. J. Hortic. Sci. Biotechnol. 2010, 85, 78–82. [Google Scholar] [CrossRef]
- Giné-Bordonaba, J.; Eduardo, I.; Arús, P.; Cantín, C.M. Biochemical and Genetic Implications of the Slow Ripening Phenotype in Peach Fruit. Sci. Hortic. 2020, 259, 108824. [Google Scholar] [CrossRef]
- Eduardo, I.; Picañol, R.; Rojas, E.; Batlle, I.; Howad, W.; Aranzana, M.J.; Arús, P. Mapping of a Major Gene for the Slow Ripening Character in Peach: Co-Location with the Maturity Date Gene and Development of a Candidate Gene-Based Diagnostic Marker for Its Selection. Euphytica 2015, 205, 627–636. [Google Scholar] [CrossRef]
- Meneses, C.; Ulloa-Zepeda, L.; Cifuentes-Esquivel, A.; Infante, R.; Cantin, C.M.; Batlle, I.; Arús, P.; Eduardo, I. A Codominant Diagnostic Marker for the Slow Ripening Trait in Peach. Mol. Breed. 2016, 36, 77. [Google Scholar] [CrossRef]
- Nuñez-Lillo, G.; Cifuentes-Esquivel, A.; Troggio, M.; Micheletti, D.; Infante, R.; Campos-Vargas, R.; Orellana, A.; Blanco-Herrera, F.; Meneses, C. Identification of Candidate Genes Associated with Mealiness and Maturity Date in Peach [Prunus persica (L.) Batsch] Using QTL Analysis and Deep Sequencing. Tree Genet. Genomes 2015, 11, 86. [Google Scholar] [CrossRef]
- Botton, A.; Rasori, A.; Ziliotto, F.; Moing, A.; Maucourt, M.; Bernillon, S.; Deborde, C.; Petterle, A.; Varotto, S.; Bonghi, C. The Peach HECATE3-like Gene FLESHY Plays a Double Role during Fruit Development. Plant Mol. Biol. 2016, 91, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Zhou, Y.-H.; Shu, W.-S.; Cheng, H.-Y.; Wang, L.; Han, Y.-P.; Zhang, Y.-Y.; Yu, M.-L.; Joldersma, D.; Zhang, S.-L. RNA-Seq Analysis Unveils Gene Regulation of Fruit Size Cooperatively Determined by Velocity and Duration of Fruit Swelling in Peach. Physiol. Plant. 2018, 164, 320–336. [Google Scholar] [CrossRef]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular Genetics of Blood-Fleshed Peach Reveals Activation of Anthocyanin Biosynthesis by NAC Transcription Factors. Plant J. Cell Mol. Biol. 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Dirlewanger, E.; Moing, A.; Rothan, C.; Svanella, L.; Pronier, V.; Guye, A.; Plomion, C.; Monet, R. Mapping QTLs Controlling Fruit Quality in Peach (Prunus persica (L.) Batsch). Theor. Appl. Genet. 1999, 98, 18–31. [Google Scholar] [CrossRef]
- Quilot, B.; Wu, B.H.; Kervella, J.; Genard, M.; Foulongne, M.; Moreau, K. QTL Analysis of Quality Traits in an Advanced Backcross between Prunus persica Cultivars and the Wild Relative Species P. davidiana. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2004, 109, 884–897. [Google Scholar] [CrossRef]
- Martinez-Garcia, P.J.; Parfitt, D.E.; Bostock, R.M.; Fresnedo-Ramirez, J.; Vazquez-Lobo, A.; Ogundiwin, E.A.; Gradziel, T.M.; Crisosto, C.H. Application of Genomic and Quantitative Genetic Tools to Identify Candidate Resistance Genes for Brown Rot Resistance in Peach. PLoS ONE 2013, 8, e78634. [Google Scholar] [CrossRef]
- Zeballos, J.L.; Abidi, W.; Giménez, R.; Monforte, A.J.; Moreno, M.Á.; Gogorcena, Y. Mapping QTLs Associated with Fruit Quality Traits in Peach [Prunus persica (L.) Batsch] Using SNP Maps. Tree Genet. Genomes 2016, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Marshall, W.F.; Young, K.D.; Swaffer, M.; Wood, E.; Nurse, P.; Kimura, A.; Frankel, J.; Wallingford, J.; Walbot, V.; Qu, X.; et al. What Determines Cell Size? BMC Biol. 2012, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Ferrandiz, C.; Yanofsky, M.F.; Martienssen, R. The FRUITFULL MADS-Box Gene Mediates Cell Differentiation during Arabidopsis Fruit Development. Dev. Camb. Engl. 1998, 125, 1509–1517. [Google Scholar]
- Kim, Y.-S.; Kim, S.-G.; Park, J.-E.; Park, H.-Y.; Lim, M.-H.; Chua, N.-H.; Park, C.-M. A Membrane-Bound NAC Transcription Factor Regulates Cell Division in Arabidopsis. Plant Cell 2006, 18, 3132–3144. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Potuschak, T.; Colon-Carmona, A.; Gutierrez, R.A.; Doerner, P. Arabidopsis TCP20 Links Regulation of Growth and Cell Division Control Pathways. Proc. Natl. Acad. Sci. USA 2005, 102, 12978–12983. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, J.; Sakai, T.; Ishida, S.; Yamaguchi, I.; Kamiya, Y.; Takahashi, Y. Repression of Shoot Growth, a BZIP Transcriptional Activator, Regulates Cell Elongation by Controlling the Level of Gibberellins. Plant Cell 2000, 12, 901–915. [Google Scholar] [CrossRef] [Green Version]
- Nibau, C.; Gibbs, D.J.; Bunting, K.A.; Moody, L.A.; Smiles, E.J.; Tubby, J.A.; Bradshaw, S.J.; Coates, J.C. ARABIDILLO Proteins Have a Novel and Conserved Domain Structure Important for the Regulation of Their Stability. Plant Mol. Biol. 2011, 75, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.M.; Henikoff, S. Histone Variants: Dynamic Punctuation in Transcription. Genes Dev. 2014, 28, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Catarino, R.R.; Stark, A. Assessing Sufficiency and Necessity of Enhancer Activities for Gene Expression and the Mechanisms of Transcription Activation. Genes Dev. 2018, 32, 202–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, F.; Alves, E.; Pio, R.; Castro, E.; Reighard, G.; Freire, A.I.; Mayer, N.A.; Pimentel, R. Influence of Temperature on the Development of Peach Fruit in a Subtropical Climate Region. Agronomy 2019, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Ziliotto, F.; Corso, M.; Rizzini, F.M.; Rasori, A.; Botton, A.; Bonghi, C. Grape Berry Ripening Delay Induced by a Pre-Veraison NAA Treatment Is Paralleled by a Shift in the Expression Pattern of Auxin- and Ethylene- Related Genes. BMC Plant Biol. 2012, 12, 185. [Google Scholar] [CrossRef] [Green Version]
- Giulia, E.; Alessandro, B.; Mariano, D.; Andrea, B.; Benedetto, R.; Angelo, R. Early Induction of Apple Fruitlet Abscission Is Characterized by an Increase of Both Isoprene Emission and Abscisic Acid Content. Plant Physiol. 2013, 161, 1952–1969. [Google Scholar] [CrossRef] [Green Version]
- Simon, P. Q-Gene: Processing Quantitative Real-Time RT-PCR Data. Bioinform. Oxf. Engl. 2003, 19, 1439–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of Reliable Reference Genes for Gene Expression Studies in Peach Using Real-Time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Bonghi, C.; Trainotti, L.; Botton, A.; Tadiello, A.; Rasori, A.; Ziliotto, F.; Zaffalon, V.; Casadoro, G.; Ramina, A. A Microarray Approach to Identify Genes Involved in Seed-Pericarp Cross- Talk and Development in Peach. BMC Plant Biol. 2011, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper--Excel-Based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Rossi, V.; Locatelli, S.; Varotto, S.; Donn, G.; Pirona, R.; Henderson, D.A.; Hartings, H.; Motto, M. Maize Histone Deacetylase Hda101 Is Involved in Plant Development, Gene Transcription, and Sequence-Specific Modulation of Histone Modification of Genes and Repeats. Plant Cell 2007, 19, 1145–1162. [Google Scholar] [CrossRef] [Green Version]
- Tonutti, P.; Bonghi, C.; Ruperti, B.; Tornielli, G.B.; Ramina, A. Ethylene Evolution and 1-Aminocyclopropane-1-Carboxylate Oxidase Gene Expression during Early Development and Ripening of Peach Fruit. J. Am. Soc. Hortic. Sci. Jashs 1997, 122, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Malladi, A.; Hirst, P.M. Increase in Fruit Size of a Spontaneous Mutant of “Gala” Apple (Malus x Domestica Borkh.) Is Facilitated by Altered Cell Production and Enhanced Cell Size. J. Exp. Bot. 2010, 61, 3003–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Liu, C.; Song, L.; Li, Y.; Li, M. PaCYP78A9, a Cytochrome P450, Regulates Fruit Size in Sweet Cherry (Prunus avium L.). Front. Plant Sci. 2017, 8, 2076. [Google Scholar] [CrossRef] [Green Version]
- Dardick, C.D.; Callahan, A.M.; Chiozzotto, R.; Schaffer, R.J.; Piagnani, M.C.; Scorza, R. Stone Formation in Peach Fruit Exhibits Spatial Coordination of the Lignin and Flavonoid Pathways and Similarity to Arabidopsis Dehiscence. BMC Biol. 2010, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Farinati, S.; Rasori, A.; Varotto, S.; Bonghi, C. Rosaceae Fruit Development, Ripening and Post-Harvest: An Epigenetic Perspective. Front. Plant Sci. 2017, 8, 1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forestan, C.; (University of Bologna, Bologna, Italy); Varotto, S.; (University of Padova, Padova, Italy); Bonghi, C.; (University of Padova, Padova, Italy). Personal communication, 2019.
- Chevalier, C.; Bourdon, M.; Pirrello, J.; Cheniclet, C.; Gevaudant, F.; Frangne, N. Endoreduplication and Fruit Growth in Tomato: Evidence in Favour of the Karyoplasmic Ratio Theory. J. Exp. Bot. 2014, 65, 2731–2746. [Google Scholar] [CrossRef] [Green Version]
- Nilo-Poyanco, R.; Moraga, C.; Benedetto, G.; Orellana, A.; Almeida, A.M. Shotgun Proteomics of Peach Fruit Reveals Major Metabolic Pathways Associated to Ripening. BMC Genom. 2021, 22, 17. [Google Scholar] [CrossRef]
- Musseau, C.; Jorly, J.; Gadin, S.; Sorensen, I.; Deborde, C.; Bernillon, S.; Mauxion, J.-P.; Atienza, I.; Moing, A.; Lemaire-Chamley, M.; et al. The Tomato Guanylate-Binding Protein SlGBP1 Enables Fruit Tissue Differentiation by Maintaining Endopolyploid Cells in a Non-Proliferative State. Plant Cell 2020, 32, 3188–3205. [Google Scholar] [CrossRef]
- Cheniclet, C.; Rong, W.Y.; Causse, M.; Frangne, N.; Bolling, L.; Carde, J.-P.; Renaudin, J.-P. Cell Expansion and Endoreduplication Show a Large Genetic Variability in Pericarp and Contribute Strongly to Tomato Fruit Growth. Plant Physiol. 2005, 139, 1984–1994. [Google Scholar] [CrossRef] [Green Version]
- Verkest, A.; Weinl, C.; Inze, D.; De Veylder, L.; Schnittger, A. Switching the Cell Cycle. Kip-Related Proteins in Plant Cell Cycle Control. Plant Physiol. 2005, 139, 1099–1106. [Google Scholar] [CrossRef] [Green Version]
- Lees, E. Cyclin Dependent Kinase Regulation. Curr. Opin. Cell Biol. 1995, 7, 773–780. [Google Scholar] [CrossRef]
- Berry, L.D.; Gould, K.L. Novel Alleles of Cdc13 and Cdc2 Isolated as Suppressors of Mitotic Catastrophe in Schizosaccharomyces Pombe. Mol. Gen. Genet. MGG 1996, 251, 635–646. [Google Scholar] [CrossRef]
- Gonzalez, N.; Gevaudant, F.; Hernould, M.; Chevalier, C.; Mouras, A. The Cell Cycle-Associated Protein Kinase WEE1 Regulates Cell Size in Relation to Endoreduplication in Developing Tomato Fruit. Plant J. Cell Mol. Biol. 2007, 51, 642–655. [Google Scholar] [CrossRef]
- Desvoyes, B.; Fernandez-Marcos, M.; Sequeira-Mendes, J.; Otero, S.; Vergara, Z.; Gutierrez, C. Looking at Plant Cell Cycle from the Chromatin Window. Front. Plant Sci. 2014, 5, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, H.; Wildhaber, T.; Siretskiy, A.; Gruissem, W.; Hennig, L. Distinct Modes of DNA Accessibility in Plant Chromatin. Nat. Commun. 2012, 3, 1281. [Google Scholar] [CrossRef] [Green Version]
- Tsukaya, H. Re-Examination of the Role of Endoreduplication on Cell-Size Control in Leaves. J. Plant Res. 2019, 132, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Mitsuda, N.; Ohme-Takagi, M. NAC Transcription Factors NST1 and NST3 Regulate Pod Shattering in a Partially Redundant Manner by Promoting Secondary Wall Formation after the Establishment of Tissue Identity. Plant J. Cell Mol. Biol. 2008, 56, 768–778. [Google Scholar] [CrossRef]
- Dardick, C.; Callahan, A.M. Evolution of the Fruit Endocarp: Molecular Mechanisms Underlying Adaptations in Seed Protection and Dispersal Strategies. Front. Plant Sci. 2014, 5, 284. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farinati, S.; Forestan, C.; Canton, M.; Galla, G.; Bonghi, C.; Varotto, S. Regulation of Fruit Growth in a Peach Slow Ripening Phenotype. Genes 2021, 12, 482. https://doi.org/10.3390/genes12040482
Farinati S, Forestan C, Canton M, Galla G, Bonghi C, Varotto S. Regulation of Fruit Growth in a Peach Slow Ripening Phenotype. Genes. 2021; 12(4):482. https://doi.org/10.3390/genes12040482
Chicago/Turabian StyleFarinati, Silvia, Cristian Forestan, Monica Canton, Giulio Galla, Claudio Bonghi, and Serena Varotto. 2021. "Regulation of Fruit Growth in a Peach Slow Ripening Phenotype" Genes 12, no. 4: 482. https://doi.org/10.3390/genes12040482
APA StyleFarinati, S., Forestan, C., Canton, M., Galla, G., Bonghi, C., & Varotto, S. (2021). Regulation of Fruit Growth in a Peach Slow Ripening Phenotype. Genes, 12(4), 482. https://doi.org/10.3390/genes12040482