
Nutritional Regulation of Mammary Gland Development and Milk Synthesis in Animal Models and Dairy Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
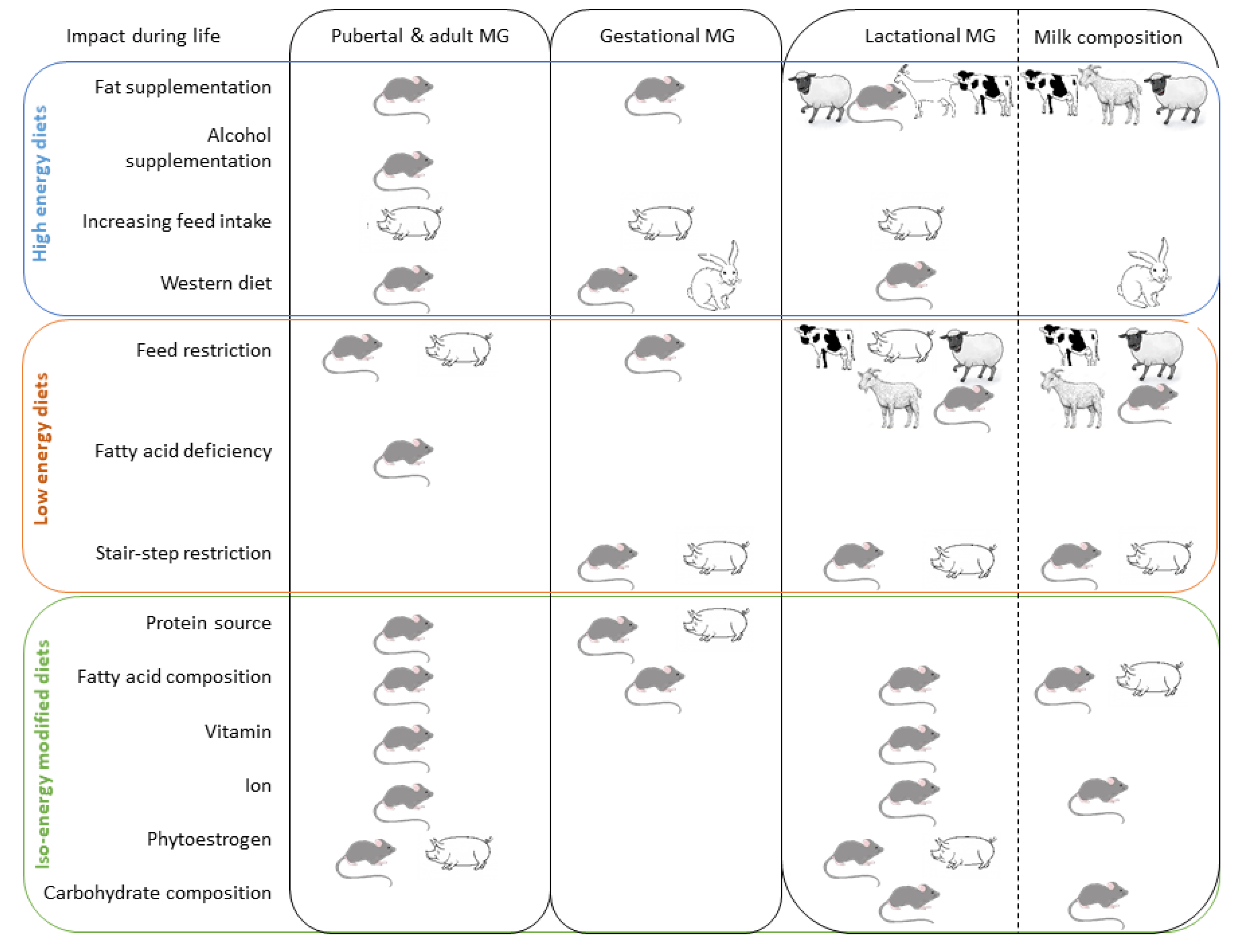
2. Nutritional Regulation of the Mammary Gland Development
3. Effects of Energetic Supplementation
4. Effects of Energetic Restriction
5. Effects of Isoenergetic Modifications
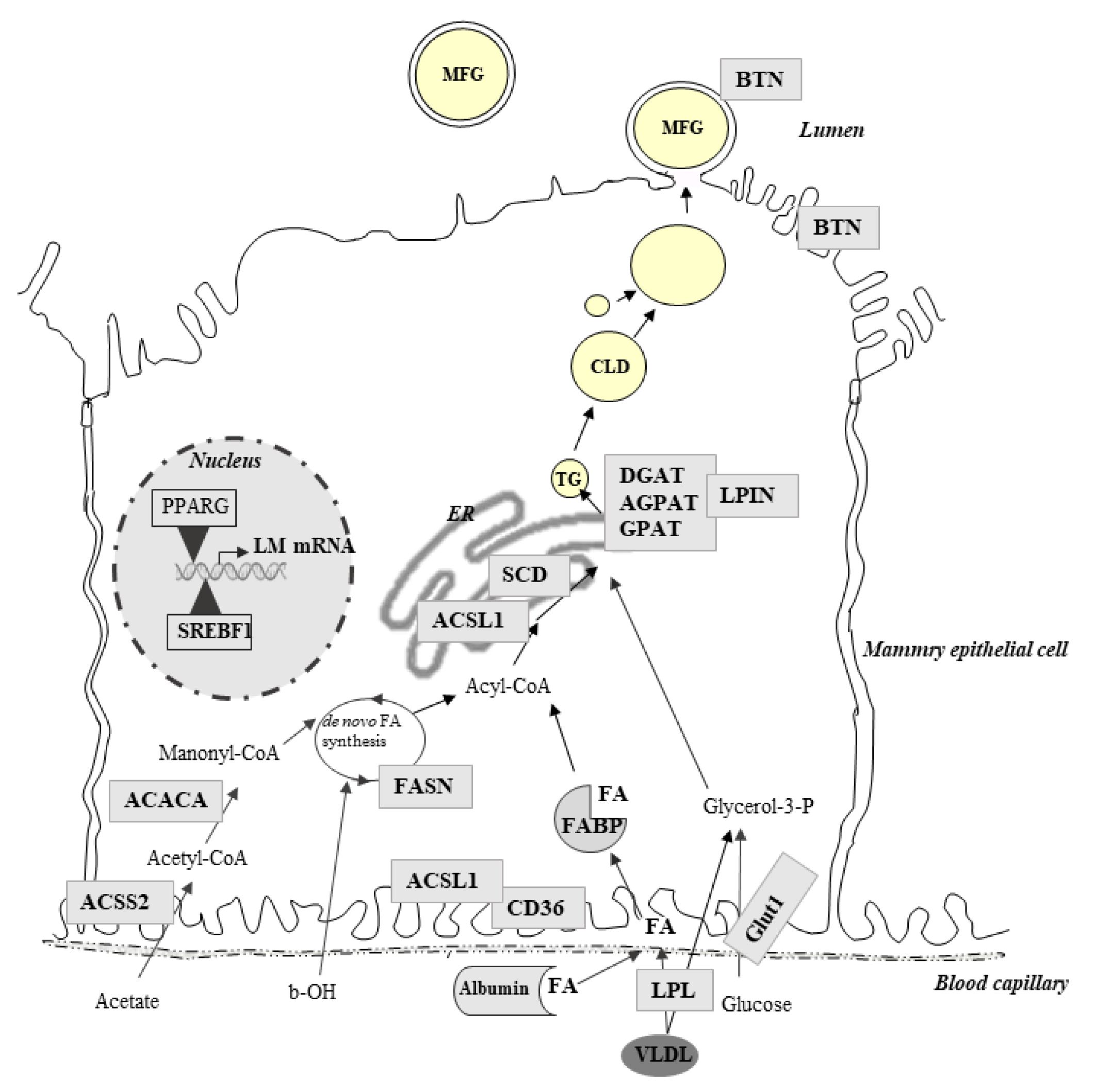
6. Nutritional Regulation of Gene Expression Involved in Milk Component Synthesis in Ruminants
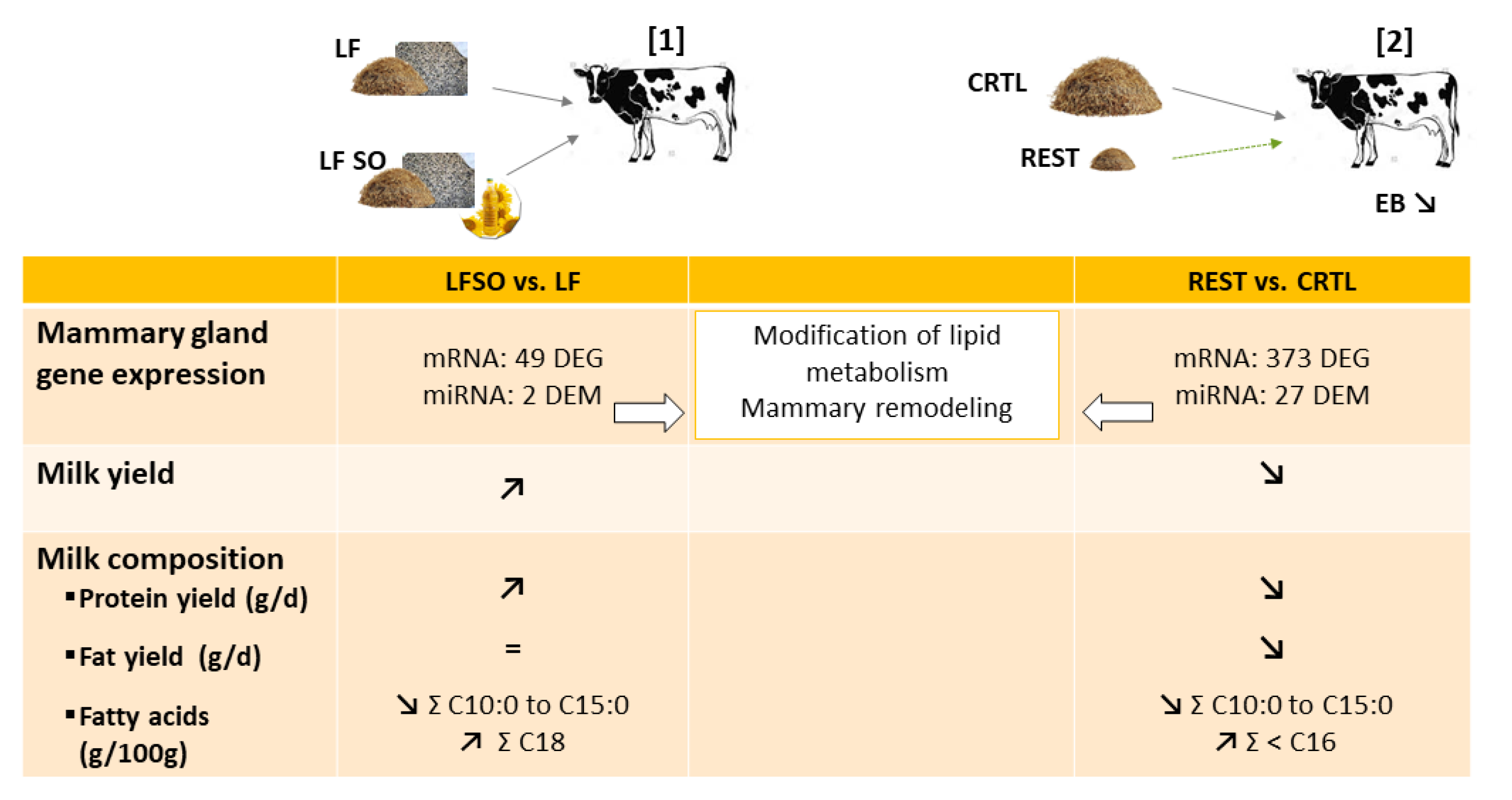
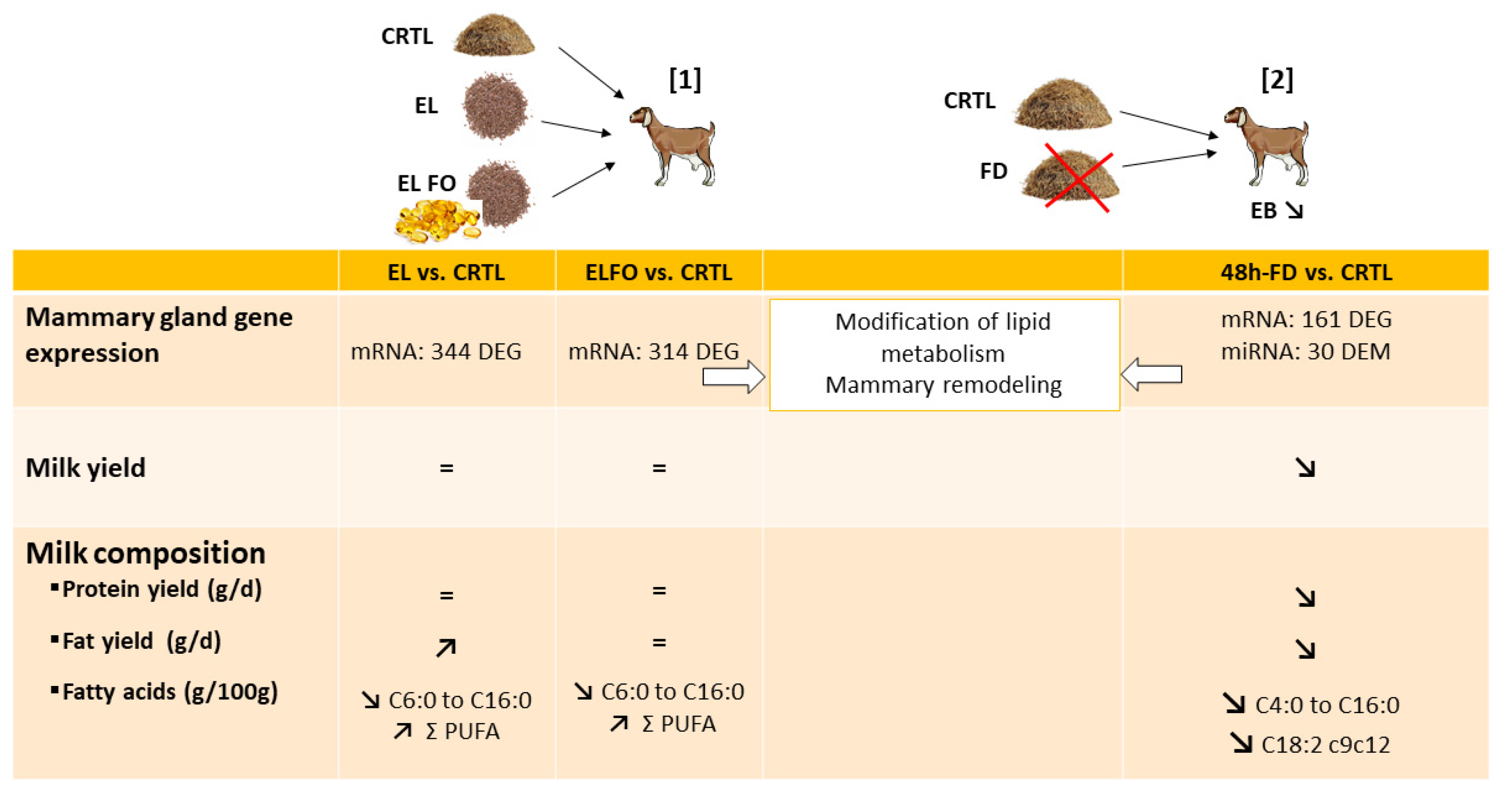
7. Effects of Lipid-Supplemented Diets on Ruminant Mammary Gene Expression
8. Effects of Feed Restriction and Deprivation on Ruminant Mammary Gene Expression
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| (CLA) | conjugated linoleic acid |
| (DHA) | docosahexaenoic acid |
| (FA) | fatty acid |
| (MG) | Mammary Gland |
| (miRNA) | microRNA |
| (RNA-seq) | RNA sequencing |
References
- Hurley, W.L. Review: Mammary gland development in swine: Embryo to early lactation. Animal 2019, 13, s11–s19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.; Le Guillou, S.; Hue-Beauvais, C.; Le Provost, F. Epigenetics: New Insights into Mammary Gland Biology. Genes 2021, 12, 231. [Google Scholar] [CrossRef]
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinde, K. Richer milk for sons but more milk for daughters: Sex-biased investment during lactation varies with maternal life history in rhesus macaques. Am. J. Hum. Biol. 2009, 21, 512–519. [Google Scholar] [CrossRef]
- McNally, S.; Stein, T. Overview of Mammary Gland Development: A Comparison of Mouse and Human. Methods Mol. Biol. 2017, 1501, 1–17. [Google Scholar] [CrossRef]
- Hughes, K. Comparative mammary gland postnatal development and tumourigenesis in the sheep, cow, cat and rabbit: Exploring the menagerie. Semin. Cell Dev. Biol. 2020. [Google Scholar] [CrossRef]
- Givens, D.I. Milk and meat in our diet: Good or bad for health? Animal 2010, 4, 1941–1952. [Google Scholar] [CrossRef] [Green Version]
- German, J.B.; Dillard, C.J. Saturated fats: A perspective from lactation and milk composition. Lipids 2010, 45, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Tech. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Hennighausen, L.R.G. Think globally, act locally: The making of a mouse mammary gland. Genes Dev. 1998, 12, 449–455. [Google Scholar] [CrossRef] [Green Version]
- McNally, S.; Martin, F. Molecular regulators of pubertal mammary gland development. Ann. Med. 2011, 43, 212–234. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.K.; Govindarajah, V.; Cheong, A.; Veevers, J.; Song, D.; Gear, R.; Zhu, X.; Ying, J.; Kendler, A.; Medvedovic, M.; et al. Gestational high-fat diet and bisphenol A exposure heightens mammary cancer risk. Endocr. Relat. Cancer 2017, 24, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, S.; Yan, L. Dietary Supplementation with Methylseleninic Acid Inhibits Mammary Tumorigenesis and Metastasis in Male MMTV-PyMT Mice. Biol. Trace Elem. Res. 2018, 184, 186–195. [Google Scholar] [CrossRef]
- Da Silva-Santi, L.G.; Antunes, M.M.; Caparroz-Assef, S.M.; Carbonera, F.; Masi, L.N.; Curi, R.; Visentainer, J.V.; Bazotte, R.B. Liver Fatty Acid Composition and Inflammation in Mice Fed with High-Carbohydrate Diet or High-Fat Diet. Nutrients 2016, 8, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Ren, H.; Cao, Y.; Wang, Y.; Huo, G. Low dietary protein and high carbohydrate infant formula affects the microbial ecology of the large intestine in neonatal rats. Can. J. Microbiol. 2017, 63, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Nazni, P. Association of western diet & lifestyle with decreased fertility. Indian J. Med. Res. 2014, 140, S78–S81. [Google Scholar] [PubMed]
- Myles, I.A. Fast food fever: Reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Andrade Fde, O.; De Assis, S.; Jin, L.; Fontelles, C.C.; Barbisan, L.F.; Purgatto, E.; Hilakivi-Clarke, L.; Ong, T.P. Lipidomic fatty acid profile and global gene expression pattern in mammary gland of rats that were exposed to lard-based high fat diet during fetal and lactation periods associated to breast cancer risk in adulthood. Chem. Biol. Interact 2015, 239, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Farmer, C. Nutritional impact on mammary development in pigs: A review. J. Anim. Sci. 2018, 96, 3748–3756. [Google Scholar] [CrossRef] [Green Version]
- Farmer, C.; Palin, M.F.; Martel-Kennes, Y. Impact of diet deprivation and subsequent over-allowance during prepuberty. Part 1. Effects on growth performance, metabolite status, and mammary gland development in gilts. J. Anim. Sci. 2012, 90, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Farmer, C.; Petitclerc, D.; Sorensen, M.T.; Vignola, M.; Dourmad, J.Y. Impacts of dietary protein level and feed restriction during prepuberty on mammogenesis in gilts. J. Anim. Sci. 2004, 82, 2343–2351. [Google Scholar] [CrossRef]
- Kim, S.W.; Hurley, W.L.; Han, I.K.; Stein, H.H.; Easter, R.A. Effect of nutrient intake on mammary gland growth in lactating sows. J. Anim. Sci. 1999, 77, 3304–3315. [Google Scholar] [CrossRef] [PubMed]
- Olson, L.K.; Tan, Y.; Zhao, Y.; Aupperlee, M.D.; Haslam, S.Z. Pubertal exposure to high fat diet causes mouse strain-dependent alterations in mammary gland development and estrogen responsiveness. Int. J. Obes. 2010, 34, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamikawa, A.; Ichii, O.; Yamaji, D.; Imao, T.; Suzuki, C.; Okamatsu-Ogura, Y.; Terao, A.; Kon, Y.; Kimura, K. Diet-induced obesity disrupts ductal development in the mammary glands of nonpregnant mice. Dev. Dyn. 2009, 238, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Juneja, S.C.; Maihle, N.J.; Cleary, M.P. Leptin--a growth factor in normal and malignant breast cells and for normal mammary gland development. J. Natl. Cancer Inst. 2002, 94, 1704–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, L.L.; Grayson, B.E.; Yadav, E.; Seeley, R.J.; Horseman, N.D. High fat diet alters lactation outcomes: Possible involvement of inflammatory and serotonergic pathways. PloS ONE 2012, 7, e32598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.A.; Li, W.; Hernandez, L.L. Effect of high-fat diet feeding and associated transcriptome changes in the peak lactation mammary gland in C57BL/6 dams. Physiol. Genom. 2018, 50, 1059–1070. [Google Scholar] [CrossRef] [Green Version]
- Hue-Beauvais, C.; Chavatte-Palmer, P.; Aujean, E.; Dahirel, M.; Laigre, P.; Pechoux, C.; Bouet, S.; Devinoy, E.; Charlier, M. An obesogenic diet started before puberty leads to abnormal mammary gland development during pregnancy in the rabbit. Dev. Dyn. 2011, 240, 347–356. [Google Scholar] [CrossRef]
- Hue-Beauvais, C.; Miranda, G.; Aujean, E.; Jaffrezic, F.; Devinoy, E.; Martin, P.; Charlier, M. Diet-induced modifications to milk composition have long-term effects on offspring growth in rabbits. J. Anim. Sci. 2017, 95, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Hue-Beauvais, C.; Laubier, J.; Brun, N.; Houtia, I.; Jaffrezic, F.; Bevilacqua, C.; Le Provost, F.; Charlier, M. Puberty is a critical window for the impact of diet on mammary gland development in the rabbit. Dev. Dyn. 2019. [Google Scholar] [CrossRef] [Green Version]
- Sejrsen, K. Relationships between nutrition, puberty and mammary development in cattle. Proc. Nutr. Soc. 1994, 53, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Flint, D.J.; Travers, M.T.; Barber, M.C.; Binart, N.; Kelly, P.A. Diet-induced obesity impairs mammary development and lactogenesis in murine mammary gland. Am. J. Physiol. 2005, 288, E1179–E1187. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.O.; Clark, J.A.; Myers, P.H.; Korach, K.S. Mammary gland development in adult mice requires epithelial and stromal estrogen receptor α. Endocrinology 2002, 143, 2357–2365. [Google Scholar] [CrossRef]
- Dekker, L.V.; Parker, P.J. Protein kinase C--a question of specificity. Trends Biochem. Sci. 1994, 19, 73–77. [Google Scholar] [CrossRef]
- Kris, E.S.; Choe, M.; Luthra, R.; Conway, H.; Barnett, T.; Yaktine, A.; Birt, D.F. Protein kinase C activity is reduced in epidermal cells from energy-restricted SENCAR mice. J. Nutr. 1994, 124, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilakivi-Clarke, L.; Stoica, A.; Raygada, M.; Martin, M.B. Consumption of a high-fat diet alters estrogen receptor content, protein kinase C activity, and mammary gland morphology in virgin and pregnant mice and female offspring. Cancer Res. 1998, 58, 654–660. [Google Scholar]
- Silva, D.C.; Lima-Leopoldo, A.P.; Leopoldo, A.S.; Campos, D.H.; Nascimento, A.F.; Oliveira Junior, S.A.; Padovani, C.R.; Cicogna, A.C. Influence of term of exposure to high-fat diet-induced obesity on myocardial collagen type I and III. Arq. Bras. Cardiol. 2014, 102, 157–163. [Google Scholar] [CrossRef]
- Ishimoto, T.; Lanaspa, M.A.; Rivard, C.J.; Roncal-Jimenez, C.A.; Orlicky, D.J.; Cicerchi, C.; McMahan, R.H.; Abdelmalek, M.F.; Rosen, H.R.; Jackman, M.R.; et al. High-fat and high-sucrose (western) diet induces steatohepatitis that is dependent on fructokinase. Hepatology 2013, 58, 1632–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, S.; Li, C.; Gao, T.; Liu, Y.; Rangiani, A.; Sun, Y.; Hao, J.; George, A.; Lu, Y.; et al. Inactivation of a novel FGF23 regulator, FAM20C, leads to hypophosphatemic rickets in mice. Plos Genet. 2012, 8, e1002708. [Google Scholar] [CrossRef]
- Singh, K.; Hartley, D.G.; McFadden, T.B.; Mackenzie, D.D. Dietary fat regulates mammary stearoyl coA desaturase expression and activity in lactating mice. J. Dairy Res. 2004, 71, 1–6. [Google Scholar] [CrossRef]
- Butruille, L.; Marousez, L.; Pourpe, C.; Oger, F.; Lecoutre, S.; Catheline, D.; Görs, S.; Metges, C.C.; Guinez, C.; Laborie, C.; et al. Maternal high-fat diet during suckling programs visceral adiposity and epigenetic regulation of adipose tissue stearoyl-CoA desaturase-1 in offspring. Int. J. Obes. 2019, 43, 2381–2393. [Google Scholar] [CrossRef]
- Newmark, H.L.; Lipkin, M.; Maheshwari, N. Colonic hyperproliferation induced in rats and mice by nutritional-stress diets containing four components of a human Western-style diet (series 2). Am. J. Clin. Nutr. 1991, 54, 209s–214s. [Google Scholar] [CrossRef]
- Kurihara, N.; Fan, K.; Thaler, H.T.; Yang, K.; Lipkin, M. Effect of a Western-style diet fortified with increased calcium and vitamin D on mammary gland of C57BL/6 mice. J. Med. Food 2008, 11, 201–206. [Google Scholar] [CrossRef]
- Medina, D. Preneoplastic lesions in murine mammary cancer. Cancer Res. 1976, 36, 2589–2595. [Google Scholar]
- Zhang, L.; Bird, R.P.; Bruce, W.R. Proliferative activity of murine mammary epithelium as affected by dietary fat and calcium. Cancer Res. 1987, 47, 4905–4908. [Google Scholar]
- Lok, E.; Ratnayake, W.M.; Scott, F.W.; Mongeau, R.; Fernie, S.; Nera, E.A.; Malcolm, S.; McMullen, E.; Jee, P.; Clayson, D.B. Effect of varying the type of fat in a semi-purified AIN-76A diet on cellular proliferation in the mammary gland and intestinal crypts in female Swiss Webster mice. Carcinogenesis 1992, 13, 1735–1741. [Google Scholar] [CrossRef]
- Xue, L.; Newmark, H.; Yang, K.; Lipkin, M. Model of mouse mammary gland hyperproliferation and hyperplasia induced by a western-style diet. Nutr. Cancer 1996, 26, 281–287. [Google Scholar] [CrossRef]
- Xue, L.; Lipkin, M.; Newmark, H.; Wang, J. Influence of dietary calcium and vitamin D on diet-induced epithelial cell hyperproliferation in mice. J. Natl. Cancer Inst. 1999, 91, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Grassi, T.F.; Bidinotto, L.T.; Lopes, G.A.D.; Zapaterini, J.R.; Rodrigues, M.A.M.; Barbisan, L.F. Maternal western-style diet enhances the effects of chemically-induced mammary tumors in female rat offspring through transcriptome changes. Nutr. Res. 2019, 61, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Gautier, Y.; Luneau, I.; Coquery, N.; Meurice, P.; Malbert, C.H.; Guerin, S.; Kemp, B.; Bolhuis, J.E.; Clouard, C.; Le Huërou-Luron, I.; et al. Maternal Western diet during gestation and lactation modifies adult offspring’s cognitive and hedonic brain processes, behavior, and metabolism in Yucatan minipigs. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, fj201701541. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, I.U.; Luo, L.; Berton, T.R.; Schwartz, S.L.; Hursting, S.D.; Conti, C.J.; Fuchs-Young, R. Early Exposure to a High Fat/High Sugar Diet Increases the Mammary Stem Cell Compartment and Mammary Tumor Risk in Female Mice. Cancer Prev. Res. 2017, 10, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weldon, W.C.; Thulin, A.J.; MacDougald, O.A.; Johnston, L.J.; Miller, E.R.; Tucker, H.A. Effects of increased dietary energy and protein during late gestation on mammary development in gilts. J. Anim. Sci. 1991, 69, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Howard, K.A. Effect of Porcine Somatotropin, Soybean Oil and Energy Intake on Mammary Development in Gestating Gilts. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, October 1995. [Google Scholar]
- Singletary, K.W.; McNary, M.Q. Effect of moderate ethanol consumption on mammary gland structural development and DNA synthesis in the female rat. Alcohol 1992, 9, 95–101. [Google Scholar] [CrossRef]
- Harvie, M.; Howell, A. Energy restriction and the prevention of breast cancer. Proc. Nutr. Soc. 2012, 71, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogan, S.; Rogozina, O.P.; Lokshin, A.E.; Grande, J.P.; Cleary, M.P. Effects of chronic vs. intermittent calorie restriction on mammary tumor incidence and serum adiponectin and leptin levels in MMTV-TGF-α mice at different ages. Oncol. Lett. 2010, 1, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Rosso, P.; Keyou, G.; Bassi, J.A.; Slusser, W.M. Effect of malnutrition during pregnancy on the development of the mammary glands of rats. J. Nutr. 1981, 111, 1937–1941. [Google Scholar] [CrossRef] [PubMed]
- Ravi, R.T.; Ganguli, N.C. Effect of dietary protein restriction on chromatin proteins of rat mammary gland. J. Nutr. 1980, 110, 1144–1151. [Google Scholar] [CrossRef]
- Lyvers-Peffer, P.A.; Rozeboom, D. The effects of a growth-altering pre-pubertal feeding regimen on mammary development and parity-one lactation potential in swine. Livest. Prod. Sci. 2001, 70, 167–173. [Google Scholar] [CrossRef]
- Kfir, S.H.; Barash, I. Calorie restriction and rapamycin administration induce stem cell self-renewal and consequent development and production in the mammary gland. Exp. Cell Res. 2019, 382, 111477. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Cota, D.; Matter, E.K.; Woods, S.C.; Seeley, R.J. The role of hypothalamic mammalian target of rapamycin complex 1 signaling in diet-induced obesity. J. Neurosci. 2008, 28, 7202–7208. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.; Beltran, L.; Carbajal, S.; Strom, S.; Traag, J.; Hursting, S.D.; DiGiovanni, J. Dietary energy balance modulates signaling through the Akt/mammalian target of rapamycin pathways in multiple epithelial tissues. Cancer Prev. Res. 2008, 1, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Zhu, Z.; Thompson, H.J. Dietary energy restriction modulates the activity of AMP-activated protein kinase, Akt, and mammalian target of rapamycin in mammary carcinomas, mammary gland, and liver. Cancer Res. 2008, 68, 5492–5499. [Google Scholar] [CrossRef] [Green Version]
- Dogan, S.; Johannsen, A.C.; Grande, J.P.; Cleary, M.P. Effects of intermittent and chronic calorie restriction on mammalian target of rapamycin (mTOR) and IGF-I signaling pathways in mammary fat pad tissues and mammary tumors. Nutr. Cancer 2011, 63, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Knazek, R.A.; Liu, S.C.; Bodwin, J.S.; Vonderhaar, B.K. Requirement of essential fatty acids in the diet for development of the mouse mammary gland. J. Natl. Cancer Inst. 1980, 64, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.C.; Alves, B.R.; Prezotto, L.D.; Thorson, J.F.; Tedeschi, L.O.; Keisler, D.H.; Park, C.S.; Amstalden, M.; Williams, G.L. Use of a stair-step compensatory gain nutritional regimen to program the onset of puberty in beef heifers. J. Anim. Sci. 2014, 92, 2942–2949. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Baik, M.G.; Keller, W.L.; Slanger, W.D. Dietary energy restriction-mediated growth and mammary development in rats. J. Anim. Sci. 1994, 72, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Park, C.S.; Choi, Y.J.; Keller, W.L.; Harrold, R.L. Effects of compensatory growth on milk protein gene expression and mammary differentiation. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1988, 2, 2619–2624. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Moon, Y.S.; Keller, W.L.; Park, C.S. Compensatory nutrition-directed mammary cell proliferation and lactation in rats. Br. J. Nutr. 1998, 79, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Park, C.S. A Compensatory Nutrition Regimen during Gestation Stimulates Mammary Development and Lactation Potential in Rats. J. Nutr. 2004, 134, 756–761. [Google Scholar] [CrossRef] [Green Version]
- Grigor, M.R.; Gain, K.R. The effect of starvation and refeeding on lipogenic enzymes in mammary glands and livers of lactating rats. Biochem. J. 1983, 216, 515–518. [Google Scholar] [CrossRef]
- Farmer, C. Review: Mammary development in swine: Effects of hormonal status, nutrition and management. Can. J. Anim. Sci. 2013, 93, 1–7. [Google Scholar] [CrossRef]
- Farmer, C.; Palin, M.F.; Martel-Kennes, Y. Impact of diet deprivation and subsequent overallowance during gestation on mammary gland development and lactation performance. J. Anim. Sci. 2014, 92, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Mousa, A.; Naqash, A.; Lim, S. Macronutrient and Micronutrient Intake during Pregnancy: An Overview of Recent Evidence. Nutrients 2019, 11, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLennan, M.B.; Clarke, S.E.; Perez, K.; Wood, G.A.; Muller, W.J.; Kang, J.X.; Ma, D.W. Mammary tumor development is directly inhibited by lifelong n-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2013, 24, 388–395. [Google Scholar] [CrossRef]
- Hilakivi-Clarke, L.; Olivo, S.E.; Shajahan, A.; Khan, G.; Zhu, Y.; Zwart, A.; Cho, E.; Clarke, R. Mechanisms mediating the effects of prepubertal (n-3) polyunsaturated fatty acid diet on breast cancer risk in rats. J. Nutr. 2005, 135, 2946s–2952s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, B.M.; MacLennan, M.B.; Hillyer, L.M.; Ma, D.W. Lifelong exposure to n-3 PUFA affects pubertal mammary gland development. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Et Metab. 2014, 39, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delpal, S.; Pauloin, A.; Hue-Beauvais, C.; Berthelot, V.; Schmidely, P.; Ollivier-Bousquet, M. Effects of dietary fish oil and corn oil on rat mammary tissue. Cell Tissue Res. 2013, 351, 453–464. [Google Scholar] [CrossRef]
- Ollivier-Bousquet, M.; Guesnet, P.; Seddiki, T.; Durand, G. Deficiency of (n-6) but not (n-3) polyunsaturated fatty acids inhibits the secretagogue effect of prolactin in lactating rat mammary epithelial cells. J. Nutr. 1993, 123, 2090–2100. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Canela, M.; Martínez-González, M.A. Olive oil in the primary prevention of cardiovascular disease. Maturitas 2011, 68, 245–250. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, J.; Yuan, C.; Zhang, F.; Fu, Q.; Su, H.; Zhu, X.; Wang, L.; Gao, P.; Shu, G.; et al. Oleic acid stimulates HC11 mammary epithelial cells proliferation and mammary gland development in peripubertal mice through activation of CD36-Ca(2+) and PI3K/Akt signaling pathway. Oncotarget 2018, 9, 12982–12994. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, J.; Zhang, F.; Ai, W.; Zhu, X.; Shu, G.; Wang, L.; Gao, P.; Xi, Q.; Zhang, Y.; et al. Lauric Acid Stimulates Mammary Gland Development of Pubertal Mice through Activation of GPR84 and PI3K/Akt Signaling Pathway. J. Agric. Food Chem. 2017, 65, 95–103. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Q.; Li, F.; Yi, W.; Liu, F.; Wang, S.; Jiang, Q. Effects of Dietary Supplementation of Lauric Acid on Lactation Function, Mammary Gland Development, and Serum Lipid Metabolites in Lactating Mice. Anim. Open Access J. Mdpi. 2020, 10, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, C.J.; Rodriguez-Gonzalez, G.L.; Torres, N.; Hernandez-Pando, R.; Ramirez, V.; Rodriguez-Cruz, M.; Nathanielsz, P.W.; Zambrano, E. Protein restriction in the rat negatively impacts long-chain polyunsaturated fatty acid composition and mammary gland development at the end of gestation. Arch. Med Res. 2013, 44, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Bautista, C.J.; Bautista, R.J.; Montaño, S.; Reyes-Castro, L.A.; Rodriguez-Peña, O.N.; Ibáñez, C.A.; Nathanielsz, P.W.; Zambrano, E. Effects of maternal protein restriction during pregnancy and lactation on milk composition and offspring development. Br. J. Nutr. 2019, 122, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Pau, M.Y.; Milner, J.A. Arginine deficiency during gestation and lactation in the rat. J. Nutr. 1981, 111, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Eason, R.R.; Velarde, M.C.; Chatman, L., Jr.; Till, S.R.; Geng, Y.; Ferguson, M.; Badger, T.M.; Simmen, R.C. Dietary exposure to whey proteins alters rat mammary gland proliferation, apoptosis, and gene expression during postnatal development. J. Nutr. 2004, 134, 3370–3377. [Google Scholar] [CrossRef] [PubMed]
- Franke, T.F.; Hornik, C.P.; Segev, L.; Shostak, G.A.; Sugimoto, C. PI3K/Akt and apoptosis: Size matters. Oncogene 2003, 22, 8983–8998. [Google Scholar] [CrossRef] [Green Version]
- Kusina, J.; Pettigrew, J.E.; Sower, A.F.; Hathaway, M.R.; White, M.E.; Crooker, B.A. Effect of protein intake during gestation on mammary development of primiparous sows. J. Anim. Sci. 1999, 77, 925–930. [Google Scholar] [CrossRef]
- Che, L.; Xu, M.; Gao, K.; Wang, L.; Yang, X.; Wen, X.; Xiao, H.; Jiang, Z. Effects of dietary valine supplementation during late gestation on the reproductive performance and mammary gland development of gilts. J. Anim. Sci. Biotechnol. 2020, 11, 15. [Google Scholar] [CrossRef]
- Krogh, U.; Oksbjerg, N.; Storm, A.C.; Feyera, T.; Theil, P.K. Mammary nutrient uptake in multiparous sows fed supplementary arginine during gestation and lactation. J. Anim. Sci. 2017, 95, 2517–2532. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Hurley, W.L.; Wu, G.; Ji, F. Ideal amino acid balance for sows during gestation and lactation. J. Anim. Sci. 2009, 87, E123–E132. [Google Scholar] [CrossRef] [Green Version]
- Holanda, D.M.; Marcolla, C.S.; Guimarães, S.E.F.; Neves, M.M.; Hausman, G.J.; Duarte, M.S.; Abreu, M.L.T.; Saraiva, A. Dietary L-arginine supplementation increased mammary gland vascularity of lactating sows. Animal 2019, 13, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Adewole, D.; Liu, S.; Xi, P.; Yang, C.; Yin, Y. Tryptophan Supplementation Increases Reproduction Performance, Milk Yield, and Milk Composition in Lactating Sows and Growth Performance of Their Piglets. J. Agric. Food Chem. 2019, 67, 5096–5104. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; He, L.; Hou, Y.; Chen, J.; Duan, Y.; Deng, D.; Wu, G.; Yin, Y.; Yao, K. α-ketoglutarate enhances milk protein synthesis by porcine mammary epithelial cells. Amino. Acids 2016, 48, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Adebowale, T.O.; Tian, J.; Yin, Y.; Yao, K. Effects of maternal α-ketoglutarate supplementation during lactation on the performance of lactating sows and suckling piglets. Arch Anim. Nutr. 2019, 73, 457–471. [Google Scholar] [CrossRef]
- Koski, K.G.; Hill, F.W.; Lönnerdal, B. Altered lactational performance in rats fed low carbohydrate diets and its effect on growth of neonatal rat pups. J. Nutr. 1990, 120, 1028–1036. [Google Scholar] [CrossRef]
- Koski, K.G.; Lanoue, L.; Young, S.N. Restriction of maternal dietary carbohydrate decreases fetal brain indoles and glycogen in rats. J. Nutr. 1993, 123, 42–51. [Google Scholar] [CrossRef]
- McDaniel, S.M.; O’Neill, C.; Metz, R.P.; Tarbutton, E.; Stacewicz-Sapuntzakis, M.; Heimendinger, J.; Wolfe, P.; Thompson, H.; Schedin, P. Whole-food sources of vitamin A more effectively inhibit female rat sexual maturation, mammary gland development, and mammary carcinogenesis than retinyl palmitate. J. Nutr. 2007, 137, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Cabezuelo, M.T.; Zaragozá, R.; Barber, T.; Viña, J.R. Role of Vitamin A in Mammary Gland Development and Lactation. Nutrients 2019, 12, 80. [Google Scholar] [CrossRef] [Green Version]
- Spear, A.T.; Sherman, A.R. Iron deficiency alters DMBA-induced tumor burden and natural killer cell cytotoxicity in rats. J. Nutr. 1992, 122, 46–55. [Google Scholar] [CrossRef]
- Grill, C.J.; Cohick, W.S.; Sherman, A.R. Postpubertal development of the rat mammary gland is preserved during iron deficiency. J. Nutr. 2001, 131, 1444–1448. [Google Scholar] [CrossRef]
- Bostanci, Z.; Mack, R.P., Jr.; Lee, S.; Soybel, D.I.; Kelleher, S.L. Paradoxical zinc toxicity and oxidative stress in the mammary gland during marginal dietary zinc deficiency. Reprod. Toxicol. 2015, 54, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Dempsey, C.; McCormick, N.H.; Croxford, T.P.; Seo, Y.A.; Grider, A.; Kelleher, S.L. Marginal maternal zinc deficiency in lactating mice reduces secretory capacity and alters milk composition. J. Nutr. 2012, 142, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Murrill, W.B.; Brown, N.M.; Zhang, J.X.; Manzolillo, P.A.; Barnes, S.; Lamartiniere, C.A. Prepubertal genistein exposure suppresses mammary cancer and enhances gland differentiation in rats. Carcinogenesis 1996, 17, 1451–1457. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.A., Jr.; Clark, S.G.; Walters, E.M.; Wheeler, M.B.; Hurley, W.L. Estrogenic effects of genistein on reproductive tissues of ovariectomized gilts. J. Anim. Sci. 2006, 84, 834–842. [Google Scholar] [CrossRef]
- Tou, J.C.; Thompson, L.U. Exposure to flaxseed or its lignan component during different developmental stages influences rat mammary gland structures. Carcinogenesis 1999, 20, 1831–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, W.A.; Coward, L.; Wang, J.; Lamartiniere, C.A. Dietary genistein: Perinatal mammary cancer prevention, bioavailability and toxicity testing in the rat. Carcinogenesis 1998, 19, 2151–2158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, W.E.; Jiang, F.O.; Thompson, L.U. Exposure to flaxseed or purified lignan during lactation influences rat mammary gland structures. Nutr Cancer 2000, 37, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Farmer, C.; Palin, M.F.; Gilani, G.S.; Weiler, H.; Vignola, M.; Choudhary, R.K.; Capuco, A.V. Dietary genistein stimulates mammary hyperplasia in gilts. Animal 2010, 4, 454–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, C.; Petit, H.V.; Weiler, H.; Capuco, A.V. Effects of dietary supplementation with flax during prepuberty on fatty acid profile, mammogenesis, and bone resorption in gilts. J. Anim. Sci. 2007, 85, 1675–1686. [Google Scholar] [CrossRef]
- Wink, M.; Roberts, M.F. Compartmentation of Alkaloid Synthesis, Transport, and Storage; Roberts, M.F., Wink, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar] [CrossRef]
- Kopinski, J.S.; Blaney, B.J.; Downing, J.A.; McVeigh, J.F.; Murray, S.A. Feeding sorghum ergot (Claviceps africana) to sows before farrowing inhibits milk production. Aust. Vet. J. 2007, 85, 169–176. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev. 2004, 44, 467–492. [Google Scholar] [CrossRef] [PubMed]
- Coulon, J.B.; Dupont, D.; Pochet, S.; Pradel, P.; Duployer, H. Effect of genetic potential and level of feeding on milk protein composition. J. Dairy Res. 2001, 68, 569–577. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Tech. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahnadi, C.E.; Beswick, N.; Delbecchi, L.; Kennelly, J.J.; Lacasse, P. Addition of fish oil to diets for dairy cows. II. Effects on milk fat and gene expression of mammary lipogenic enzymes. J. Dairy Res. 2002, 69, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, A.; Bernard, L.; Meynadier, A.; Malpuech-Brugere, C. Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: A review. Biochimie 2017, 141, 107–120. [Google Scholar] [CrossRef]
- Angulo, J.; Mahecha, L.; Nuernberg, K.; Nuernberg, G.; Dannenberger, D.; Olivera, M.; Boutinaud, M.; Leroux, C.; Albrecht, E.; Bernard, L. Effects of polyunsaturated fatty acids from plant oils and algae on milk fat yield and composition are associated with mammary lipogenic and SREBF1 gene expression. Animal 2012, 6, 1961–1972. [Google Scholar] [CrossRef] [Green Version]
- Piperova, L.S.; Teter, B.B.; Bruckental, I.; Sampugna, J.; Mills, S.E.; Yurawecz, M.P.; Fritsche, J.; Ku, K.; Erdman, R.A. Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat-depressing diet. J. Nutr. 2000, 130, 2568–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvatine, K.J.; Bauman, D.E. SREBP1 and thyroid hormone responsive spot 14 (S14) are involved in the regulation of bovine mammary lipid synthesis during diet-induced milk fat depression and treatment with CLA. J. Nutr. 2006, 136, 2468–2474. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.G.; Matitashvili, E.A.; Bauman, D.E. Diet-induced milk fat depression in dairy cows results in increased trans-10, cis-12 CLA in milk fat and coordinate suppression of mRNA abundance for mammary enzymes involved in milk fat synthesis. J. Nutr. 2003, 133, 3098–3102. [Google Scholar] [CrossRef] [Green Version]
- Castro-Carrera, T.; Frutos, P.; Leroux, C.; Chilliard, Y.; Hervas, G.; Belenguer, A.; Bernard, L.; Toral, P.G. Dietary sunflower oil modulates milk fatty acid composition without major changes in adipose and mammary tissue fatty acid profile or related gene mRNA abundance in sheep. Animal 2015, 9, 582–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toral, P.G.; Hervas, G.; Carreno, D.; Belenguer, A.; Frutos, P. Comparison of milk fatty acid responses during fish oil- and trans-10 cis-12 18:2-induced milk fat depression in dairy ewes. Anim. Feed Sci. Tech. 2015, 210, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Toral, P.G.; Hervas, G.; Belenguer, A.; Carreno, D.; Frutos, P. mRNA abundance of genes involved in mammary lipogenesis during fish oil- or trans-10,cis-12 CLA-induced milk fat depression in dairy ewes. J. Dairy Sci. 2017, 100, 3182–3192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frutos, P.; Toral, P.G.; Hervas, G. Individual variation of the extent of milk fat depression in dairy ewes fed fish oil: Milk fatty acid profile and mRNA abundance of candidate genes involved in mammary lipogenesis. J. Dairy Sci. 2017, 100, 9611–9622. [Google Scholar] [CrossRef] [Green Version]
- Shingfield, K.J.; Bernard, L.; Leroux, C.; Chilliard, Y. Role of trans fatty acids in the nutritional regulation of mammary lipogenesis in ruminants. Animal 2010, 4, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Toral, P.G.; Bernard, L.; Delavaud, C.; Gruffat, D.; Leroux, C.; Chilliard, Y. Effects of fish oil and additional starch on tissue fatty acid profile and lipogenic gene mRNA abundance in lactating goats fed a diet containing sunflower-seed oil. Animal 2013, 7, 948–956. [Google Scholar] [CrossRef]
- Fougere, H.; Bernard, L. Effect of diets supplemented with starch and corn oil, marine algae, or hydrogenated palm oil on mammary lipogenic gene expression in cows and goats: A comparative study. J. Dairy Sci. 2019, 102, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Zhang, T.; Li, C.; Wang, J.; Huang, J.; Li, Z. trans-10,cis-12-Conjugated Linoleic Acid Affects Expression of Lipogenic Genes in Mammary Glands of Lactating Dairy Goats. J. Agric. Food Chem. 2017, 65, 9460–9467. [Google Scholar] [CrossRef]
- Bernard, L.; Rouel, J.; Leroux, C.; Ferlay, A.; Faulconnier, Y.; Legrand, P.; Chilliard, Y. Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable lipids. J. Dairy Sci. 2005, 88, 1478–1489. [Google Scholar] [CrossRef] [Green Version]
- Bernard, L.; Bonnet, M.; Leroux, C.; Shingfield, K.J.; Chilliard, Y. Effect of sunflower-seed oil and linseed oil on tissue lipid metabolism, gene expression, and milk fatty acid secretion in Alpine goats fed maize silage-based diets. J. Dairy Sci. 2009, 92, 6083–6094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, A.A.; Dijkstra, J.; Liesman, J.S.; Vandehaar, M.J.; Lock, A.L.; van Vuuren, A.M.; Hendriks, W.H.; van Baal, J. Effects of short- and long-chain fatty acids on the expression of stearoyl-CoA desaturase and other lipogenic genes in bovine mammary epithelial cells. Animal 2013, 7, 1508–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Invernizzi, G.; Thering, B.J.; McGuire, M.A.; Savoini, G.; Loor, J.J. Sustained upregulation of stearoyl-CoA desaturase in bovine mammary tissue with contrasting changes in milk fat synthesis and lipogenic gene networks caused by lipid supplements. Funct. Integr. Genom. 2010, 10, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Mouriot, J.; Rouel, J.; Glasser, F.; Capitan, P.; Pujos-Guillot, E.; Chardigny, J.M.; Chilliard, Y. Effects of fish oil and starch added to a diet containing sunflower-seed oil on dairy goat performance, milk fatty acid composition and in vivo delta9-desaturation of [13C]vaccenic acid. Br. J. Nutr. 2010, 104, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Vahmani, P.; Glover, K.E.; Fredeen, A.H. Effects of pasture versus confinement and marine oil supplementation on the expression of genes involved in lipid metabolism in mammary, liver, and adipose tissues of lactating dairy cows. J. Dairy Sci. 2014, 97, 4174–4183. [Google Scholar] [CrossRef]
- Bichi, E.; Frutos, P.; Toral, P.G.; Keisler, D.; Hervas, G.; Loor, J.J. Dietary marine algae and its influence on tissue gene network expression during milk fat depression in dairy ewes. Anim Feed Sci. Tech. 2013, 186, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Mach, N.; Van Baal, J.; Kruijt, L.; Jacobs, A.; Smits, M. Dietary unsaturated fatty acids affect the mammary gland integrity and health in lactating dairy cows. BMC Proc. 2011, 3, 1753–6561. [Google Scholar] [CrossRef] [PubMed]
- Ibeagha-Awemu, E.M.; Li, R.; Ammah, A.A.; Dudemaine, P.L.; Bissonnette, N.; Benchaar, C.; Zhao, X. Transcriptome adaptation of the bovine mammary gland to diets rich in unsaturated fatty acids shows greater impact of linseed oil over safflower oil on gene expression and metabolic pathways. BMC Genom. 2016, 17, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroux, C.; Bernard, L.; Faulconnier, Y.; Rouel, J.; De la Foye, A.; Domagalski, J.; Chilliard, Y. Bovine Mammary Nutrigenomics and Changes in the Milk Composition due to Rapeseed or Sunflower Oil Supplementation of High-Forage or High-Concentrate Diets. J. Nutr. Nutr. 2016, 9, 65–82. [Google Scholar] [CrossRef]
- Ollier, S.; Leroux, C.; De la Foye, A.; Bernard, L.; Rouel, J.; Chilliard, Y. Whole intact rapeseeds or sunflower oil in high-forage or high-concentrate diets affects milk yield, milk composition, and mammary gene expression profile in goats. J. Dairy Sci. 2009, 92, 5544–5560. [Google Scholar] [CrossRef]
- Faulconnier, Y.; Bernard, L.; Boby, C.; Domagalski, J.; Chilliard, Y.; Leroux, C. Extruded linseed alone or in combination with fish oil modifies mammary gene expression profiles in lactating goats. Animal 2018, 12, 1564–1575. [Google Scholar] [CrossRef] [PubMed]
- Canovas, A.; Rincon, G.; Bevilacqua, C.; Islas-Trejo, A.; Brenaut, P.; Hovey, R.C.; Boutinaud, M.; Morgenthaler, C.; VanKlompenberg, M.K.; Martin, P.; et al. Comparison of five different RNA sources to examine the lactating bovine mammary gland transcriptome using RNA-Sequencing. Sci. Rep. 2014, 4, 5297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Vega, A.; Gutierrez-Gil, B.; Toral, P.G.; Hervas, G.; Arranz, J.J.; Frutos, P. Conjugated linoleic acid (CLA)-induced milk fat depression: Application of RNA-Seq technology to elucidate mammary gene regulation in dairy ewes. Sci. Rep. 2019, 9, 4473. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eleswarapu, S.; Jiang, H. Identification and characterization of microRNAs from the bovine adipose tissue and mammary gland. Febs. Lett. 2007, 581, 981–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guillou, S.; Marthey, S.; Laloe, D.; Laubier, J.; Mobuchon, L.; Leroux, C.; Le Provost, F. Characterisation and comparison of lactating mouse and bovine mammary gland miRNomes. PloS ONE 2014, 9, e91938. [Google Scholar] [CrossRef] [Green Version]
- Mobuchon, L.; Marthey, S.; Le Guillou, S.; Laloe, D.; Le Provost, F.; Leroux, C. Food Deprivation Affects the miRNome in the Lactating Goat Mammary Gland. PloS ONE 2015, 10, e0140111. [Google Scholar] [CrossRef]
- Li, R.; Dudemaine, P.L.; Zhao, X.; Lei, C.; Ibeagha-Awemu, E.M. Comparative Analysis of the miRNome of Bovine Milk Fat, Whey and Cells. PloS ONE 2016, 11, e0154129. [Google Scholar] [CrossRef] [Green Version]
- Mobuchon, L.; Le Guillou, S.; Marthey, S.; Laubier, J.; Laloe, D.; Bes, S.; Le Provost, F.; Leroux, C. Sunflower oil supplementation affects the expression of miR-20a-5p and miR-142-5p in the lactating bovine mammary gland. PloS ONE 2017, 12, e0185511. [Google Scholar] [CrossRef]
- Li, R.; Beaudoin, F.; Ammah, A.A.; Bissonnette, N.; Benchaar, C.; Zhao, X.; Lei, C.; Ibeagha-Awemu, E.M. Deep sequencing shows microRNA involvement in bovine mammary gland adaptation to diets supplemented with linseed oil or safflower oil. BMC Genom. 2015, 16, 884. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; Bauman, D.E. Adaptations of glucose metabolism during pregnancy and lactation. J. Mammary Gland Biol. Neoplasia 1997, 2, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Lérias, J.R.; Hernández-Castellano, L.E.; Morales-Delanuez, A.; Araújo, S.S.; Castro, N.; Argüello, A.; Capote, J.; Almeida, A.M. Body live weight and milk production parameters in the Majorera and Palmera goat breeds from the Canary Islands: Influence of weight loss. Trop. Anim. Health Prod. 2013, 45, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Cugno, G.; Parreira, J.R.; Ferlizza, E.; Hernandez-Castellano, L.E.; Carneiro, M.; Renaut, J.; Castro, N.; Arguello, A.; Capote, J.; Campos, A.M.; et al. The Goat (Capra hircus) Mammary Gland Mitochondrial Proteome: A Study on the Effect of Weight Loss Using Blue-Native PAGE and Two-Dimensional Gel Electrophoresis. PloS ONE 2016, 11, e0151599. [Google Scholar] [CrossRef] [Green Version]
- Parreira, J.R.; Hernandez-Castellano, L.E.; Arguello, A.; Capote, J.; Castro, N.; de Sousa Araujo, S.; de Almeida, A.M. Understanding seasonal weight loss tolerance in dairy goats: A transcriptomics approach. Bmc Genom. 2020, 21, 629. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.; Van Dorland, H.A.; Bruckmaier, R.M.; Schwarz, F.J. Performance and metabolic profile of dairy cows during a lactational and deliberately induced negative energy balance with subsequent realimentation. J. Dairy Sci. 2011, 94, 1820–1830. [Google Scholar] [CrossRef]
- Bjerre-Harpoth, V.; Friggens, N.C.; Thorup, V.M.; Larsen, T.; Damgaard, B.M.; Ingvartsen, K.L.; Moyes, K.M. Metabolic and production profiles of dairy cows in response to decreased nutrient density to increase physiological imbalance at different stages of lactation. J. Dairy Sci. 2012, 95, 2362–2380. [Google Scholar] [CrossRef] [Green Version]
- Friggens, N.C.; Duvaux-Ponter, C.; Etienne, M.P.; Mary-Huard, T.; Schmidely, P. Characterizing individual differences in animal responses to a nutritional challenge: Toward improved robustness measures. J. Dairy Sci. 2016, 99, 2704–2718. [Google Scholar] [CrossRef]
- Abdelatty, A.M.; Iwaniuk, M.E.; Garcia, M.; Moyes, K.M.; Teter, B.B.; Delmonte, P.; Kadegowda, A.K.G.; Tony, M.A.; Mohamad, F.F.; Erdman, R.A. Effect of short-term feed restriction on temporal changes in milk components and mammary lipogenic gene expression in mid-lactation Holstein dairy cows. J. Dairy Sci. 2017, 100, 4000–4013. [Google Scholar] [CrossRef] [Green Version]
- Pires, J.A.A.; Pawlowski, K.; Rouel, J.; Delavaud, C.; Foucras, G.; Germon, P.; Leroux, C. Undernutrition modified metabolic responses to intramammary lipopolysaccharide but had limited effects on selected inflammation indicators in early-lactation cows. J. Dairy Sci. 2019, 102, 5347–5360. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Loor, J.J. ACSL1, AGPAT6, FABP3, LPIN1, and SLC27A6 are the most abundant isoforms in bovine mammary tissue and their expression is affected by stage of lactation. J. Nutr. 2008, 138, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Delbecchi, L.; Petitclerc, D.; Wagner, G.F.; Talbot, B.G.; Lacasse, P. Effect of stage of lactation and parity on mammary gland cell renewal. J. Dairy Sci. 2006, 89, 4669–4677. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Flemetakis, E.; Kouri, E.D.; Sotirakoglou, K.; Zervas, G. The effect of long term under- and over-feeding on the expression of genes related to lipid metabolism in mammary tissue of sheep. J. Dairy Res. 2015, 82, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Tsiplakou, E.; Chadio, S.; Zervas, G. The effect of long term under- and over-feeding of sheep on milk and plasma fatty acid profiles and on insulin and leptin concentrations. J. Dairy Res. 2012, 79, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Massart-Leen, A.M.; Peeters, G. Changes in the fatty acid composition of goat milk fat after a 48-hour fast. Reprod. Nutr. Dev. 1985, 25, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Ollier, S.; Robert-Granié, C.; Bernard, L.; Chilliard, Y.; Leroux, C. Mammary transcriptome analysis of food-deprived lactating goats highlights genes involved in milk secretion and programmed cell death. J. Nutr. 2007, 137, 560–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiplakou, E.; Flemetakis, E.; Kouri, E.D.; Zervas, G. The effect of long term under- and over-feeding on the expression of genes related to lipid metabolism in the mammary tissue of goats. Livest. Sci. 2015, 173, 32–37. [Google Scholar] [CrossRef]
- Moyes, K.M.; Drackley, J.K.; Morin, D.E.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Loor, J.J. Predisposition of cows to mastitis in non-infected mammary glands: Effects of dietary-induced negative energy balance during mid-lactation on immune-related genes. Funct. Integr. Genom. 2011, 11, 151–156. [Google Scholar] [CrossRef]
- Pawlowski, K.; Pires, J.A.A.; Faulconnier, Y.; Chambon, C.; Germon, P.; Boby, C.; Leroux, C. Mammary Gland Transcriptome and Proteome Modifications by Nutrient Restriction in Early Lactation Holstein Cows Challenged with Intra-Mammary Lipopolysaccharide. Int. J. Mol. Sci. 2019, 20, 1156. [Google Scholar] [CrossRef] [Green Version]
- Billa, P.A.; Faulconnier, Y.; Ye, T.; Bourdon, C.; Pires, J.A.A.; Leroux, C. Nutrigenomic analyses reveal miRNAs and mRNAs affected by feed restriction in the mammary gland of midlactation dairy cows. PLoS ONE 2021, in press. [Google Scholar]
- Martin, P.; Szymanowska, M.; Zwierzchowski, L.; Leroux, C. The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev. 2002, 42, 433–459. [Google Scholar] [CrossRef]
- Shah, M.S.; Davidson, L.A.; Chapkin, R.S. Mechanistic insights into the role of microRNAs in cancer: Influence of nutrient crosstalk. Front. Genet. 2012, 3, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Segura, L.; Perez-Andrade, M.; Miranda-Rios, J. The emerging role of MicroRNAs in the regulation of gene expression by nutrients. J. Nutr. Nutr. 2013, 6, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Moisa, S.; Khan, M.J.; Wang, J.; Bu, D.; Loor, J.J. MicroRNA expression patterns in the bovine mammary gland are affected by stage of lactation. J. Dairy Sci. 2012, 95, 6529–6535. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Liu, Z.; Chao, T.; Hou, L.; Fan, R.; He, R.; Wang, G.; Wang, J. Screening of miRNA profiles and construction of regulation networks in early and late lactation of dairy goat mammary glands. Sci. Rep. 2017, 7, 11933. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Ferreira, A.M.; Nanni, P.; Grossmann, J.; Arguello, A.; Capote, J.; Cai, G.; Lippolis, J.; Castro, N.; De Almeida, A.M. The goat (Capra hircus) mammary gland secretory tissue proteome as influenced by weight loss: A study using label free proteomics. J. Proteom. 2016, 145, 60–69. [Google Scholar] [CrossRef]
- Pawlowski, K.; Lago-Novais, D.; Bevilacqua, C.; Mobuchon, L.; Crapart, N.; Faulconnier, Y.; Boby, C.; Carvalho, G.; Martin, P.; Leroux, C. Different miRNA contents between mammary epithelial cells and milk fat globules: A random or a targeted process? Mol. Biol. Rep. 2020, 47, 8259–8264. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.L.; Smalley, M.J. Integrating single-cell RNA-sequencing and functional assays to decipher mammary cell states and lineage hierarchies. NPJ Breast Cancer 2020, 6, 32. [Google Scholar] [CrossRef]
- Kanaya, N.; Chang, G.; Wu, X.; Saeki, K.; Bernal, L.; Shim, H.-J.; Wang, J.; Warden, C.; Yamamoto, T.; Li, J.; et al. Single-cell RNA-sequencing analysis of estrogen- and endocrine-disrupting chemical-induced reorganization of mouse mammary gland. Commun. Biol. 2019, 2, 406. [Google Scholar] [CrossRef] [Green Version]
- Bartol, F.F.; Bagnell, C.A. Lactocrine programming of female reproductive tract development: Environmental connections to the reproductive continuum. Mol. Cell. Endocrinol. 2013, 354, 16–21. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hue-Beauvais, C.; Faulconnier, Y.; Charlier, M.; Leroux, C. Nutritional Regulation of Mammary Gland Development and Milk Synthesis in Animal Models and Dairy Species. Genes 2021, 12, 523. https://doi.org/10.3390/genes12040523
Hue-Beauvais C, Faulconnier Y, Charlier M, Leroux C. Nutritional Regulation of Mammary Gland Development and Milk Synthesis in Animal Models and Dairy Species. Genes. 2021; 12(4):523. https://doi.org/10.3390/genes12040523
Chicago/Turabian StyleHue-Beauvais, Cathy, Yannick Faulconnier, Madia Charlier, and Christine Leroux. 2021. "Nutritional Regulation of Mammary Gland Development and Milk Synthesis in Animal Models and Dairy Species" Genes 12, no. 4: 523. https://doi.org/10.3390/genes12040523
APA StyleHue-Beauvais, C., Faulconnier, Y., Charlier, M., & Leroux, C. (2021). Nutritional Regulation of Mammary Gland Development and Milk Synthesis in Animal Models and Dairy Species. Genes, 12(4), 523. https://doi.org/10.3390/genes12040523