
Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
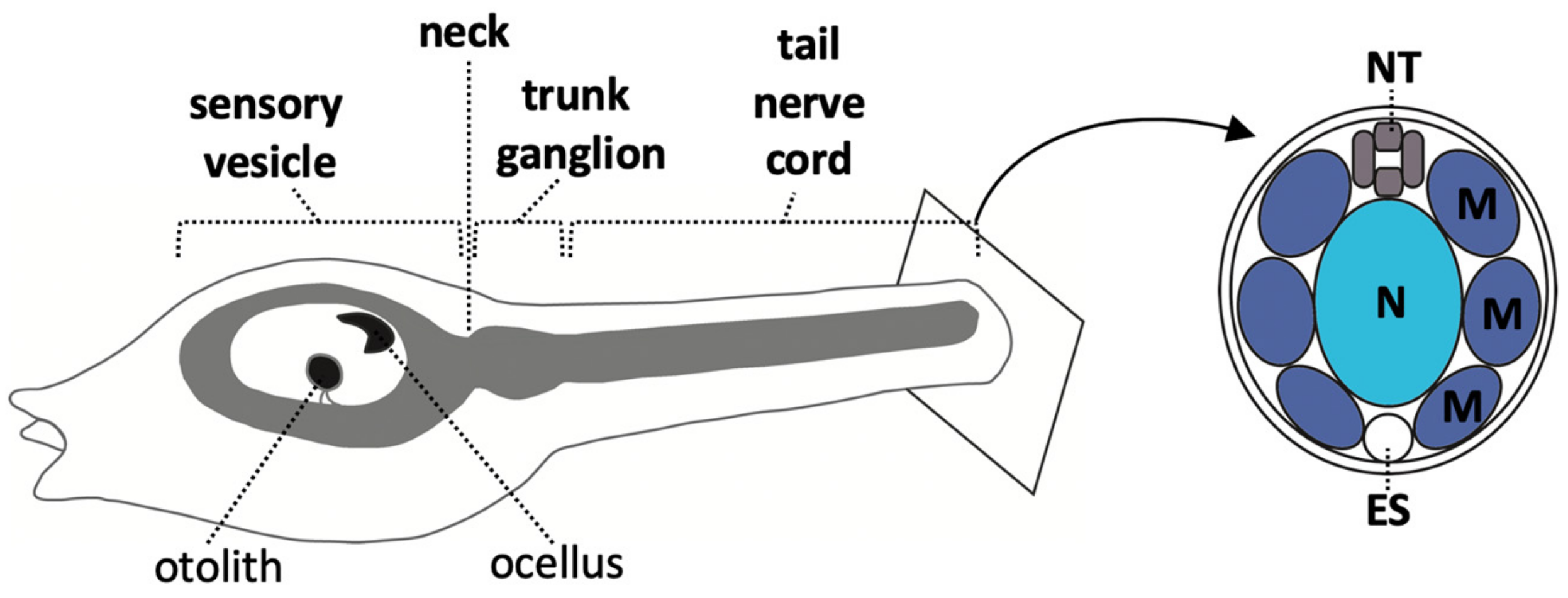
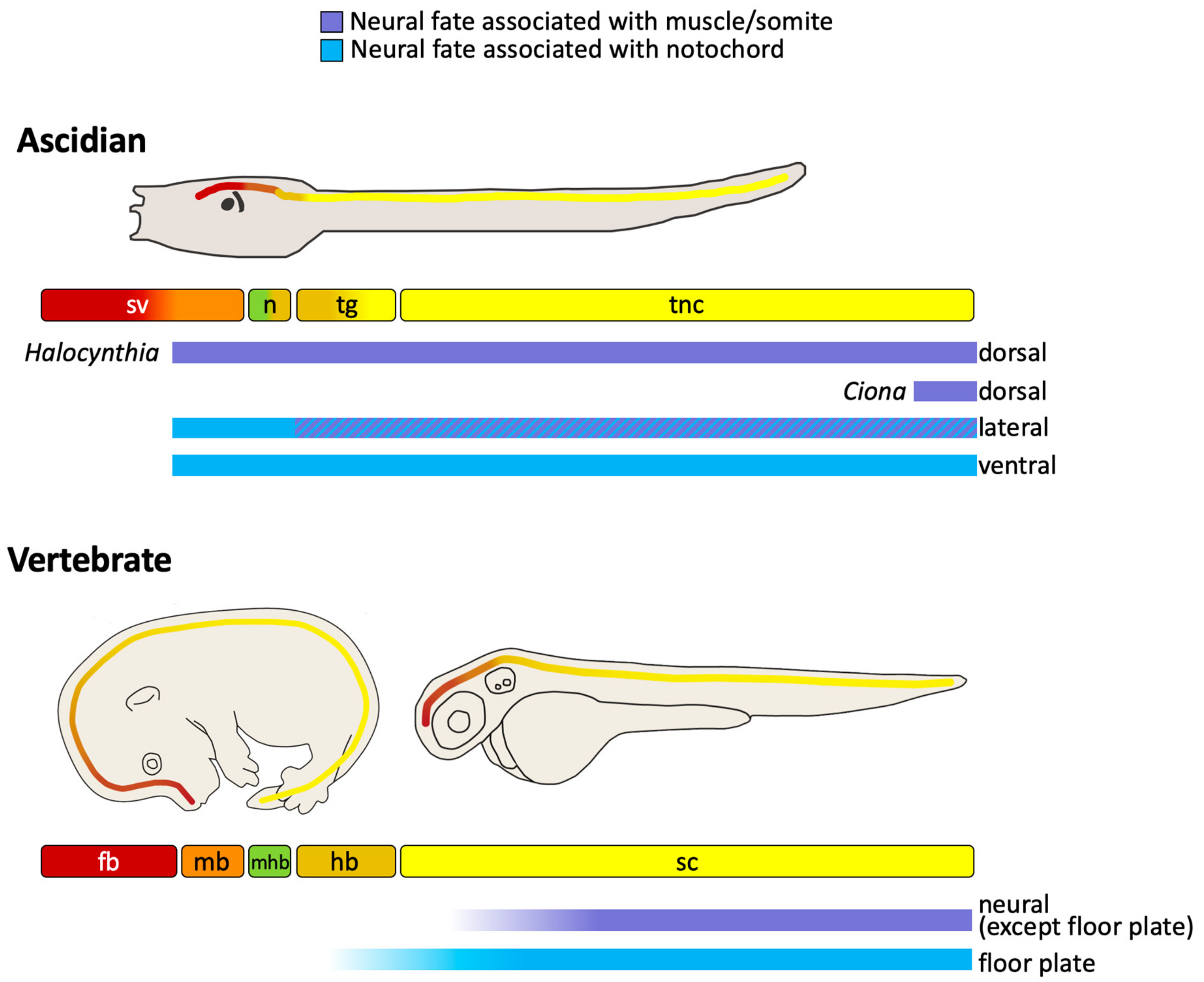
2. The Ascidian Larval CNS
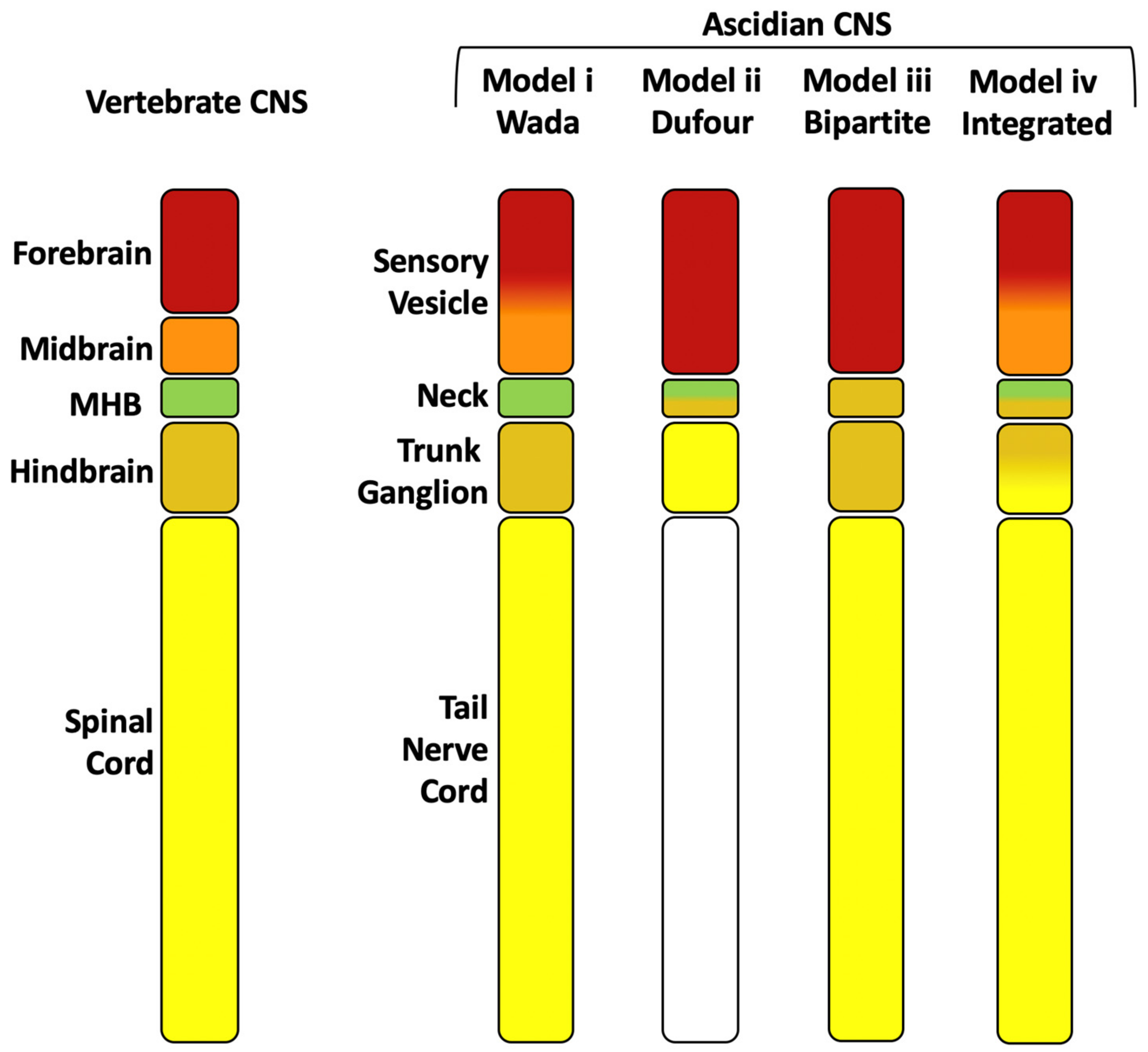
3. Anterior-Posterior Regionalization of the Chordate Neural Tube
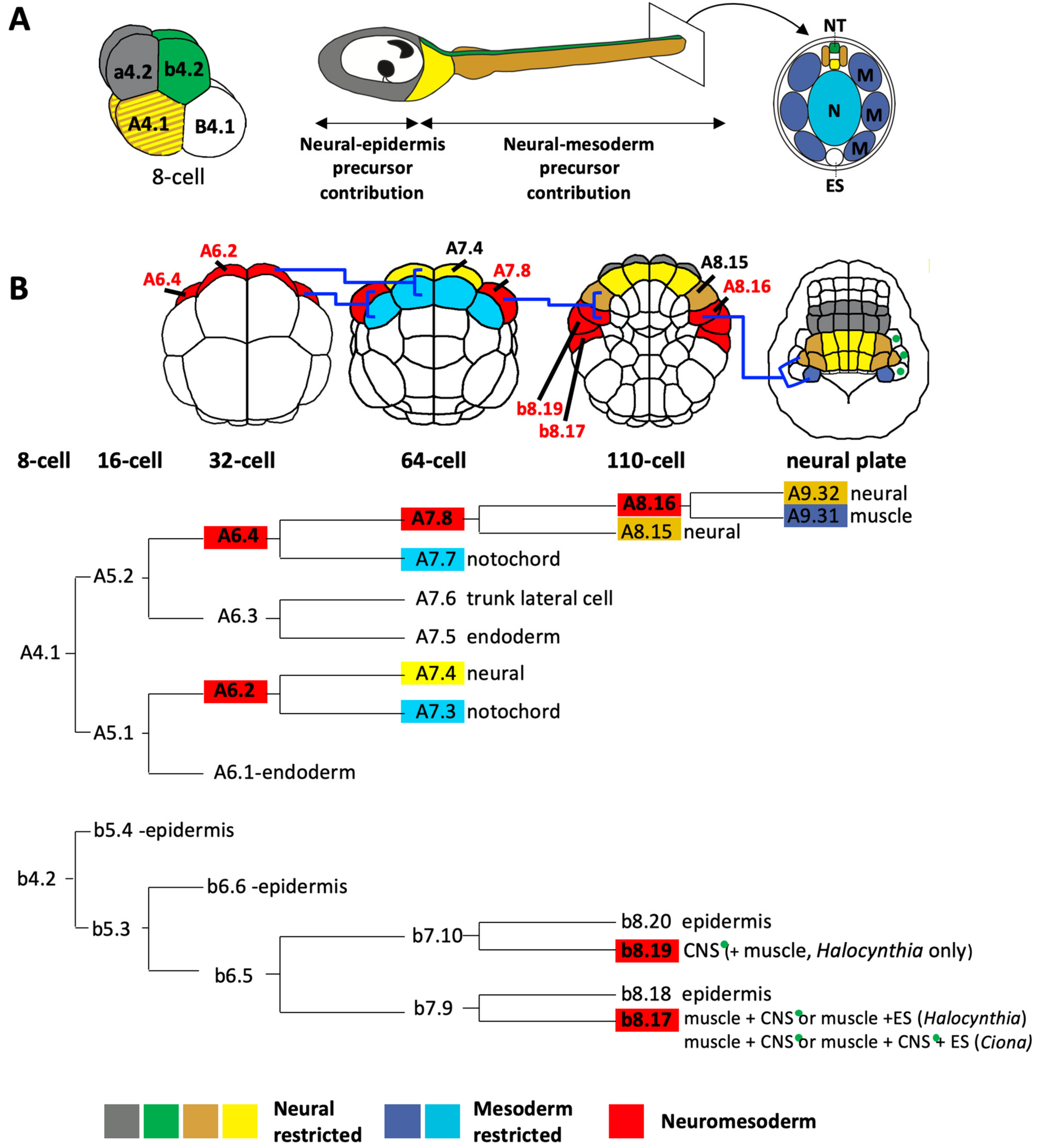
4. A Large Part of the Ascidian CNS Is Derived from Bi-Fated Neuromesodermal Precursors
5. Evidence of Neural-Mesoderm Lineages in Vertebrates
5.1. Neural-Somite Fate Choices
5.2. Neural-Notochord Fate Choices
5.3. A Note on the Historical Controversy Surrounding Floor Plate Induction in Chick
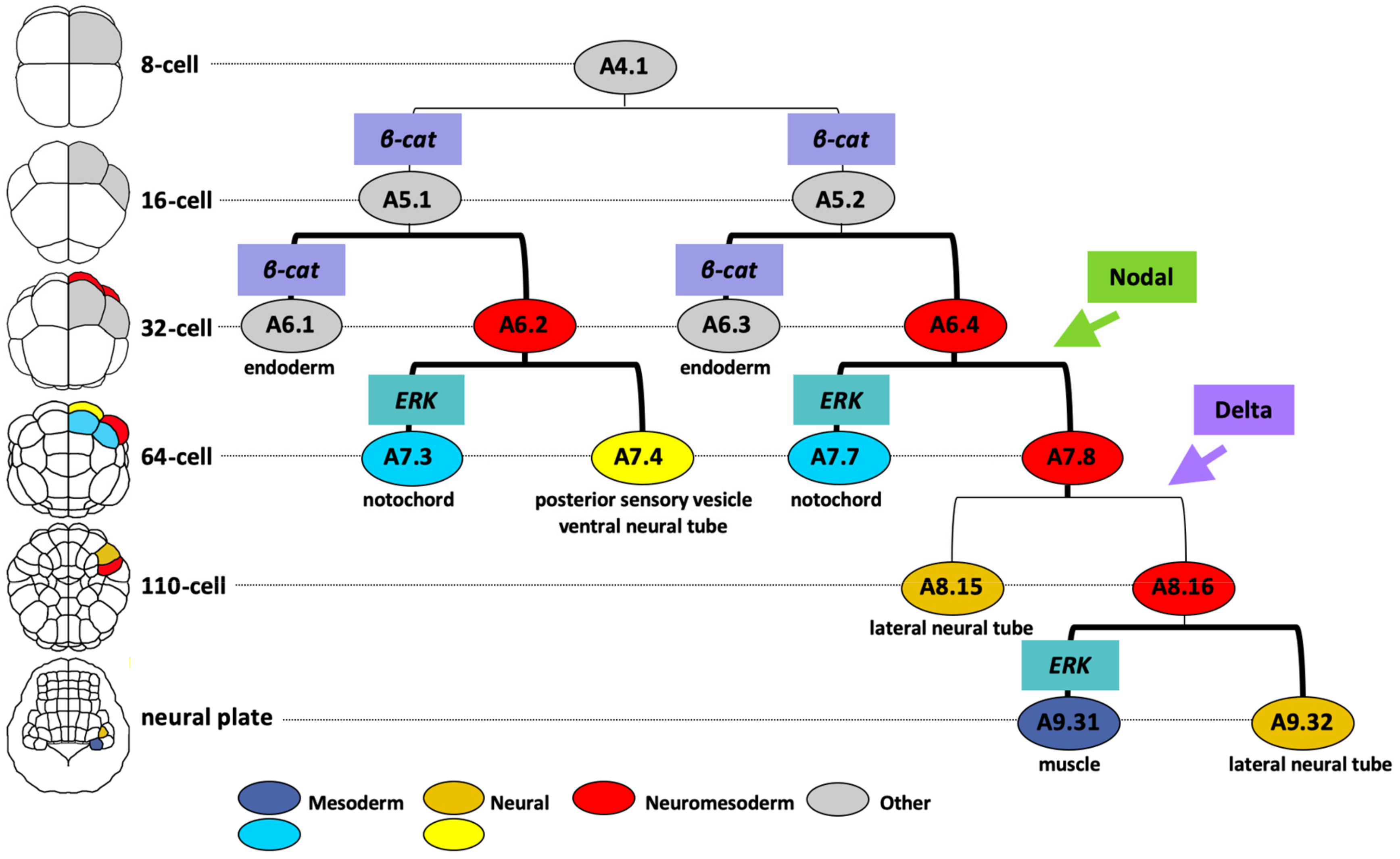
6. Molecular Mechanisms of Neuromesoderm Fate Segregation in Ascidians
7. A Common Origin for Neuromesoderm Precursors at the Base of Chordates?
7.1. Neural-Muscle Precursors
7.2. Notochord-Neural Precursors
7.3. Ascidian b-Lineage Cells: Neural, Muscle, Endoderm
8. Conclusions
9. C. robusta Unique Gene IDs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and Not Cephalochordates Are the Closest Living Relatives of Vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Minc, N.; Salez, G.; Aicha, S.B.; Bekkouche, F.; Hebras, C.; Besnardeau, L.; McDougall, A. The Invariant Cleavage Pattern Displayed by Ascidian Embryos Depends on Spindle Positioning along the Cell’s Longest Axis in the Apical Plane and Relies on Asynchronous Cell Divisions. eLife 2017, 6, e19290. [Google Scholar] [CrossRef] [Green Version]
- Guignard, L.; Fiúza, U.-M.; Leggio, B.; Laussu, J.; Faure, E.; Michelin, G.; Biasuz, K.; Hufnagel, L.; Malandain, G.; Godin, C.; et al. Contact Area–Dependent Cell Communication and the Morphological Invariance of Ascidian Embryogenesis. Science 2020, 369, eaar5663. [Google Scholar] [CrossRef] [PubMed]
- Hudson, C.; Yasuo, H. Similarity and Diversity in Mechanisms of Muscle Fate Induction between Ascidian Species. Biol. Cell 2008, 100, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, P. Unfolding a Chordate Developmental Program, One Cell at a Time: Invariant Cell Lineages, Short-Range Inductions and Evolutionary Plasticity in Ascidians. Dev. Biol. 2009, 332, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, P. Evolutionary Crossroads in Developmental Biology: The Tunicates. Development 2011, 138, 2143–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, E.K.; Stolfi, A. Developmental System Drift in Motor Ganglion Patterning between Distantly Related Tunicates. Evodevo 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, H. Cell Lineage Analysis in Ascidian Embryos by Intracellular Injection of a Tracer Enzyme. III. Up to the Tissue Restricted Stage. Dev. Biol. 1987, 121, 526–541. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Stolfi, A. Regulation and Evolution of Muscle Development in Tunicates. EvoDevo 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolfi, A.; Lowe, E.K.; Racioppi, C.; Ristoratore, F.; Brown, C.T.; Swalla, B.J.; Christiaen, L. Divergent Mechanisms Regulate Conserved Cardiopharyngeal Development and Gene Expression in Distantly Related Ascidians. Elife 2014, 3, e03728. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N.; Rokhsar, D.; Nishikawa, T. Chordate Evolution and the Three-Phylum System. Proc. Biol. Sci. 2014, 281, 20141729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, A.; Nishida, H. Distinct Parameters Are Involved in Controlling the Number of Rounds of Cell Division in Each Tissue during Ascidian Embryogenesis. J. Exp. Zool. 1999, 284, 379–391. [Google Scholar] [CrossRef]
- Navarrete, I.A.; Levine, M. Nodal and FGF Coordinate Ascidian Neural Tube Morphogenesis. Development 2016, 143, 4665–4675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicol, D.; Meinertzhagen, I.A. Development of the Central Nervous System of the Larva of the Ascidian, Ciona Intestinalis L. II. Neural Plate Morphogenesis and Cell Lineages during Neurulation. Dev. Biol. 1988, 130, 737–766. [Google Scholar] [CrossRef]
- Henrique, D.; Abranches, E.; Verrier, L.; Storey, K.G. Neuromesodermal Progenitors and the Making of the Spinal Cord. Development 2015, 142, 2864–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, C.D.; Charité, J.; Deschamps, J.; Duboule, D.; Durston, A.J.; Kmita, M.; Nicolas, J.-F.; Palmeirim, I.; Smith, J.C.; Wolpert, L. Head-Tail Patterning of the Vertebrate Embryo: One, Two or Many Unresolved Problems? Int. J. Dev. Biol. 2006, 50, 3–15. [Google Scholar] [CrossRef]
- Gouti, M.; Metzis, V.; Briscoe, J. The Route to Spinal Cord Cell Types: A Tale of Signals and Switches. Trends Genet. 2015, 31, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Takada, S.; Takemoto, T. Axial Level-Dependent Molecular and Cellular Mechanisms Underlying the Genesis of the Embryonic Neural Plate. Dev. Growth Differ. 2016, 58, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Al Anber, A.; Martin, B.L. Transformation of a Neural Activation and Patterning Model. EMBO Rep. 2019, 20, e48060. [Google Scholar] [CrossRef] [PubMed]
- Attardi, A.; Fulton, T.; Florescu, M.; Shah, G.; Muresan, L.; Lenz, M.O.; Lancaster, C.; Huisken, J.; van Oudenaarden, A.; Steventon, B. Neuromesodermal Progenitors Are a Conserved Source of Spinal Cord with Divergent Growth Dynamics. Development 2018, 145, dev166728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, R.K. Cell Movements and Cell Fate during Zebrafish Gastrulation. Development 1992, 116, 65–73. [Google Scholar]
- Hashimoto, H.; Robin, F.B.; Sherrard, K.M.; Munro, E.M. Sequential Contraction and Exchange of Apical Junctions Drives Zippering and Neural Tube Closure in a Simple Chordate. Dev. Cell 2015, 32, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasakura, Y.; Mita, K.; Ogura, Y.; Horie, T. Ascidians as Excellent Chordate Models for Studying the Development of the Nervous System during Embryogenesis and Metamorphosis. Dev. Growth Differ. 2012, 54, 420–437. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G. Cell Type and Function of Neurons in the Ascidian Nervous System. Dev. Growth Differ. 2009, 51, 207–220. [Google Scholar] [CrossRef]
- Hudson, C. The Central Nervous System of Ascidian Larvae. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 538–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Stolfi, A.; Levine, M.; Satou, Y. Gene Regulatory Networks Underlying the Compartmentalization of the Ciona Central Nervous System. Development 2009, 136, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, K.; Meinertzhagen, I.A. Neuronal Identity: The Neuron Types of a Simple Chordate Sibling, the Tadpole Larva of Ciona Intestinalis. Curr. Opin. Neurobiol. 2019, 56, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. The CNS Connectome of a Tadpole Larva of Ciona Intestinalis (L.) Highlights Sidedness in the Brain of a Chordate Sibling. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Lemaire, L.A.; Wang, W.; Yoon, P.H.; Choi, Y.A.; Parsons, L.R.; Matese, J.C.; Wang, W.; Levine, M.; Chen, K. Comprehensive Single-Cell Transcriptome Lineages of a Proto-Vertebrate. Nature 2019, 571, 349–354. [Google Scholar] [CrossRef]
- Horie, R.; Hazbun, A.; Chen, K.; Cao, C.; Levine, M.; Horie, T. Shared Evolutionary Origin of Vertebrate Neural Crest and Cranial Placodes. Nature 2018, 560, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Horie, R.; Chen, K.; Cao, C.; Nakagawa, M.; Kusakabe, T.G.; Satoh, N.; Sasakura, Y.; Levine, M. Regulatory Cocktail for Dopaminergic Neurons in a Protovertebrate Identified by Whole-Embryo Single-Cell Transcriptomics. Genes Dev. 2018, 32, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Wang, W.; Stolfi, A. Single-Cell Transcriptome Profiling of the Ciona Larval Brain. Dev. Biol. 2019, 448, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Sladitschek, H.L.; Fiuza, U.-M.; Pavlinic, D.; Benes, V.; Hufnagel, L.; Neveu, P.A. MorphoSeq: Full Single-Cell Transcriptome Dynamics Up to Gastrulation in a Chordate. Cell 2020, 181, 922–935.e21. [Google Scholar] [CrossRef]
- Borba, C.; Kourakis, M.J.; Schwennicke, S.; Brasnic, L.; Smith, W.C. Visual Processing and Fold-Change Detection by the Larva of the Simple Chordate Ciona. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bostwick, M.; Smith, E.L.; Borba, C.; Newman-Smith, E.; Guleria, I.; Kourakis, M.J.; Smith, W.C. Antagonistic Inhibitory Circuits Integrate Visual and Gravitactic Behaviors. Curr. Biol. 2020, 30, 600–609.e2. [Google Scholar] [CrossRef] [Green Version]
- Kourakis, M.J.; Borba, C.; Zhang, A.; Newman-Smith, E.; Salas, P.; Manjunath, B.; Smith, W.C. Parallel Visual Circuitry in a Basal Chordate. Elife 2019, 8. [Google Scholar] [CrossRef]
- Oonuma, K.; Kusakabe, T.G. Spatio-Temporal Regulation of Rx and Mitotic Patterns Shape the Eye-Cup of the Photoreceptor Cells in Ciona. Dev. Biol. 2019, 445, 245–255. [Google Scholar] [CrossRef]
- Oonuma, K.; Tanaka, M.; Nishitsuji, K.; Kato, Y.; Shimai, K.; Kusakabe, T.G. Revised Lineage of Larval Photoreceptor Cells in Ciona Reveals Archetypal Collaboration between Neural Tube and Neural Crest in Sensory Organ Formation. Dev. Biol. 2016, 420, 178–185. [Google Scholar] [CrossRef]
- Stolfi, A.; Ryan, K.; Meinertzhagen, I.A.; Christiaen, L. Migratory Neuronal Progenitors Arise from the Neural Plate Borders in Tunicates. Nature 2015, 527, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Shinki, R.; Ogura, Y.; Kusakabe, T.G.; Satoh, N.; Sasakura, Y. Ependymal Cells of Chordate Larvae Are Stem-like Cells That Form the Adult Nervous System. Nature 2011, 469, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Esposito, R.; Racioppi, C.; Pezzotti, M.R.; Branno, M.; Locascio, A.; Ristoratore, F.; Spagnuolo, A. The Ascidian Pigmented Sensory Organs: Structures and Developmental Programs. Genesis 2015, 53, 15–33. [Google Scholar] [CrossRef]
- Jiang, D.; Tresser, J.W.; Horie, T.; Tsuda, M.; Smith, W.C. Pigmentation in the Sensory Organs of the Ascidian Larva Is Essential for Normal Behavior. J. Exp. Biol. 2005, 208, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Sakurai, D.; Goda, M. Direct Evidence for the Role of Pigment Cells in the Brain of Ascidian Larvae by Laser Ablation. J. Exp. Biol. 2003, 206, 1409–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, H.D.; Chettouh, Z.; Deyts, C.; de Rosa, R.; Goridis, C.; Joly, J.-S.; Brunet, J.-F. Precraniate Origin of Cranial Motoneurons. Proc. Natl. Acad. Sci. USA 2006, 103, 8727–8732. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.G.; Meinertzhagen, I.A. The Central Nervous System of the Ascidian Larva: Mitotic History of Cells Forming the Neural Tube in Late Embryonic Ciona Intestinalis. Dev. Biol. 2004, 271, 239–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibboney, S.; Orvis, J.; Kim, K.; Johnson, C.J.; Martinez-Feduchi, P.; Lowe, E.K.; Sharma, S.; Stolfi, A. Effector Gene Expression Underlying Neuron Subtype-Specific Traits in the Motor Ganglion of Ciona. Dev. Biol. 2020, 458, 52–63. [Google Scholar] [CrossRef]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G.; Tsuda, M. Simple Motor System of the Ascidian Larva: Neuronal Complex Comprising Putative Cholinergic and GABAergic/Glycinergic Neurons. Zool. Sci. 2010, 27, 181–190. [Google Scholar] [CrossRef]
- Stolfi, A.; Levine, M. Neuronal Subtype Specification in the Spinal Cord of a Protovertebrate. Development 2011, 138, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, J.H.; Meinertzhagen, I.A. Neurons of the Ascidian Larval Nervous System in Ciona Intestinalis: I. Central Nervous System. J. Comp. Neurol. 2007, 501, 316–334. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Horie, T.; Ichinose, A.; Sasakura, Y.; Yasuo, H.; Kusakabe, T.G. Cell Lineage and Cis-Regulation for a Unique GABAergic/Glycinergic Neuron Type in the Larval Nerve Cord of the Ascidian Ciona Intestinalis. Dev. Growth Differ. 2012, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Nicol, D.; Meinertzhagen, I.A. Cell Counts and Maps in the Larval Central Nervous System of the Ascidian Ciona Intestinalis (L.). J. Comp. Neurol. 1991, 309, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, R146–R152. [Google Scholar] [CrossRef] [Green Version]
- Kusakabe, T.G. Identifying Vertebrate Brain Prototypes in Deuterostomes. In Brain Evolution by Design; Diversity and Commonality in Animals; Shigeno, S., Murakami, Y., Nomura, T., Eds.; Springer Japan: Tokyo, Japan, 2017; pp. 153–186. ISBN 978-4-431-56467-6. [Google Scholar]
- Lacalli, T.C. Prospective Protochordate Homologs of Vertebrate Midbrain and MHB, with Some Thoughts on MHB Origins. Int. J. Biol. Sci. 2006, 2, 104–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, B.; Shimeld, S.M. Evolution of Vertebrate Spinal Cord Patterning. Dev. Dyn. 2019, 248, 1028–1043. [Google Scholar] [CrossRef] [Green Version]
- Shimeld, S.M.; Holland, P.W. Vertebrate Innovations. Proc. Natl. Acad. Sci. USA 2000, 97, 4449–4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cañestro, C.; Bassham, S.; Postlethwait, J. Development of the Central Nervous System in the Larvacean Oikopleura Dioica and the Evolution of the Chordate Brain. Dev. Biol. 2005, 285, 298–315. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Saiga, H. Dynamic Change in the Expression of Developmental Genes in the Ascidian Central Nervous System: Revisit to the Tripartite Model and the Origin of the Midbrain-Hindbrain Boundary Region. Dev. Biol. 2007, 312, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.S.; Satoh, N.; Satou, Y. Region Specific Gene Expressions in the Central Nervous System of the Ascidian Embryo. Mech. Dev. 2002, 119 (Suppl. 1), S275–S277. [Google Scholar] [CrossRef]
- Liu, B.; Satou, Y. The Genetic Program to Specify Ectodermal Cells in Ascidian Embryos. Dev. Growth Differ. 2020, 62, 301–310. [Google Scholar] [CrossRef]
- Satoh, N. The Ascidian Tadpole Larva: Comparative Molecular Development and Genomics. Nat. Rev. Genet. 2003, 4, 285–295. [Google Scholar] [CrossRef]
- Takahashi, T.; Holland, P.W.H. Amphioxus and Ascidian Dmbx Homeobox Genes Give Clues to the Vertebrate Origins of Midbrain Development. Development 2004, 131, 3285–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, H.; Satoh, N. Patterning the Protochordate Neural Tube. Curr. Opin. Neurobiol. 2001, 11, 16–21. [Google Scholar] [CrossRef]
- Wada, H.; Saiga, H.; Satoh, N.; Holland, P.W. Tripartite Organization of the Ancestral Chordate Brain and the Antiquity of Placodes: Insights from Ascidian Pax-2/5/8, HOX and Otx Genes. Development 1998, 125, 1113–1122. [Google Scholar]
- Nishino, A.; Okamura, Y.; Piscopo, S.; Brown, E.R. A Glycine Receptor Is Involved in the Organization of Swimming Movements in an Invertebrate Chordate. BMC Neurosci. 2010, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, A.; Baba, S.A.; Okamura, Y. A Mechanism for Graded Motor Control Encoded in the Channel Properties of the Muscle ACh Receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 2599–2604. [Google Scholar] [CrossRef] [Green Version]
- Stolfi, A.; Wagner, E.; Taliaferro, J.M.; Chou, S.; Levine, M. Neural Tube Patterning by Ephrin, FGF and Notch Signaling Relays. Development 2011, 138, 5429–5439. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. Circuit Homology between Decussating Pathways in the Ciona Larval CNS and the Vertebrate Startle-Response Pathway. Curr. Biol. 2017, 27, 721–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragon, F.; Pujades, C. FGF Signaling Controls Caudal Hindbrain Specification through Ras-ERK1/2 Pathway. BMC Dev. Biol. 2009, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maves, L.; Jackman, W.; Kimmel, C.B. FGF3 and FGF8 Mediate a Rhombomere 4 Signaling Activity in the Zebrafish Hindbrain. Development 2002, 129, 3825–3837. [Google Scholar]
- Walshe, J.; Maroon, H.; McGonnell, I.M.; Dickson, C.; Mason, I. Establishment of Hindbrain Segmental Identity Requires Signaling by FGF3 and FGF8. Curr. Biol. 2002, 12, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Conklin, E.G. The Organisation and Cell Lineage of the Ascidian Egg; The Academy of Natural Sciences: Philadelphia, PA, USA, 1905; pp. 1–119. [Google Scholar]
- Nicol, D.; Meinertzhagen, I.A. Development of the Central Nervous System of the Larva of the Ascidian, Ciona Intestinalis L. I. The Early Lineages of the Neural Plate. Dev. Biol. 1988, 130, 721–736. [Google Scholar] [CrossRef]
- Abitua, P.B.; Gainous, T.B.; Kaczmarczyk, A.N.; Winchell, C.J.; Hudson, C.; Kamata, K.; Nakagawa, M.; Tsuda, M.; Kusakabe, T.G.; Levine, M. The Pre-Vertebrate Origins of Neurogenic Placodes. Nature 2015, 524, 462–465. [Google Scholar] [CrossRef]
- Nishida, H. Determinative Mechanisms in Secondary Muscle Lineages of Ascidian Embryos: Development of Muscle-Specific Features in Isolated Muscle Progenitor Cells. Development 1990, 108, 559–568. [Google Scholar]
- Bertrand, V.; Hudson, C.; Caillol, D.; Popovici, C.; Lemaire, P. Neural Tissue in Ascidian Embryos Is Induced by FGF9/16/20, Acting via a Combination of Maternal GATA and Ets Transcription Factors. Cell 2003, 115, 615–627. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Lemaire, P. Induction of Anterior Neural Fates in the Ascidian Ciona Intestinalis. Mech. Dev. 2001, 100, 189–203. [Google Scholar] [CrossRef]
- Hudson, C.; Darras, S.; Caillol, D.; Yasuo, H.; Lemaire, P. A Conserved Role for the MEK Signalling Pathway in Neural Tissue Specification and Posteriorisation in the Invertebrate Chordate, the Ascidian Ciona Intestinalis. Development 2003, 130, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsuka, Y.; Matsumoto, J.; Katsuyama, Y.; Okamura, Y. Nodal Signaling Regulates Specification of Ascidian Peripheral Neurons through Control of the BMP Signal. Development 2014, 141, 3889–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.; Stolfi, A.; Gi Choi, Y.; Levine, M. Islet Is a Key Determinant of Ascidian Palp Morphogenesis. Development 2014, 141, 3084–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Matsuoka, T.; Satou, Y. A Time Delay Gene Circuit Is Required for Palp Formation in the Ascidian Embryo. Development 2013, 140, 4703–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, N.; Satou, Y. Multiple Signaling Pathways Coordinate to Induce a Threshold Response in a Chordate Embryo. PLoS Genet. 2013, 9, e1003818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.; Levine, M. FGF Signaling Establishes the Anterior Border of the Ciona Neural Tube. Development 2012, 139, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Satou, Y. Differential Temporal Control of Foxa.a and Zic-r.b Specifies Brain versus Notochord Fate in the Ascidian Embryo. Development 2017, 144, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Satoh, N. Developmental Genomics of Ascidians; Wiley-Blackwell: Hoboken, NJ, USA, 2014; ISBN 978-1-118-65618-1. [Google Scholar]
- Pasini, A.; Amiel, A.; Rothbächer, U.; Roure, A.; Lemaire, P.; Darras, S. Formation of the Ascidian Epidermal Sensory Neurons: Insights into the Origin of the Chordate Peripheral Nervous System. PLoS Biol. 2006, 4, e225. [Google Scholar] [CrossRef] [PubMed]
- Minokawa, T.; Yagi, K.; Makabe, K.W.; Nishida, H. Binary Specification of Nerve Cord and Notochord Cell Fates in Ascidian Embryos. Development 2001, 128, 2007–2017. [Google Scholar] [PubMed]
- Picco, V.; Hudson, C.; Yasuo, H. Ephrin-Eph Signalling Drives the Asymmetric Division of Notochord/Neural Precursors in Ciona Embryos. Development 2007, 134, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Lotito, S.; Yasuo, H. Sequential and Combinatorial Inputs from Nodal, Delta2/Notch and FGF/MEK/ERK Signalling Pathways Establish a Grid-like Organisation of Distinct Cell Identities in the Ascidian Neural Plate. Development 2007, 134, 3527–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, P.; Smith, W.C.; Nishida, H. Ascidians and the Plasticity of the Chordate Developmental Program. Curr. Biol. 2008, 18, R620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuoka, M.; Kumano, G.; Nishida, H. FGF9/16/20 and Wnt-5alpha Signals Are Involved in Specification of Secondary Muscle Fate in Embryos of the Ascidian, Halocynthia Roretzi. Dev. Genes Evol. 2007, 217, 515–527. [Google Scholar] [CrossRef]
- Nishida, H.; Stach, T. Cell Lineages and Fate Maps in Tunicates: Conservation and Modification. Zoolog. Sci. 2014, 31, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Stach, T.; Winter, J.; Bouquet, J.-M.; Chourrout, D.; Schnabel, R. Embryology of a Planktonic Tunicate Reveals Traces of Sessility. Proc. Natl. Acad. Sci. USA 2008, 105, 7229–7234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cajal, M.; Lawson, K.A.; Hill, B.; Moreau, A.; Rao, J.; Ross, A.; Collignon, J.; Camus, A. Clonal and Molecular Analysis of the Prospective Anterior Neural Boundary in the Mouse Embryo. Development 2012, 139, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Metzis, V.; Steinhauser, S.; Pakanavicius, E.; Gouti, M.; Stamataki, D.; Ivanovitch, K.; Watson, T.; Rayon, T.; Mousavy Gharavy, S.N.; Lovell-Badge, R.; et al. Nervous System Regionalization Entails Axial Allocation before Neural Differentiation. Cell 2018, 175, 1105–1118.e17. [Google Scholar] [CrossRef] [Green Version]
- Tzouanacou, E.; Wegener, A.; Wymeersch, F.J.; Wilson, V.; Nicolas, J.-F. Redefining the Progression of Lineage Segregations during Mammalian Embryogenesis by Clonal Analysis. Dev. Cell 2009, 17, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouti, M.; Tsakiridis, A.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. In Vitro Generation of Neuromesodermal Progenitors Reveals Distinct Roles for Wnt Signalling in the Specification of Spinal Cord and Paraxial Mesoderm Identity. PLoS Biol. 2014, 12, e1001937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimelman, D. Tales of Tails (and Trunks). In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 116, pp. 517–536. ISBN 978-0-12-802956-5. [Google Scholar]
- Kimelman, D.; Griffin, K.J. Vertebrate Mesendoderm Induction and Patterning. Curr. Opin. Genet. Dev. 2000, 10, 350–356. [Google Scholar] [CrossRef]
- Martin, B.L.; Kimelman, D. Canonical Wnt Signaling Dynamically Controls Multiple Stem Cell Fate Decisions during Vertebrate Body Formation. Dev. Cell 2012, 22, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steventon, B.; Martinez Arias, A. Evo-Engineering and the Cellular and Molecular Origins of the Vertebrate Spinal Cord. Dev. Biol. 2017, 432, 3–13. [Google Scholar] [CrossRef]
- Brown, J.M.; Storey, K.G. A Region of the Vertebrate Neural Plate in Which Neighbouring Cells Can Adopt Neural or Epidermal Fates. Curr. Biol. 2000, 10, 869–872. [Google Scholar] [CrossRef] [Green Version]
- Cambray, N.; Wilson, V. Axial Progenitors with Extensive Potency Are Localised to the Mouse Chordoneural Hinge. Development 2002, 129, 4855–4866. [Google Scholar]
- Cambray, N.; Wilson, V. Two Distinct Sources for a Population of Maturing Axial Progenitors. Development 2007, 134, 2829–2840. [Google Scholar] [CrossRef] [Green Version]
- Catala, M.; Teillet, M.A.; De Robertis, E.M.; Le Douarin, M.L. A Spinal Cord Fate Map in the Avian Embryo: While Regressing, Hensen’s Node Lays down the Notochord and Floor Plate Thus Joining the Spinal Cord Lateral Walls. Development 1996, 122, 2599–2610. [Google Scholar] [PubMed]
- Tam, P.P.; Beddington, R.S. The Formation of Mesodermal Tissues in the Mouse Embryo during Gastrulation and Early Organogenesis. Development 1987, 99, 109–126. [Google Scholar]
- Kimmel, C.B.; Warga, R.M.; Schilling, T.F. Origin and Organization of the Zebrafish Fate Map. Development 1990, 108, 581–594. [Google Scholar]
- Ciruna, B.G.; Schwartz, L.; Harpal, K.; Yamaguchi, T.P.; Rossant, J. Chimeric Analysis of Fibroblast Growth Factor Receptor-1 (Fgfr1) Function: A Role for FGFR1 in Morphogenetic Movement through the Primitive Streak. Development 1997, 124, 2829–2841. [Google Scholar] [PubMed]
- Yoshikawa, Y.; Fujimori, T.; McMahon, A.P.; Takada, S. Evidence That Absence of Wnt-3a Signaling Promotes Neuralization Instead of Paraxial Mesoderm Development in the Mouse. Dev. Biol. 1997, 183, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, D.L.; Papaioannou, V.E. Three Neural Tubes in Mouse Embryos with Mutations in the T-Box Gene Tbx6. Nature 1998, 391, 695–697. [Google Scholar] [CrossRef]
- Goto, H.; Kimmey, S.C.; Row, R.H.; Matus, D.Q.; Martin, B.L. FGF and Canonical Wnt Signaling Cooperate to Induce Paraxial Mesoderm from Tailbud Neuromesodermal Progenitors through Regulation of a Two-Step Epithelial to Mesenchymal Transition. Development 2017, 144, 1412–1424. [Google Scholar] [CrossRef] [Green Version]
- Gouti, M.; Delile, J.; Stamataki, D.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. A Gene Regulatory Network Balances Neural and Mesoderm Specification during Vertebrate Trunk Development. Dev. Cell 2017, 41, 243–261.e7. [Google Scholar] [CrossRef]
- Wilson, V.; Olivera-Martinez, I.; Storey, K.G. Stem Cells, Signals and Vertebrate Body Axis Extension. Development 2009, 136, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wymeersch, F.J.; Huang, Y.; Blin, G.; Cambray, N.; Wilkie, R.; Wong, F.C.K.; Wilson, V. Position-Dependent Plasticity of Distinct Progenitor Types in the Primitive Streak. Elife 2016, 5, e10042. [Google Scholar] [CrossRef]
- Sambasivan, R.; Steventon, B. Neuromesodermal Progenitors: A Basis for Robust Axial Patterning in Development and Evolution. Front. Cell Dev. Biol. 2021, 8, 607516. [Google Scholar] [CrossRef] [PubMed]
- Bouldin, C.M.; Snelson, C.D.; Farr, G.H.; Kimelman, D. Restricted Expression of Cdc25a in the Tailbud Is Essential for Formation of the Zebrafish Posterior Body. Genes Dev. 2014, 28, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Charrier, J.-B.; Lapointe, F.; Le Douarin, N.M.; Teillet, M.-A. Dual Origin of the Floor Plate in the Avian Embryo. Development 2002, 129, 4785–4796. [Google Scholar] [PubMed]
- Odenthal, J.; van Eeden, F.J.; Haffter, P.; Ingham, P.W.; Nüsslein-Volhard, C. Two Distinct Cell Populations in the Floor Plate of the Zebrafish Are Induced by Different Pathways. Dev. Biol. 2000, 219, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Placzek, M.; Briscoe, J. The Floor Plate: Multiple Cells, Multiple Signals. Nat. Rev. Neurosci. 2005, 6, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Strähle, U.; Lam, C.S.; Ertzer, R.; Rastegar, S. Vertebrate Floor-Plate Specification: Variations on Common Themes. Trends Genet. 2004, 20, 155–162. [Google Scholar] [CrossRef]
- Kinder, S.J.; Tsang, T.E.; Wakamiya, M.; Sasaki, H.; Behringer, R.R.; Nagy, A.; Tam, P.P. The Organizer of the Mouse Gastrula Is Composed of a Dynamic Population of Progenitor Cells for the Axial Mesoderm. Development 2001, 128, 3623–3634. [Google Scholar] [PubMed]
- Wilson, V.; Beddington, R.S. Cell Fate and Morphogenetic Movement in the Late Mouse Primitive Streak. Mech. Dev. 1996, 55, 79–89. [Google Scholar] [CrossRef]
- Halpern, M.E.; Ho, R.K.; Walker, C.; Kimmel, C.B. Induction of Muscle Pioneers and Floor Plate Is Distinguished by the Zebrafish No Tail Mutation. Cell 1993, 75, 99–111. [Google Scholar] [CrossRef]
- Halpern, M.E.; Thisse, C.; Ho, R.K.; Thisse, B.; Riggleman, B.; Trevarrow, B.; Weinberg, E.S.; Postlethwait, J.H.; Kimmel, C.B. Cell-Autonomous Shift from Axial to Paraxial Mesodermal Development in Zebrafish Floating Head Mutants. Development 1995, 121, 4257–4264. [Google Scholar]
- Kudoh, T.; Concha, M.L.; Houart, C.; Dawid, I.B.; Wilson, S.W. Combinatorial Fgf and Bmp Signalling Patterns the Gastrula Ectoderm into Prospective Neural and Epidermal Domains. Development 2004, 131, 3581–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latimer, A.J.; Appel, B. Notch Signaling Regulates Midline Cell Specification and Proliferation in Zebrafish. Dev. Biol. 2006, 298, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Melby, A.E.; Warga, R.M.; Kimmel, C.B. Specification of Cell Fates at the Dorsal Margin of the Zebrafish Gastrula. Development 1996, 122, 2225–2237. [Google Scholar] [PubMed]
- Shih, J.; Fraser, S.E. Distribution of Tissue Progenitors within the Shield Region of the Zebrafish Gastrula. Development 1995, 121, 2755–2765. [Google Scholar] [PubMed]
- Shih, J.; Fraser, S.E. Characterizing the Zebrafish Organizer: Microsurgical Analysis at the Early-Shield Stage. Development 1996, 122, 1313–1322. [Google Scholar] [PubMed]
- Selleck, M.A.; Stern, C.D. Fate Mapping and Cell Lineage Analysis of Hensen’s Node in the Chick Embryo. Development 1991, 112, 615–626. [Google Scholar] [PubMed]
- Kimmel, C.B.; Warga, R.M. Cell Lineage and Developmental Potential of Cells in the Zebrafish Embryo. Trends Genet. 1988, 4, 68–74. [Google Scholar] [CrossRef]
- Amacher, S.L.; Draper, B.W.; Summers, B.R.; Kimmel, C.B. The Zebrafish T-Box Genes No Tail and Spadetail Are Required for Development of Trunk and Tail Mesoderm and Medial Floor Plate. Development 2002, 129, 3311–3323. [Google Scholar] [PubMed]
- Row, R.H.; Tsotras, S.R.; Goto, H.; Martin, B.L. The Zebrafish Tailbud Contains Two Independent Populations of Midline Progenitor Cells That Maintain Long-Term Germ Layer Plasticity and Differentiate in Response to Local Signaling Cues. Development 2016, 143, 244–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, K.; Rubinstein, A.L.; Cheng, A.M.; Liang, J.O.; Fekany, K.; Solnica-Krezel, L.; Korzh, V.; Halpern, M.E.; Wright, C.V. Induction of the Zebrafish Ventral Brain and Floorplate Requires Cyclops/Nodal Signalling. Nature 1998, 395, 185–189. [Google Scholar] [CrossRef]
- Tian, J.; Yam, C.; Balasundaram, G.; Wang, H.; Gore, A.; Sampath, K. A Temperature-Sensitive Mutation in the Nodal-Related Gene Cyclops Reveals That the Floor Plate Is Induced during Gastrulation in Zebrafish. Development 2003, 130, 3331–3342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Epstein, D.J. Distinct Regulators of Shh Transcription in the Floor Plate and Notochord Indicate Separate Origins for These Tissues in the Mouse Node. Development 2003, 130, 3891–3902. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, Y.; Tamplin, O.J.; Beckers, A.; Gossler, A.; Rossant, J. Live Imaging and Genetic Analysis of Mouse Notochord Formation Reveals Regional Morphogenetic Mechanisms. Dev. Cell 2007, 13, 884–896. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.D.; Dale, J.K. Notch Signalling Regulates the Contribution of Progenitor Cells from the Chick Hensen’s Node to the Floor Plate and Notochord. Development 2010, 137, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, M.E.; Hatta, K.; Amacher, S.L.; Talbot, W.S.; Yan, Y.L.; Thisse, B.; Thisse, C.; Postlethwait, J.H.; Kimmel, C.B. Genetic Interactions in Zebrafish Midline Development. Dev. Biol. 1997, 187, 154–170. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.; Rembold, M.; Wittbrodt, J.; Schartl, M.; Winkler, C. Medial Floor Plate Formation in Zebrafish Consists of Two Phases and Requires Trunk-Derived Midkine-a. Genes Dev. 2005, 19, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Odenthal, J.; Haffter, P.; Vogelsang, E.; Brand, M.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; et al. Mutations Affecting the Formation of the Notochord in the Zebrafish, Danio Rerio. Development 1996, 123, 103–115. [Google Scholar]
- Strähle, U.; Blader, P.; Ingham, P.W. Expression of Axial and Sonic Hedgehog in Wildtype and Midline Defective Zebrafish Embryos. Int. J. Dev. Biol. 1996, 40, 929–940. [Google Scholar]
- Epstein, D.J.; McMahon, A.P.; Joyner, A.L. Regionalization of Sonic Hedgehog Transcription along the Anteroposterior Axis of the Mouse Central Nervous System Is Regulated by Hnf3-Dependent and -Independent Mechanisms. Development 1999, 126, 281–292. [Google Scholar] [PubMed]
- Patten, I.; Kulesa, P.; Shen, M.M.; Fraser, S.; Placzek, M. Distinct Modes of Floor Plate Induction in the Chick Embryo. Development 2003, 130, 4809–4821. [Google Scholar] [CrossRef] [Green Version]
- Le Douarin, N.M.; Halpern, M.E. Discussion Point. Origin and Specification of the Neural Tube Floor Plate: Insights from the Chick and Zebrafish. Curr. Opin. Neurobiol. 2000, 10, 23–30. [Google Scholar] [CrossRef]
- Placzek, M.; Dodd, J.; Jessell, T.M. Discussion Point. The Case for Floor Plate Induction by the Notochord. Curr. Opin. Neurobiol. 2000, 10, 15–22. [Google Scholar] [CrossRef]
- Hudson, C.; Kawai, N.; Negishi, T.; Yasuo, H. β-Catenin-Driven Binary Fate Specification Segregates Germ Layers in Ascidian Embryos. Curr. Biol. 2013, 23, 491–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobral, D.; Tassy, O.; Lemaire, P. Highly Divergent Gene Expression Programs Can Lead to Similar Chordate Larval Body Plans. Curr. Biol. 2009, 19, 2014–2019. [Google Scholar] [CrossRef]
- Takatori, N.; Kumano, G.; Saiga, H.; Nishida, H. Segregation of Germ Layer Fates by Nuclear Migration-Dependent Localization of Not MRNA. Dev. Cell 2010, 19, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothbacher, U.; Bertrand, V.; Lamy, C.; Lemaire, P. A Combinatorial Code of Maternal GATA, Ets and -Catenin-TCF Transcription Factors Specifies and Patterns the Early Ascidian Ectoderm. Development 2007, 134, 4023–4032. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.; Takada, N.; Satoh, N.; Satou, Y. (β)-Catenin Mediates the Specification of Endoderm Cells in Ascidian Embryos. Development 2000, 127, 3009–3020. [Google Scholar] [PubMed]
- Imai, K.S.; Hudson, C.; Oda-Ishii, I.; Yasuo, H.; Satou, Y. Antagonism between β-Catenin and Gata.a Sequentially Segregates the Germ Layers of Ascidian Embryos. Development 2016, 143, 4167–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.J.; Kumano, G.; Nishida, H. Cell Fate Polarization in Ascidian Mesenchyme/Muscle Precursors by Directed FGF Signaling and Role for an Additional Ectodermal FGF Antagonizing Signal in Notochord/Nerve Cord Precursors. Development 2007, 134, 1509–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Satoh, N.; Satou, Y. Early Embryonic Expression of FGF4/6/9 Gene and Its Role in the Induction of Mesenchyme and Notochord in Ciona Savignyi Embryos. Development 2002, 129, 1729–1738. [Google Scholar] [PubMed]
- Kobayashi, K.; Sawada, K.; Yamamoto, H.; Wada, S.; Saiga, H.; Nishida, H. Maternal Macho-1 Is an Intrinsic Factor That Makes Cell Response to the Same FGF Signal Differ between Mesenchyme and Notochord Induction in Ascidian Embryos. Development 2003, 130, 5179–5190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, C.; Yasuo, H. Patterning across the Ascidian Neural Plate by Lateral Nodal Signalling Sources. Development 2005, 132, 1199–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mita, K.; Fujiwara, S. Nodal Regulates Neural Tube Formation in the Ciona Intestinalis Embryo. Dev. Genes Evol. 2007, 217, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.S.; Levine, M.; Satoh, N.; Satou, Y. Regulatory Blueprint for a Chordate Embryo. Science 2006, 312, 1183–1187. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating Cellular Diversity and Spatial Form: Wnt Signaling and the Evolution of Multicellular Animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.Q.; Bowerman, B. Animal Development: An Ancient β-Catenin Switch? Curr. Biol. 2013, 23, R313–315. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, J.; Ericson, J. The Specification of Neuronal Identity by Graded Sonic Hedgehog Signalling. Semin. Cell Dev. Biol. 1999, 10, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C.; Ba, M.; Rouvière, C.; Yasuo, H. Divergent Mechanisms Specify Chordate Motoneurons: Evidence from Ascidians. Development 2011, 138, 1643–1652. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.F.M.T.; Moly, P.K.; Miyamoto, Y.; Kusakabe, T.G. Distinctive Expression Patterns of Hedgehog Pathway Genes in the Ciona Intestinalis Larva: Implications for a Role of Hedgehog Signaling in Postembryonic Development and Chordate Evolution. Zoolog. Sci. 2010, 27, 84–90. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Okada, T.; Matsumoto, J.; Ohtsuka, Y.; Terashima, T.; Okamura, Y. Early Specification of Ascidian Larval Motor Neurons. Dev. Biol. 2005, 278, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Takatori, N.; Satou, Y.; Satoh, N. Expression of Hedgehog Genes in Ciona Intestinalis Embryos. Mech. Dev. 2002, 116, 235–238. [Google Scholar] [CrossRef]
- Hudson, C.; Sirour, C.; Yasuo, H. Snail Mediates Medial-Lateral Patterning of the Ascidian Neural Plate. Dev. Biol. 2015, 403, 172–179. [Google Scholar] [CrossRef]
- Hudson, C.; Yasuo, H. A Signalling Relay Involving Nodal and Delta Ligands Acts during Secondary Notochord Induction in Ciona Embryos. Development 2006, 133, 2855–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Hino, K.; Yagi, K.; Satoh, N.; Satou, Y. Gene Expression Profiles of Transcription Factors and Signaling Molecules in the Ascidian Embryo: Towards a Comprehensive Understanding of Gene Networks. Development 2004, 131, 4047–4058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, C.; Sirour, C.; Yasuo, H. Co-Expression of Foxa.a, Foxd and Fgf9/16/20 Defines a Transient Mesendoderm Regulatory State in Ascidian Embryos. Elife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.-L.; Akiyama, R.; Bessho, Y.; Matsui, T. ERK Activity Dynamics during Zebrafish Embryonic Development. Int. J. Mol. Sci. 2018, 20, 109. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.L.; Kimelman, D. Wnt Signaling and the Evolution of Embryonic Posterior Development. Curr. Biol. 2009, 19, R215–219. [Google Scholar] [CrossRef] [Green Version]
- Pasini, A.; Manenti, R.; Rothbächer, U.; Lemaire, P. Antagonizing Retinoic Acid and FGF/MAPK Pathways Control Posterior Body Patterning in the Invertebrate Chordate Ciona Intestinalis. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, S.; Roure, A.; Piron, J.; Darras, S. Antero-Posterior Ectoderm Patterning by Canonical Wnt Signaling during Ascidian Development. PLoS Genet. 2019, 15, e1008054. [Google Scholar] [CrossRef] [Green Version]
- Kanda, M.; Ikeda, T.; Fujiwara, S. Identification of a Retinoic Acid-Responsive Neural Enhancer in the Ciona Intestinalis Hox1 Gene. Dev. Growth Differ. 2013, 55, 260–269. [Google Scholar] [CrossRef]
- Sasakura, Y.; Kanda, M.; Ikeda, T.; Horie, T.; Kawai, N.; Ogura, Y.; Yoshida, R.; Hozumi, A.; Satoh, N.; Fujiwara, S. Retinoic Acid-Driven Hox1 Is Required in the Epidermis for Forming the Otic/Atrial Placodes during Ascidian Metamorphosis. Development 2012, 139, 2156–2160. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Satoh, N.; Saiga, H. Limited Functions of HOX Genes in the Larval Development of the Ascidian Ciona Intestinalis. Development 2010, 137, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Polevoy, H.; Gutkovich, Y.E.; Michaelov, A.; Volovik, Y.; Elkouby, Y.M.; Frank, D. New Roles for Wnt and BMP Signaling in Neural Anteroposterior Patterning. EMBO Rep. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, W.R.; Swalla, B.J. Evolution of Alternate Modes of Development in Ascidians. Bioessays 1992, 14, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N. Developmental Biology of Ascidians; Developmental and Cell Biology Series; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1994; ISBN 978-0-521-35221-5. [Google Scholar]
- Cavey, M.J. Myogenic Events in Compound Ascidian Larvae. Am. Zool. 1982, 22, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Corbo, J.C.; Erives, A.; Di Gregorio, A.; Chang, A.; Levine, M. Dorsoventral Patterning of the Vertebrate Neural Tube Is Conserved in a Protochordate. Development 1997, 124, 2335–2344. [Google Scholar]
- Teillet, M.A.; Lapointe, F.; Le Douarin, N.M. The Relationships between Notochord and Floor Plate in Vertebrate Development Revisited. Proc. Natl. Acad. Sci. USA 1998, 95, 11733–11738. [Google Scholar] [CrossRef] [Green Version]
- Appel, B.; Fritz, A.; Westerfield, M.; Grunwald, D.J.; Eisen, J.S.; Riley, B.B. Delta-Mediated Specification of Midline Cell Fates in Zebrafish Embryos. Curr. Biol. 1999, 9, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Briggs, J.A.; Li, V.C.; Lee, S.; Woolf, C.J.; Klein, A.; Kirschner, M.W. Mouse Embryonic Stem Cells Can Differentiate via Multiple Paths to the Same State. Elife 2017, 6, e26945. [Google Scholar] [CrossRef]
- Arendt, D.; Musser, J.M.; Baker, C.V.H.; Bergman, A.; Cepko, C.; Erwin, D.H.; Pavlicev, M.; Schlosser, G.; Widder, S.; Laubichler, M.D.; et al. The Origin and Evolution of Cell Types. Nat. Rev. Genet. 2016, 17, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Kawashima, T.; Shoguchi, E.; Nakayama, A.; Satoh, N. An Integrated Database of the Ascidian, Ciona Intestinalis: Towards Functional Genomics. Zool. Sci. 2005, 22, 837–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozovic, M.; Dantec, C.; Dardaillon, J.; Dauga, D.; Faure, E.; Gineste, M.; Louis, A.; Naville, M.; Nitta, K.R.; Piette, J.; et al. ANISEED 2017: Extending the Integrated Ascidian Database to the Exploration and Evolutionary Comparison of Genome-Scale Datasets. Nucleic Acids Res. 2018, 46, D718–D725. [Google Scholar] [CrossRef] [PubMed]
- Takatori, N.; Hotta, K.; Mochizuki, Y.; Satoh, G.; Mitani, Y.; Satoh, N.; Satou, Y.; Takahashi, H. T-Box Genes in the Ascidian Ciona Intestinalis: Characterization of CDNAs and Spatial Expression. Dev. Dyn. 2004, 230, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Imai, K.S. Ascidian Zic Genes. Adv. Exp. Med. Biol. 2018, 1046, 87–106. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hudson, C.; Yasuo, H. Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates. Genes 2021, 12, 592. https://doi.org/10.3390/genes12040592
Hudson C, Yasuo H. Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates. Genes. 2021; 12(4):592. https://doi.org/10.3390/genes12040592
Chicago/Turabian StyleHudson, Clare, and Hitoyoshi Yasuo. 2021. "Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates" Genes 12, no. 4: 592. https://doi.org/10.3390/genes12040592
APA StyleHudson, C., & Yasuo, H. (2021). Neuromesodermal Lineage Contribution to CNS Development in Invertebrate and Vertebrate Chordates. Genes, 12(4), 592. https://doi.org/10.3390/genes12040592