
LncMyoD Promotes Skeletal Myogenesis and Regulates Skeletal Muscle Fiber-Type Composition by Sponging miR-370-3p

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Transfection
2.3. Animals and Collection of Muscles
2.4. Cell Proliferation Assay
2.5. Immunofluorescence Analysis
2.6. Isolation of RNA and RT-PCR
2.7. Luciferase Reporter Assay
2.8. RIP Assay
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
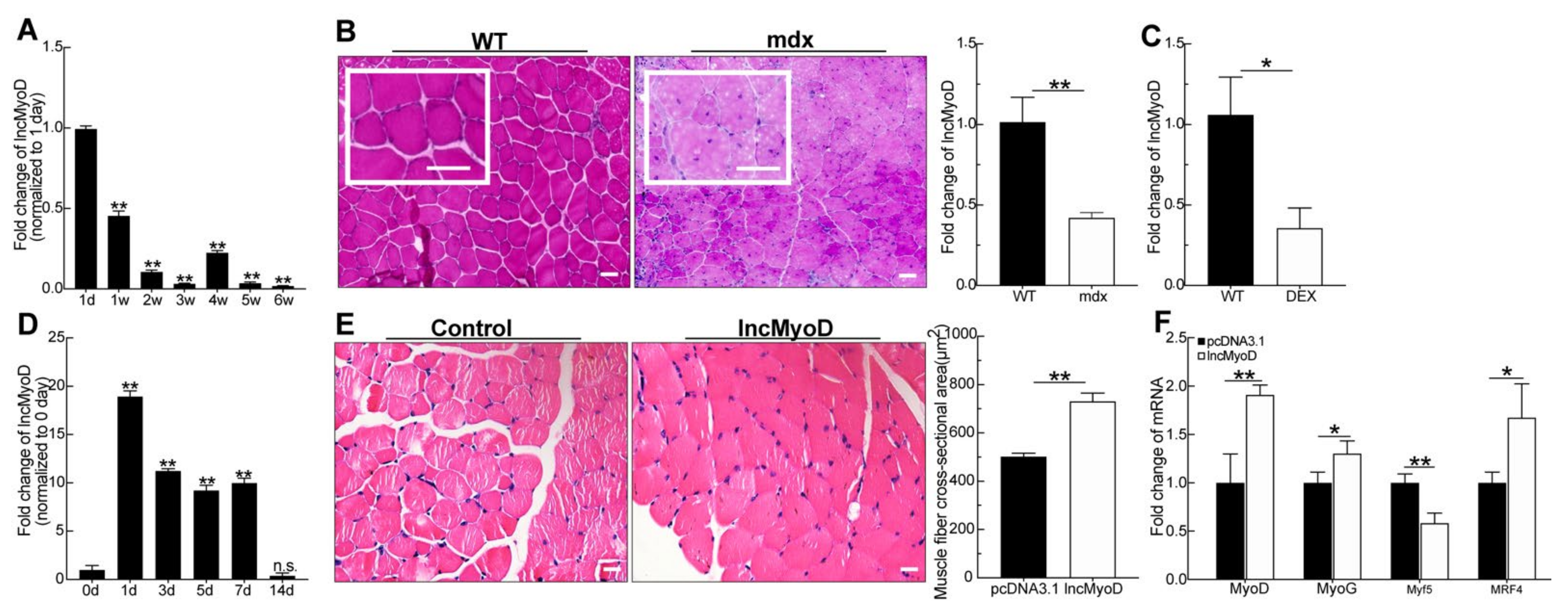
3.1. LncMyoD Is Associated with Skeletal Myogenesis
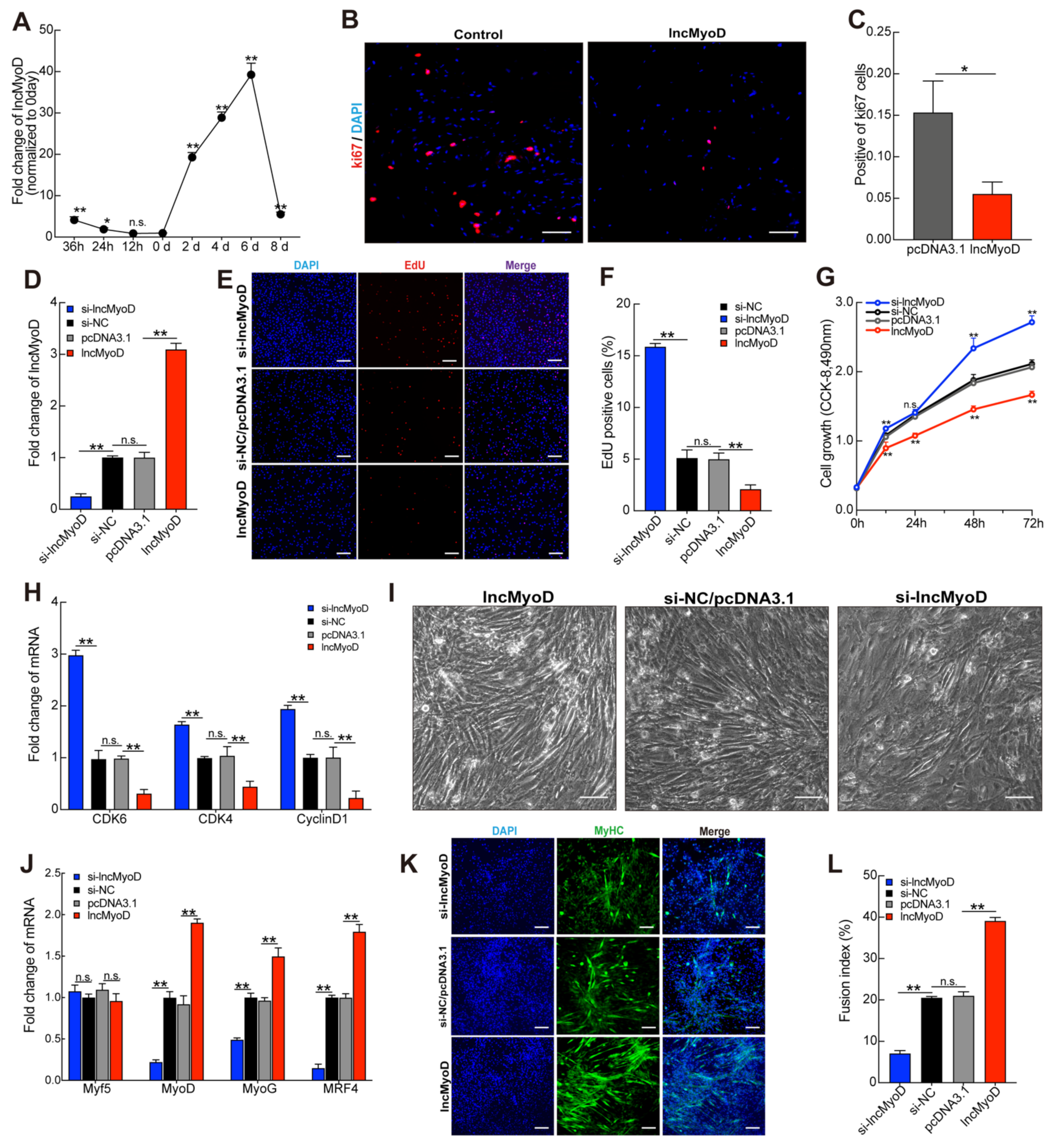
3.2. LncMyoD Regulates Myoblast Proliferation and Differentiation
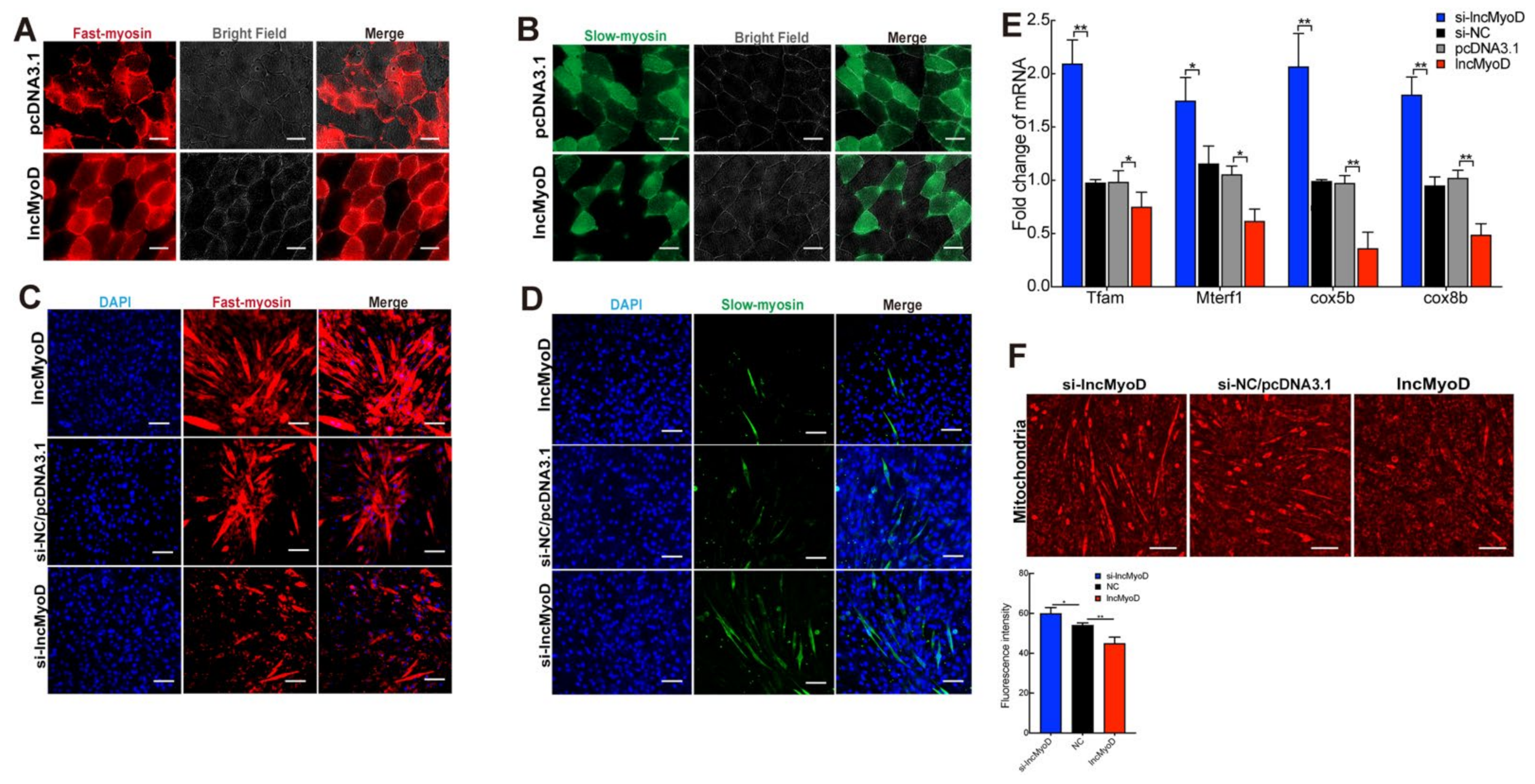
3.3. LncMyoD Regulates the Composition of Myofiber Type
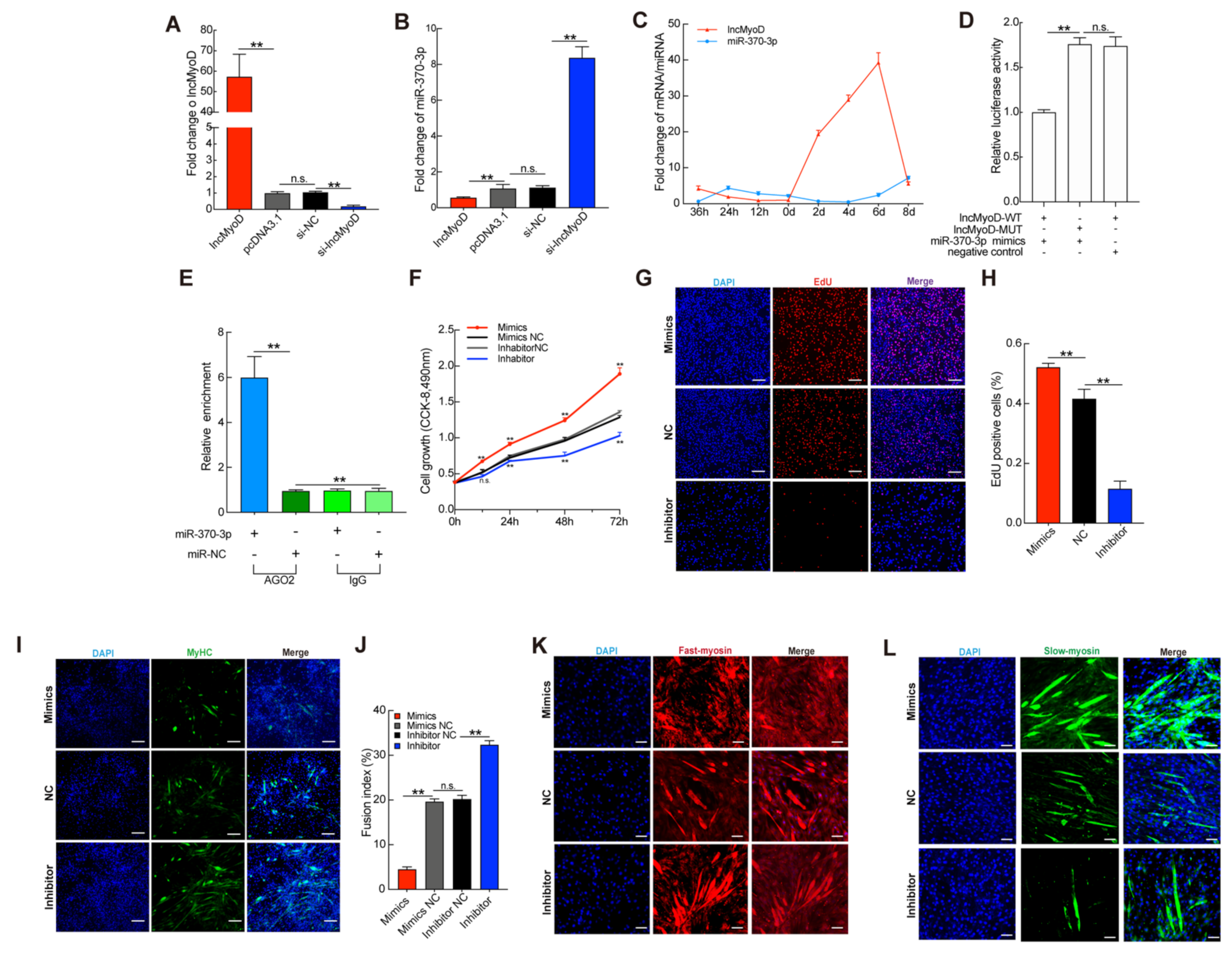
3.4. miR-370-3p Is Involved in Myogenesis and Regulates the Composition of Myofibers
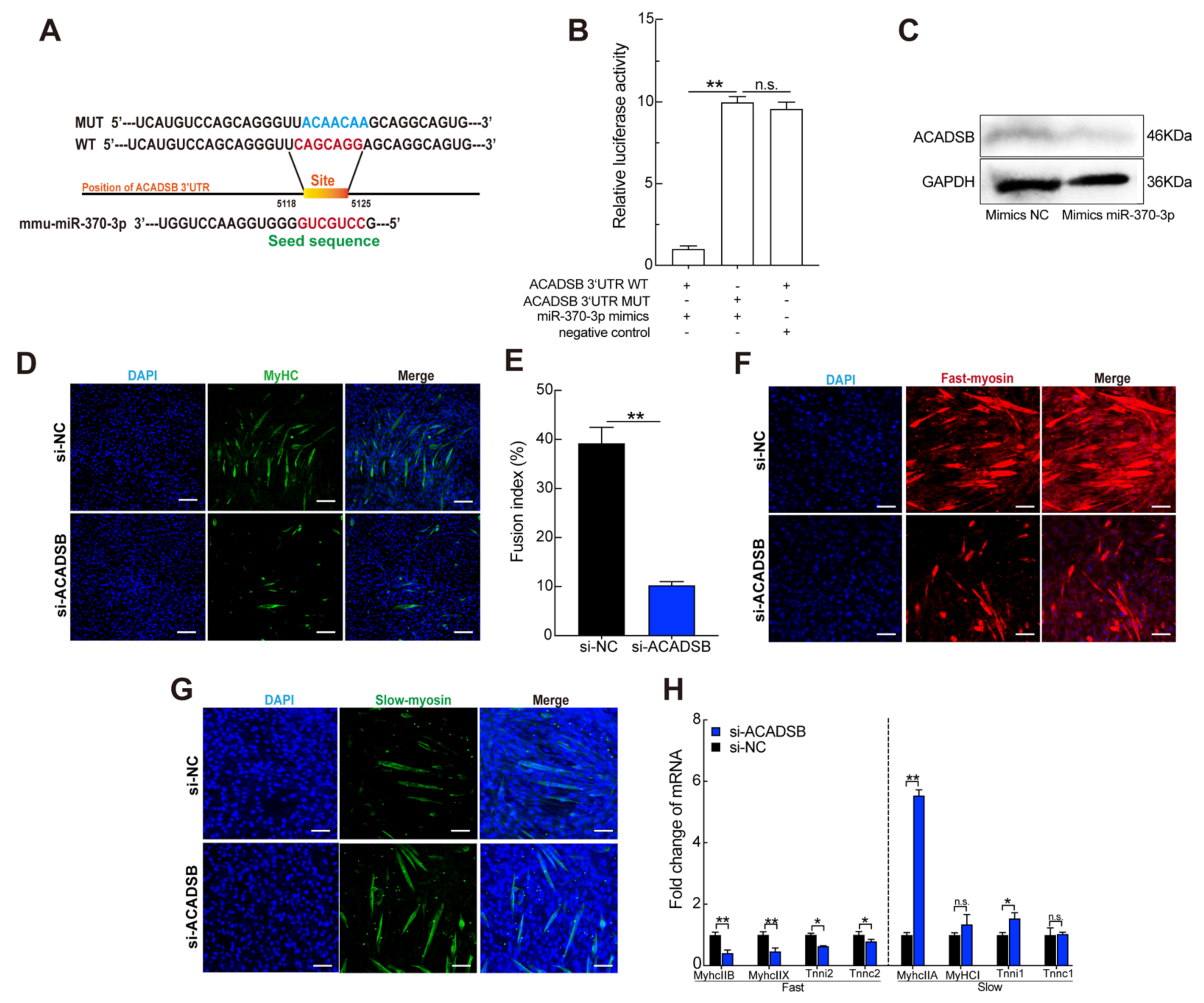
3.5. ACADSB Is a Direct Target of miR-370-3p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verche, V.L.; Sunshine, S.S.; Hammers, D.; Sweeney, H.L.; Paushkin, S.J. Chapter 21–Skeletal Muscle in Spinal Muscular Atrophy As an Opportunity for Therapeutic Intervention. Spinal Muscular Atrophy 2017, 341–356. [Google Scholar] [CrossRef]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.-Y.; Choi, I. Multifaceted Interweaving Between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K. Muscle as a secretory organ. Compr. Physiol. 2013, 3, 1337–1362. [Google Scholar]
- Buckingham, M.; Bajard, L.; Chang, T.; Daubas, P.; Hadchouel, J.; Meilhac, S.; Montarras, D.; Rocancourt, D.; Relaix, F. The formation of skeletal muscle: From somite to limb. J. Anat. 2003, 202, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Weigert, C. Skeletal Muscle as an Endocrine Organ: The Role of Myokines in Exercise Adaptations. Cold Spring Harb. Perspect. Med. 2017, 7, a029793. [Google Scholar] [CrossRef]
- Church, J.C.T.; Noronha, R.F.X.; Allbrook, D.B. Satellite cells and skeletal muscle regeneration. BJS 2005, 53, 638–642. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber Types in Mammalian Skeletal Muscles. Hysiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.A.; Suryawan, A.; Orellana, R.A.; Nguyen, H.V.; Fiorotto, M.L. Postnatal ontogeny of skeletal muscle protein synthesis in pigs. J. Anim. Sci. 2008, 86, E13–E18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeldt, C.; Henning, M.; Science, I.F.J.L. Consequences of pig domestication for skeletal muscle growth and cellularity. Livest. Sci. 2008, 116, 30–41. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome Regulation by Long Noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.J.; Chang, H.Y.J.C. Long Noncoding RNAs: Cellular Address Codes in Development and Disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, Y.; Bao, X.; Zhu, X.; Kwok, Y.K.-Y.; Sun, K.; Chen, X.; Huang, Y.; Jauch, R.; Esteban, M.; et al. LncRNA Dum interacts with Dnmts to regulate Dppa2 expression during myogenic differentiation and muscle regeneration. Cell Res. 2015, 25, 335–350. [Google Scholar] [CrossRef] [Green Version]
- Blin-Wakkach, C.; Lezot, F.; Ghoul-Mazgar, S.; Hotton, D.; Monteiro, S.; Teillaud, C.; Pibouin, L.; Orestes-Cardoso, S.; Papagerakis, P.; Macdougall, M.; et al. Endogenous Msx1 antisense transcript: In vivo and in vitro evidences, structure, and potential involvement in skeleton development in mammals. Proc. Natl. Acad. Sci. USA 2001, 98, 7336–7341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Su, T.; Zou, C.; Luo, W.; Shi, G.; Chen, L.; Fang, C.; Li, C. Long Non-coding RNA H19 Regulates Porcine Satellite Cell Differentiation Through miR-140-5p/SOX4 and DBN1. Front. Cell Dev. Biol. 2020, 8, 518724. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Zhou, X.; Cao, H.; Zhang, X.; Huang, K.; Li, X.; Yang, G.; Shi, X. miR-324-5p Inhibits C2C12 cell Differentiation and Promotes Intramuscular Lipid Deposition through lncDUM and PM20D1. Mol. Ther. Nucleic Acids 2020, 22, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhou, L.; Zhao, Y.; Wang, H.; Sun, H. Genome-wide RNA-seq and ChIP-seq reveal Linc-YY1 function in regulating YY1/PRC2 activity during skeletal myogenesis. Genom Data 2016, 7, 247–249. [Google Scholar] [CrossRef] [Green Version]
- Dong, A.; Preusch, C.B.; So, W.K.; Lin, K.; Luan, S.; Yi, R.; Wong, J.W.; Wu, Z.; Cheung, T.H. A long noncoding RNA, LncMyoD, modulates chromatin accessibility to regulate muscle stem cell myogenic lineage progression. Proc. Natl. Acad. Sci. USA 2020, 117, 32464–32475. [Google Scholar] [CrossRef]
- Han, X.; Yang, F.; Cao, H.; Liang, Z. Malat1 regulates serum response factor through miR-133 as a competing endogenous RNA in myogenesis. FASEB J. 2015, 29, 3054–3064. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lin, Y.; Song, X.; Wu, D. LINC00857 contributes to proliferation and lymphomagenesis by regulating miR-370-3p/CBX3 axis in diffuse large B cell lymphoma. Carcinogenesis 2021. [Google Scholar] [CrossRef]
- Chen, F.; Yin, S.; Feng, Z.; Liu, C.; Lv, J.; Chen, Y.; Shen, R.; Wang, J.; Deng, Z. Knockdown of circ_NEK6 Decreased (131)I Resistance of Differentiated Thyroid Carcinoma via Regulating miR-370-3p/MYH9 Axis. Technol. Cancer Res. Treat. 2021, 20, 15330338211004950. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, P.; Zhao, X.; He, J.; Xu, Y.; Zou, Q.; Luo, J.; Shen, L.; Gu, H.; Tang, Q.; et al. MicroRNA-351-5p mediates skeletal myogenesis by directly targeting lactamase-β and is regulated by lnc-mg. FASEB J. 2019, 33, 1911–1926. [Google Scholar] [CrossRef] [PubMed]
- Kawase, A.; Nomura, T.; Yasuda, K.; Kobayashi, N.; Hashida, M.; Takakura, Y. Disposition and gene expression characteristics in solid tumors and skeletal muscle after direct injection of naked plasmid DNA in mice. J. Pharm. Sci. 2003, 92, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Du, J.; Wang, L.; Niu, L.; Zhao, Y.; Tang, G.; Jiang, Y.; Shuai, S.; Bai, L.; Li, X.; et al. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother. 2018, 108, 531–539. [Google Scholar] [CrossRef]
- Gong, C.; Li, Z.; Ramanujan, K.; Clay, I.; Zhang, Y.; Lemire-Brachat, S.; Glass, D.J. A Long Non-coding RNA, LncMyoD, Regulates Skeletal Muscle Differentiation by Blocking IMP2-Mediated mRNA Translation. Dev. Cell 2015, 34, 181–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajbakhsh, S.; Borello, U.; Vivarelli, E.; Kelly, R.; Papkoff, J.; Duprez, D.; Buckingham, M.; Cossu, G. Differential activation of Myf5 and MyoD by different Wnts in explants of mouse paraxial mesoderm and the later activation of myogenesis in the absence of Myf5. Development 1998, 125, 4155–4162. [Google Scholar]
- Rawls, A.; Valdez, M.R.; Zhang, W.; Richardson, J.A.; Klein, W.H.; Olson, E.N. Overlapping functions of the myogenic bHLH genes MRF4 and MyoD revealed in double mutant mice. Development 1998, 125, 2349–2358. [Google Scholar]
- Weintraub, H. The MyoD family and myogenesis: Redundancy, networks, and thresholds. Cell 1993, 75, 1241–1244. [Google Scholar] [CrossRef]
- Benatti, R.O.; Melo, A.M.; Borges, F.O.; Ignacio-Souza, L.M.; Simino, L.A.P.; Milanski, M.; Velloso, L.A.; Torsoni, M.A.; Torsoni, A.S. Maternal high-fat diet consumption modulates hepatic lipid metabolism and microRNA-122 (miR-122) and microRNA-370 (miR-370) expression in offspring. Br. J. Nutr. 2014, 111, 2112–2122. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, G.; Yu, C.; Shen, Z.; Xu, C.; Feng, Z.; Zhang, X.; Li, Y. A role of microRNA-370 in hepatic ischaemia-reperfusion injury by targeting transforming growth factor-β receptor II. Liver Int. 2014, 35, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Xin, S.; Weng, L.; Xu, J.; Du, W. The role of RBF in developmentally regulated cell proliferation in the eye disc and in Cyclin D/Cdk4 induced cellular growth. Development 2002, 129, 1345–1356. [Google Scholar]
- Hinds, P.W. A Confederacy of Kinases: Cdk2 and Cdk4 Conspire to Control Embryonic Cell Proliferation. Mol. Cell 2006, 22, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, Y.; Shen, L.; Luo, J.; Lei, H.; Zhang, P.; Pu, Q.; Liu, Y.; Shuai, S.; Li, Q.; et al. Effect of miR-143-3p on C2C12 myoblast differentiation. Biosci. Biotechnol. Biochem. 2016, 80, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Maldegem, B.T.; Duran, M.; Wanders, R.J.A.; Niezen-Koning, K.E.; Hogeveen, M.; Ijlst, L.; Waterham, H.R.; Wijburg, F.A. Clinical, Biochemical, and Genetic Heterogeneity in Short-Chain Acyl-Coenzyme A Dehydrogenase Deficiency. JAMA 2006, 296, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Du, J.; Guo, X.; Wu, S.; He, J.; Li, X.; Shen, L.; Chen, L.; Li, B.; Zhang, J.; et al. LncMyoD Promotes Skeletal Myogenesis and Regulates Skeletal Muscle Fiber-Type Composition by Sponging miR-370-3p. Genes 2021, 12, 589. https://doi.org/10.3390/genes12040589
Zhang P, Du J, Guo X, Wu S, He J, Li X, Shen L, Chen L, Li B, Zhang J, et al. LncMyoD Promotes Skeletal Myogenesis and Regulates Skeletal Muscle Fiber-Type Composition by Sponging miR-370-3p. Genes. 2021; 12(4):589. https://doi.org/10.3390/genes12040589
Chicago/Turabian StyleZhang, Peiwen, Jingjing Du, Xinyu Guo, Shuang Wu, Jin He, Xinrong Li, Linyuan Shen, Lei Chen, Bohong Li, Jingjun Zhang, and et al. 2021. "LncMyoD Promotes Skeletal Myogenesis and Regulates Skeletal Muscle Fiber-Type Composition by Sponging miR-370-3p" Genes 12, no. 4: 589. https://doi.org/10.3390/genes12040589
APA StyleZhang, P., Du, J., Guo, X., Wu, S., He, J., Li, X., Shen, L., Chen, L., Li, B., Zhang, J., Xie, Y., Niu, L., Jiang, D., Li, X., Zhang, S., & Zhu, L. (2021). LncMyoD Promotes Skeletal Myogenesis and Regulates Skeletal Muscle Fiber-Type Composition by Sponging miR-370-3p. Genes, 12(4), 589. https://doi.org/10.3390/genes12040589