
Structures of the ß-Keratin Filaments and Keratin Intermediate Filaments in the Epidermal Appendages of Birds and Reptiles (Sauropsids)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
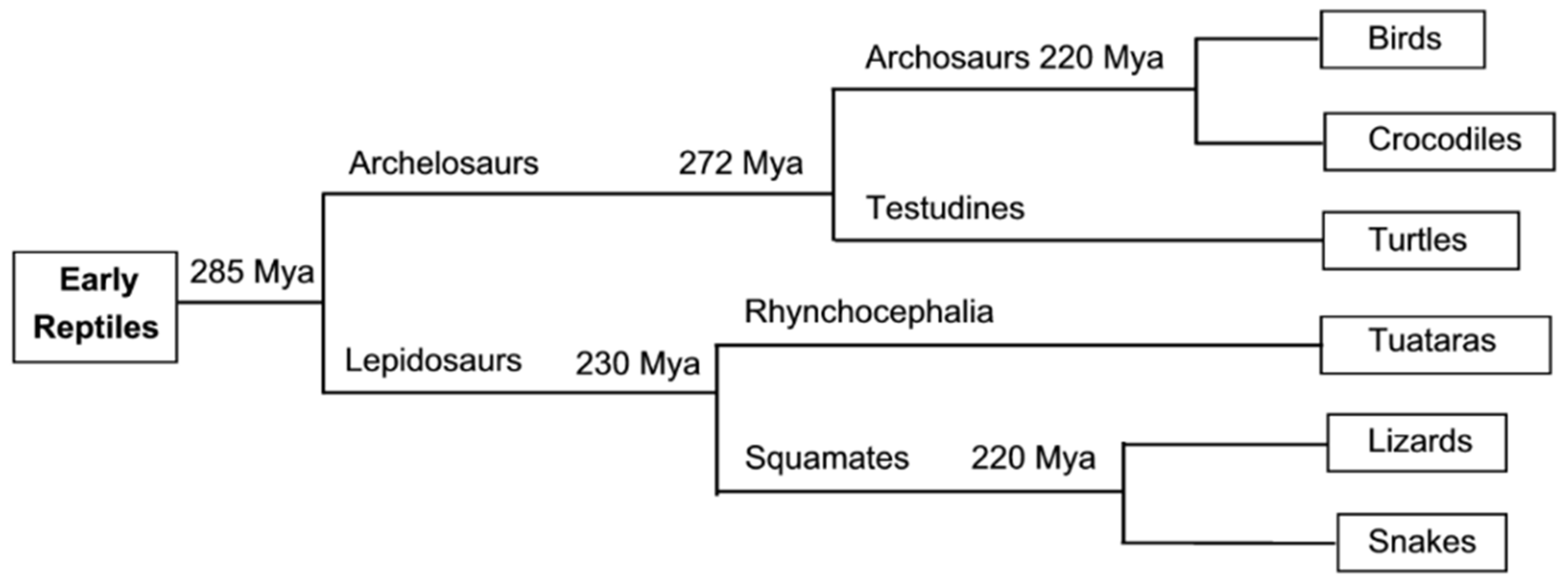
:1. Introduction
2. The 3.4 nm Diameter Filaments Formed from the Corneous ß-Proteins
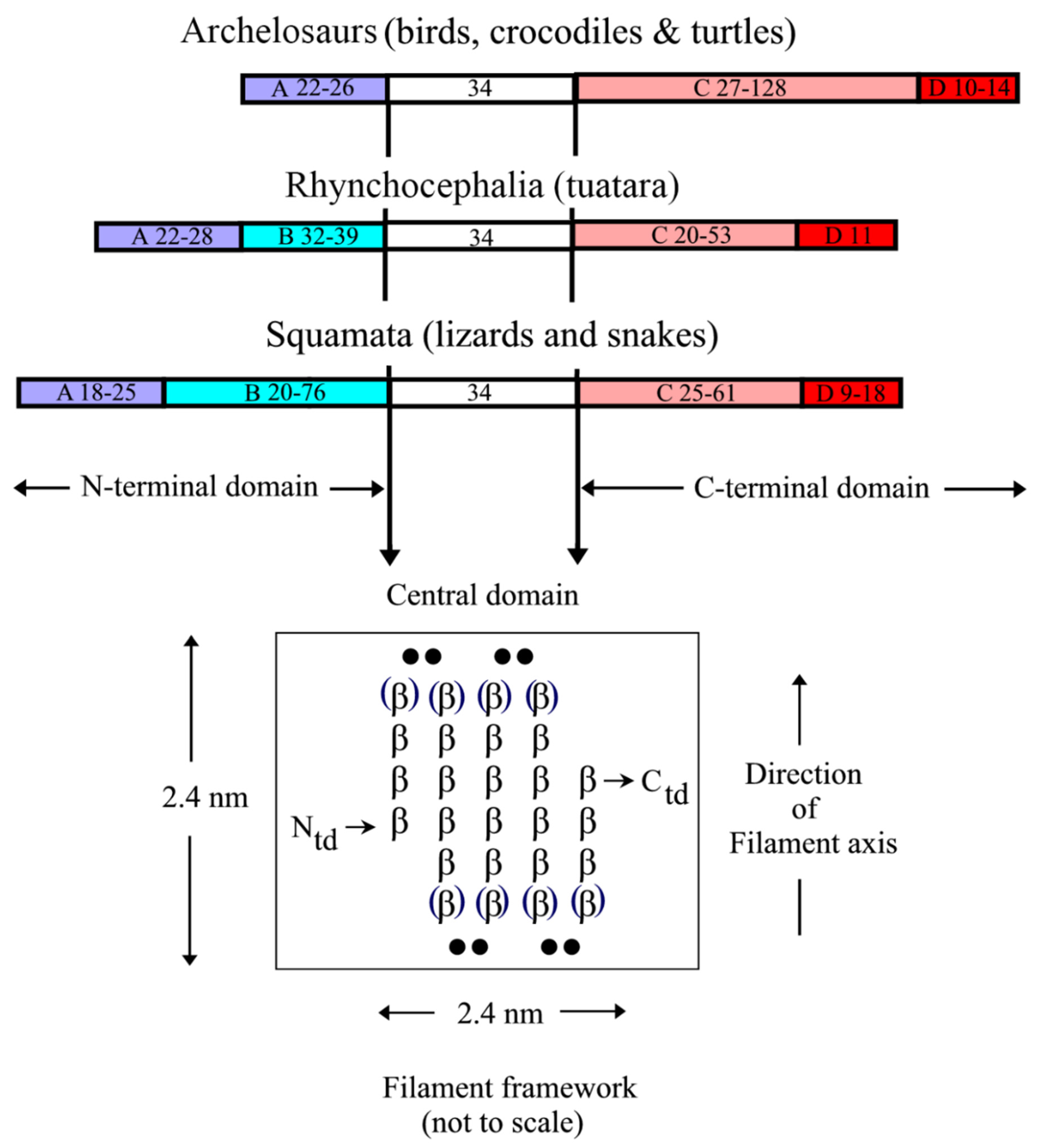
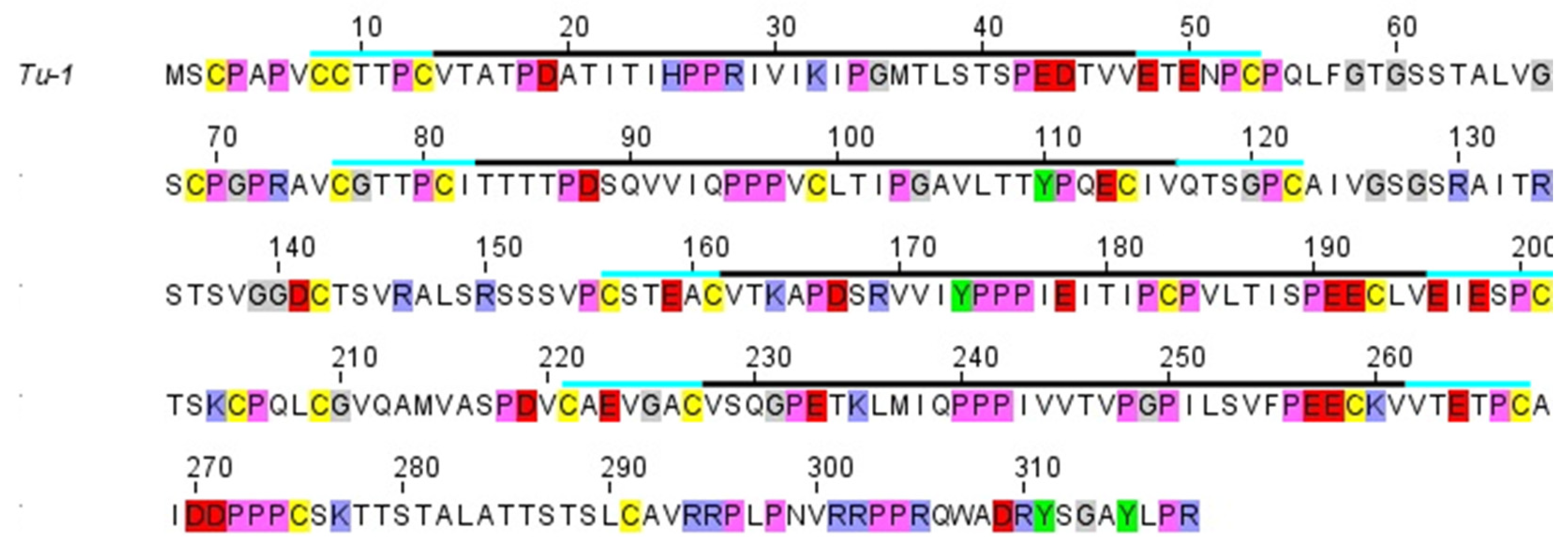
2.1. Structure of the Corneous ß-Protein Molecules and Their Assembly into Filaments
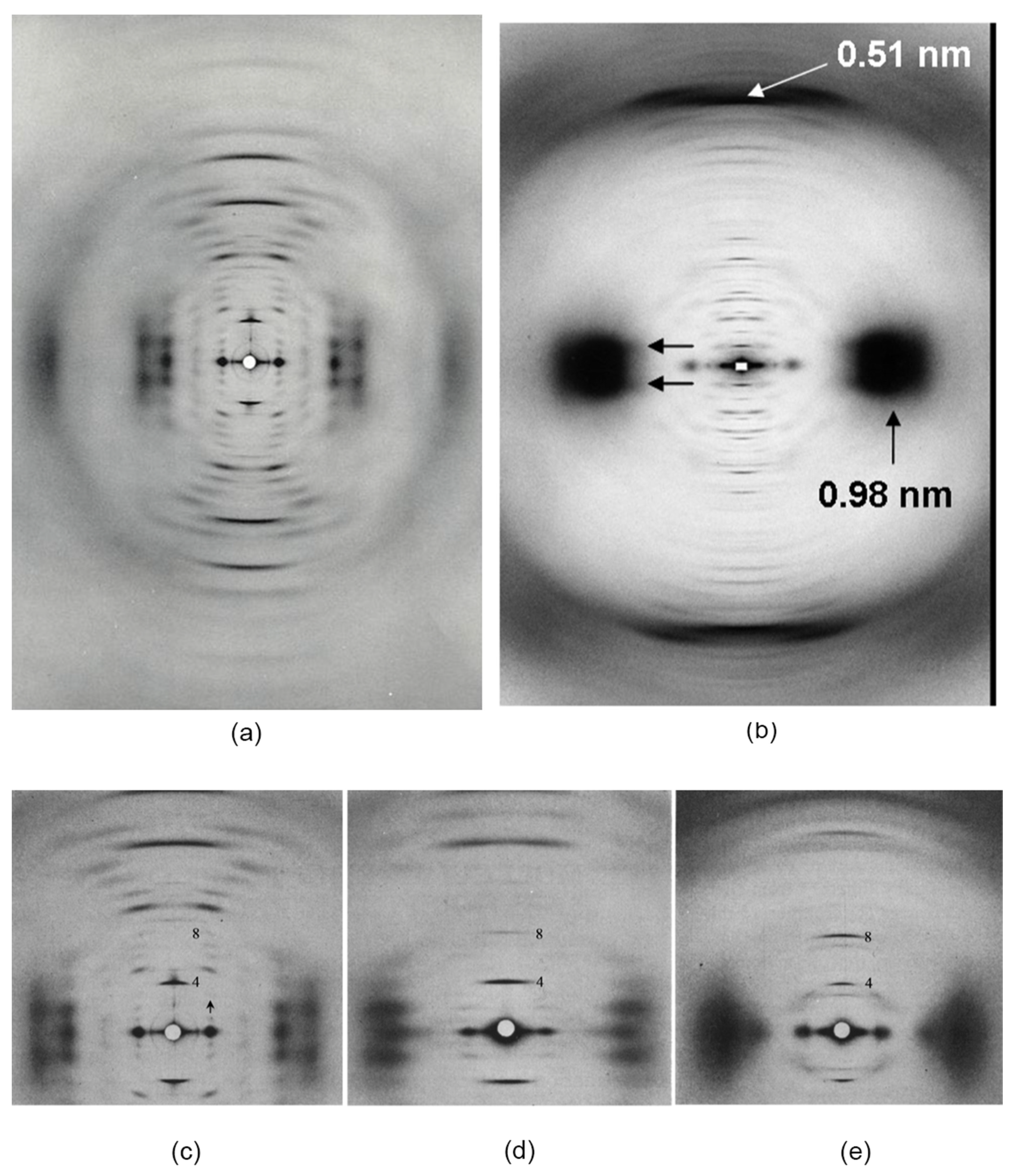
2.2. Lateral Arrangement of the 3.4 nm Diameter Filaments
2.3. Physical Properties of the 3.4 nm Diameter Filaments in Terms of Sequence Features
3. Structure of the Keratin Intermediate Filament (KIF) Molecules and the 7–10 nm Diameter Filaments
3.1. Sequence of KIF Chains in Sauropsids
3.2. Molecular Structure and Arrangement in the KIF
3.3. Sauropsids Do Not Contain the Families of Matrix Proteins Seen Around Mammalian Trichocyte KIF
4. Potential Interactions between Proteins Constituting the Epidermal Appendages
5. Summary
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Rest, J.S.; Ast, J.C.; Austin, C.C.; Wadell, P.J.; Tibbetts, E.A.; Hay, J.M.; Mindell, D.P. Molecular systematics of primary reptilian lineages and the tuatara mitochondrial genome. Mol. Phylogenet. Evol. 2003, 29, 289–297. [Google Scholar] [CrossRef]
- Rice, R.H.; Winters, B.R.; Durbin-Johnson, B.P.; Rocke, D.M. Chicken corneocyte cross-linked proteome. J. Proteome Res. 2013, 12, 771–776. [Google Scholar] [CrossRef]
- Strasser, B.; Mlitz, V.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich proteins in feathers and hair. BMC Evol. Biol. 2015, 15, 82. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Ng, C.S.; Yan, J.; Lai, Y.C.; Chen, C.K.; Lai, Y.T.; Wu, S.M.; Chen, J.J.; Luo, W.; Widelitz, R.B.; et al. Topographical mapping of α- and β-keratins on developing chicken skin integument: Functional interaction and evolutionary perspectives. Proc. Natl. Acad. Sci. USA 2015, 122, E6770–E6779. [Google Scholar] [CrossRef] [Green Version]
- Parry, D.A.D.; Fraser, R.D.B.; Alibardi, L.; Rutherford, K.; Gemmell, N. Molecular structure of sauropsid β-keratins from tuatara (Sphenodon punctatus). J. Struct. Biol. 2019, 207, 21–28. [Google Scholar] [CrossRef]
- Filshie, B.K.; Rogers, G.E. An electron microscope study of the fine structure of feather keratin. J. Cell Biol. 1962, 13, 1–12. [Google Scholar] [CrossRef]
- Alexander, N.J. Comparison of alpha and beta keratins in reptiles. Z. Zellforsch. Mikrosk. Anat. 1970, 110, 153–165. [Google Scholar]
- Maderson, P.F.A.; Flaxman, B.A.; Roth, S.I.; Szabo, G. Ultrastructural contribution to the identification of cell types in the lizard epidermal generation. J. Morphol. 1972, 136, 191–209. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P. Conformation in Fibrous Proteins and Related Synthetic Polypeptides; Academic Press: London, UK, 1973. [Google Scholar]
- Landmann, L. Keratin formation and barrier mechanisms in the epidermis of Natrix natrix (Reptilia, serpentes): An ultrastructural study. J. Morphol. 1979, 162, 93–126. [Google Scholar] [CrossRef]
- Alibardi, L.; Dalla Valle, L.; Toffolo, V.; Toni, M. Scale keratin in lizard epidermis reveals amino acid regions homologous with avian and mammalian proteins. Anat. Rec. 2006, 288A, 734–752. [Google Scholar] [CrossRef]
- Alibardi, L.; Dalla Valle, L.; Toni, M. Cell biology of adhesive setae in gecko lizards. Zoology 2009, 112, 403–424. [Google Scholar] [CrossRef]
- Strasser, B.; Mlitz, V.; Hermann, M.; Rice, R.H.; Eigenheer, R.A.; Alibardi, L.; Tschachler, E.; Eckhart, L. Evolutionary origin and diversification of epidermal barrier proteins in amniotes. Mol. Biol. Evol. 2014, 31, 3194–3205. [Google Scholar] [CrossRef] [Green Version]
- Purslow, P.P.; Vincent, J.F.V. Mechanical properties of primary feathers from the pigeon. J. Exp. Biol. 1978, 72, 251–260. [Google Scholar]
- Harris, B. The mechanical behaviour of composite materials. In The Mechanical Properties of Biological Materials; S.E.B. Symp. No. 34; Vincent, J.F.V., Currey, J.D., Eds.; The Society for Experimental Biology: London, UK, 1980; pp. 37–74. [Google Scholar]
- Fraser, R.D.B.; MacRae, T.P. Molecular structure and mechanical properties of keratin. In The Mechanical Properties of Biological Materials; S.E.B. Symp. No. 34; Vincent, J.F.V., Currey, J.D., Eds.; The Society for Experimental Biology: London, UK, 1980; pp. 211–246. [Google Scholar]
- Bonser, R.H.C.; Purslow, P.P. The Young’s modulus of feather keratin. J. Exp. Biol. 1995, 198, 1029–1033. [Google Scholar]
- Bonser, R.H.C. The mechanical properties of feather keratin. J. Zool. 1996, 239, 477–484. [Google Scholar] [CrossRef]
- Corning, W.R.; Biewener, A.A. In vivo strains in pigeon flight feather shafts: Implications for structural design. J. Exp. Biol. 1998, 201, 3057–3065. [Google Scholar]
- Cameron, G.J.; Wess, T.J.; Bonser, R.H.C. Young’s modulus varies with differential orientation of keratin in feathers. J. Struct. Biol. 2003, 143, 118–123. [Google Scholar] [CrossRef]
- Taylor, A.M.; Bonser, R.H.C.; Farrent, J.W. The influence of hydration on the tensile and compressive properties of avian keratinous tissues. J. Mater. Sci. 2004, 39, 939–942. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Amino acid sequence homologies in the hard keratins of birds and reptiles, and their implications for molecular structure and physical properties. J. Struct. Biol. 2014, 188, 213–224. [Google Scholar] [CrossRef]
- Alibardi, A.; Toni, M. Distribution and characterization of keratins in the epidermis of the tuatara (Sphenodon punctatus). Zool. Sci. 2006, 23, 801–807. [Google Scholar] [CrossRef]
- Rudall, K.M. X-ray studies of the distribution of protein chain types in the vertebrate epidermis. Biochim. Biophys. Acta 1947, 1, 549–562. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Toffolo, V.; Belvedere, P.; Alibardi, L. Isolation of a mRNA encoding a glycine-proline-rich β-keratin expressed in the regenerating epidermis of lizard. Dev. Dyn. 2005, 234, 934–947. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Nardi, A.; Belvedere, P.; Toni, M.; Alibardi, L. β-Keratins of differentiating epidermis of snake comprise glycine-proline-serine-rich proteins with an avian-like gene organization. Dev. Dyn. 2007, 236, 1939–1953. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Nardi, A.; Toffolo, V.; Niero, C.; Toni, M.; Alibardi, L. Cloning and characterization of scale β-keratins in the differentiating epidermis of geckoes show they are glycine-proline-serine-rich proteins with a central motif homologous to avian keratins. Dev. Dyn. 2007, 236, 374–388. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Nardi, A.; Gelmi, C.; Toni, M.; Emera, D.; Alibardi, L. β-keratins of the crocodilian epidermis: Composition, structure, and phylogenetic relationships. J. Exp. Zool. Part B Mol. Dev. Evol. 2009, 312B, 42–57. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Nardi, A.; Toni, M.; Alibardi, L. β-keratins of turtle shell comprise small glycine-proline-tyrosine rich proteins similar to those of crocodilians and birds. J. Anat. 2009, 214, 284–300. [Google Scholar] [CrossRef]
- Hallahan, D.L.; Keiper-Hrynko, N.M.; Shang, T.Q.; Ganzke, T.S.; Toni, M.; Dalla Valle, L.; Alibardi, L. Analysis of gene expression in gecko digital adhesive pads indicates significant production of cysteine- and glycine-rich beta keratins. J. Exp. Zool. 2009, 312B, 58–73. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. The structural basis of the filament-matrix texture in the avian/reptilian group of hard β-keratins. J. Struct. Biol. 2011, 173, 391–405. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Eckhart, L.; Dalla Valle, L.; Alibardi, L. Review: Evolution and diversification of corneous beta-proteins, the characteristic epidermal proteins of reptiles and birds. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 230, 438–453. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Alibardi, L.; Tschachler, E.; Eckhart, L. Identification of epidermal differentiation genes of the tuatara provides insights into the early evolution of lepidosaurian skin. Sci. Rep. 2020, 20, 12844. [Google Scholar] [CrossRef] [PubMed]
- Greenwold, M.J.; Sawyer, R.H. Genomic organization and molecular phylogenies of the beta (β) keratin multigene family in the chicken (Gallus gallus) and zebra finch (Taeniopygia guttata): Implications for feather evolution. BMC Evol. Biol. 2010, 10, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Greenwold, M.J.; Sawyer, R.H. Molecular evolution and expression of archosaurian β-keratins and the origin of feather β-keratins. J. Exp. Zool. Part B Mol. Dev. Evol. 2013, 320B, 393–405. [Google Scholar] [CrossRef]
- Parry, D.A.D.; Winter, D.J. Keratin intermediate filament chains in tuatara (Sphenodon punctatus): A comparison of tuatara and human sequences. J. Struct. Biol. 2021, 213, 107706. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Nardi, G.B.; Bonazza, G.; Zuccal, C.; Emera, D.; Alibardi, L. Forty keratin-associated β-proteins (β-proteins) form the hard layers of scales, claws, adhesive pads in the green anole lizard, Anolis carolinensis. J. Exp. Zool. Part B Mol. Dev. Evol. 2010, 314B, 11–32. [Google Scholar] [CrossRef]
- Eckhart, L.; Dalla Valle, L.; Jaeger, K.; Ballaun, C.; Szabo, S.; Nardi, A.; Buchberger, M.; Hermann, M.; Alibardi, L.; Tschachler, E. Identification of reptilian genes encoding hair keratin-like proteins suggests a new scenario for the evolutionary origin of hair. Proc. Natl. Acad. Sci. USA 2008, 105, 18419–18423. [Google Scholar] [CrossRef] [Green Version]
- Alföldi, J.; Di Palma, F.; Grabherr, M.; Williams, C.; Kong, L.; Mauceli, E.; Russell, P.; Lowe, C.B.; Glor, R.E.; Jaffe, J.D.; et al. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandebergh, W.; Bossuyt, F. Radiation and functional diversification of alpha keratins during early vertebrate evolution. Mol. Biol. Evol. 2012, 29, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M. The structure of chicken scale keratin. J. Ultrastruct. Res. 1977, 60, 27–33. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P.; Rogers, G.E. Keratins—Their Composition, Structure and Biosynthesis; Thomas: Springfield, IL, USA, 1972. [Google Scholar]
- Parry, D.A.D.; Fraser, R.D.B.; Squire, J.M. Fifty years of coiled-coils and α-helical bundles: A close relationship between sequence and structure. J. Struct. Biol. 2008, 163, 258–269. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. The structural basis of two-dimensional net pattern observed in the X-ray diffraction pattern of avian keratin. J. Struct. Biol. 2011, 176, 340–349. [Google Scholar] [CrossRef]
- Astbury, W.T.; Marwick, T.C. X-ray interpretation of the molecular structure of feather keratin. Nature 1932, 130, 309–310. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P. Molecular organization in feather keratin. J. Mol. Biol. 1959, 1, 387–397. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P. Structural organization in feather keratin. J. Mol. Biol. 1963, 7, 272–280. [Google Scholar] [CrossRef]
- Schorr, R.; Krimm, S. Studies on the structure of feather keratin I. X-ray diffraction studies and other experimental data. Biophys. J. 1961, 1, 467–487. [Google Scholar]
- O’Donnell, I.J. The complete amino acid sequence of a feather keratin from emu (Dromaius novae-hollandiae). Aust. J. Biol. Sci. 1973, 26, 415–437. [Google Scholar] [CrossRef] [Green Version]
- Fraser, R.D.B.; Suzuki, E. Polypeptide chain conformation in feather keratin. J. Mol. Biol. 1965, 14, 279–282. [Google Scholar] [CrossRef]
- Suzuki, E. Localization of beta-conformation in feather keratin. Aust. J. Biol. Sci. 1973, 26, 435–437. [Google Scholar]
- Fraser, R.D.B.; MacRae, T.P. The molecular structure of feather keratin. In Proceedings of the 16th International Ornithological Congress, Canberra, Australia, 12–17 August 1976; pp. 443–451. [Google Scholar]
- Sawyer, R.H.; Glenn, T.; French, J.O.; Mays, B.; Shames, R.B.; Barnes, G.L.; Rhodes, W.; Ishikawa, Y. The expression of beta (β) keratins in the epidermal appendages of reptiles and birds. Am. Zool. 2000, 40, 530–539. [Google Scholar] [CrossRef] [Green Version]
- Alibardi, L.; Toni, M. β-keratins of reptilian scales share a central amino acid sequence termed core-box. Res. J. Biol. Sci. 2007, 2, 329–339. [Google Scholar]
- Inglis, A.S.; Gillespie, M.J.; Roxborough, C.M.; Whitaker, L.A.; Casagranda, F. Sequence of a glycine-rich protein from lizard claw: Unusual dilute acid and heptafluorobutyric acid cleavages. In Proteins, Structure and Function; L’Italien, J.L., Ed.; Plenum: New York, NT, USA, 1987; pp. 757–764. [Google Scholar]
- Fraser, R.D.B.; Parry, D.A.D. The molecular structure of reptilian keratin. Int. J. Biol. Macromol. 1996, 19, 207–211. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Molecular packing in the feather keratin filament. J. Struct. Biol. 2008, 162, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; Parry, D.A.D. Filamentous structure of hard β-keratin in the epidermal appendages of birds and reptiles. In Subcellular Biochemistry: Fibrous Proteins: Structures and Mechanisms; Parry, D.A.D., Squire, J.M., Eds.; Springer: Cham, Switzerland, 2017; Volume 82, pp. 231–252. [Google Scholar]
- Chothia, C.; Finkelstein, A.V. The classification and origins of protein folding patterns. Ann. Rev. Biochem. 1990, 57, 1007–1039. [Google Scholar] [CrossRef] [PubMed]
- Kister, A.E.; Finkelstein, A.V.; Gelfand, I.M. Common features in structures and sequences of sandwich-like proteins. Proc. Natl. Acad. Sci. USA 2002, 99, 14137–14141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, R.D.B.; MacRae, T.P.; Parry, D.A.D.; Suzuki, E. The structure of feather keratin. Polymer 1971, 12, 35–56. [Google Scholar] [CrossRef]
- Calvaresi, M.; Eckhart, L.; Alibardi, L. The molecular organization of the beta-sheet region in corneous beta-proteins (beta-keratins) of sauropsids explains it stability and polymerization into filaments. J. Struct. Biol. 2016, 194, 282–291. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Lepidosaur β-keratin chains with four 34-residue repeats: Modelling reveals a potential filament-crosslinking role. J. Struct. Biol. 2020, 209, 107413. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Mlitz, V.; Strasser, B.; Tschachler, E.; Alibardi, L.; Eckhart, L. Identification and comparative analysis of the epidermal differentiation complex in snakes. Sci. Rep. 2017, 7, 45338. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, Q.; Wang, Y.; Luo, L.; Yang, J.; Yang, L.; Liu, M.; Li, Y.R.; Qian, T.M.; Zheng, Y.; et al. Gekko japonicus genome reveals evolution of adhesive toe pads and tail regeneration. Nat. Commun. 2015, 6, 10033. [Google Scholar] [CrossRef]
- Filshie, B.K.; Fraser, R.D.B.; MacRae, T.P.; Rogers, G.E. X-ray diffraction and electron microscope observations on soluble derivatives of feather keratin. Biochem. J. 1964, 92, 18–19. [Google Scholar] [CrossRef]
- Parry, D.A.D. Structure and topology of the linkers in the conserved lepidosaur β-keratin chain with four 34-residue repeats support an interfilament role for the central linker. J. Struct. Biol. 2020, 212, 107599. [Google Scholar] [CrossRef]
- Bear, R.S.; Rugo, H.J. The results of X-ray diffraction studies on keratin fibers. Ann. N. Y. Acad. Sci. 1951, 53, 627–648. [Google Scholar]
- Steinert, P.M.; Mack, J.W.; Korge, B.P.; Gan, S.-Q.; Haynes, S.R.; Steven, A.C. Glycine loops in proteins: Their occurrence in certain intermediate filament chains, loricrins and single-stranded RNA binding proteins. Int. J. Biol. Macromol. 1991, 13, 130–139. [Google Scholar]
- Filshie, B.K.; Rogers, G.E. The fine structure of α-keratin. J. Mol. Biol. 1961, 3, 784–786. [Google Scholar]
- Fraser, R.D.B.; Parry, D.A.D. Structural transition of trichocyte keratin intermediate filaments during development in the hair follicle. In Subcellular Biochemistry: Fibrous Proteins: Structures and Mechanisms; Parry, D.A.D., Squire, J.M., Eds.; Springer: Cham, Switzerland, 2017; Volume 82, pp. 131–149. [Google Scholar]
- Greenwold, M.J.; Bao, W.; Jarvis, E.D.; Hu, H.; Li, C.; Gilbert, M.T.; Zhang, G.; Sawyer, R.H. Dynamic evolution of the alpha (α) and beta (β) keratins has accompanied integument diversification and the adaption of birds into novel lifestyles. BMC Evol. Biol. 2014, 14, 249. [Google Scholar]
- Ehrlich, F.; Lachner, J.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich keratins in hard skin appendages of terrestrial vertebrates. Mol. Biol. Evol. 2020, 37, 982–993. [Google Scholar] [CrossRef]
- Crewther, W.G.; Inglis, A.S.; McKern, N.M. Amino acid sequences of alpha-helical segments from S-carboxymethylkeratin-A—complete sequence of a type II segment. Biochem. J. 1978, 73, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Gough, K.H.; Inglis, A.S.; Crewther, W.G. Amino acid sequences of alpha-helical segments from S-carboxymethylkeratin-A—complete sequence of a type I segment. Biochem. J. 1978, 73, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Parry, D.A.D.; Steinert, P.M. Intermediate Filament Structure; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Strnad, P.; Usachov, V.; Debes, C.; Grater, F.; Parry, D.A.D.; Omary, M.B. Unique amino acid signatures that are evolutionarily conserved distinguish simple-type, epidermal and hair keratins. J. Cell Sci. 2011, 124, 4221–4232. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.A.; Strelkov, S.V.; Burkhard, P.; Aebi, U.; Parry, D.A.D. Sequence comparisons of intermediate filaments chains: Evidence of a unique functional/structural role for the coiled-coil segment 1A and linker L1. J. Struct. Biol. 2002, 137, 128–145. [Google Scholar] [CrossRef]
- Steinert, P.M.; Parry, D.A.D. The conserved H1 domain of the Type II keratin 1 chain plays an essential role in the alignment of nearest neighbor molecules in mouse and human keratin 1/keratin 10 intermediate filaments at the two- to four-molecule level of structure. J. Biol. Chem. 1993, 268, 2878–2887. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P.; Miller, A. The coiled-coil model of α-keratin structure. J. Mol. Biol. 1964, 10, 147–156. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P.; Miller, A. X-ray diffraction pattern of α-fibrous proteins. J. Mol. Biol. 1965, 14, 432–442. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; MacRae, T.P.; Suzuki, E. Structure of the α-keratin microfibril. J. Mol. Biol. 1976, 108, 435–452. [Google Scholar] [CrossRef]
- Guzenko, D.; Chernyatina, A.A.; Strelkov, S.V. Crystallographic studies of intermediate filaments. In Subcellular Biochemistry: Fibrous Proteins: Structures and Mechanisms; Parry, D.A.D., Squire, J.M., Eds.; Springer: Cham, Switzerland, 2017; Volume 82, pp. 151–170. [Google Scholar]
- Parry, D.A.D.; Crewther, W.G.; Fraser, R.D.B.; MacRae, T.P. Structure of α-keratin: Structural implications of the amino acid sequences of the type I and type II chain segments. J. Mol. Biol. 1977, 113, 449–454. [Google Scholar] [CrossRef]
- Crewther, W.G.; Dowling, L.M.; Steinert, P.M.; Parry, D.A.D. Structure of intermediate filaments. Int. J. Biol. Macromol. 1983, 5, 267–274. [Google Scholar] [CrossRef]
- Parry, D.A.D.; Fraser, R.D.B. Intermediate filament structure: 1. Analysis of IF protein sequence data. Int. J. Biol. Macromol. 1985, 7, 203–213. [Google Scholar] [CrossRef]
- Steinert, P.M. The two-chain coiled-coil molecule of native epidermal keratin intermediate filaments is a type I-type II heterodimer. J. Biol. Chem. 1990, 265, 8766–8774. [Google Scholar] [CrossRef]
- Hatzfeld, M.; Weber, K. The coiled-coil of in vitro assembled keratin filaments is a heterodimer of Type I and II keratin: Use of site-specific mutagenesis and recombinant protein expression. J. Cell Biol. 1990, 110, 1199–1210. [Google Scholar] [CrossRef]
- Coulombe, P.A.; Fuchs, E. Elucidating the early stages of keratin filament assembly. J. Cell Biol. 1990, 111, 153–169. [Google Scholar] [CrossRef]
- Steinert, P.M.; Marekov, L.N.; Fraser, R.D.B.; Parry, D.A.D. Keratin intermediate filament structure: Crosslinking studies yield quantitative information on molecular dimensions and mechanism of assembly. J. Mol. Biol. 1993, 230, 436–452. [Google Scholar] [CrossRef] [PubMed]
- Steinert, P.M.; Marekov, L.N.; Parry, D.A.D. Conservation of the structure of keratin intermediate filaments: Molecular mechanism by which different keratin molecules integrate into pre-existing keratin intermediate filaments during differentiation. Biochemistry 1993, 32, 10046–10056. [Google Scholar] [CrossRef]
- Wang, H.; Parry, D.A.D.; Jones, L.N.; Idler, W.W.; Marekov, L.N.; Steinert, P.M. In vitro assembly and structure of trichocyte keratin intermediate filaments: A novel role for stabilization by disulfide bonding. J. Cell Biol. 2000, 151, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; Parry, D.A.D. Structural changes in the trichocyte intermediate filaments accompanying the transition from the reduced to the oxidized form. J. Struct. Biol. 2007, 159, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; Steinert, P.M.; Parry, D.A.D. Structural changes in trichocyte keratin intermediate filaments during keratinization. J. Struct. Biol. 2003, 142, 266–271. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Keratin intermediate filaments: Differences in the sequences of the type I and type II chains explain the origin of the stability of an enzyme-resistant four-chain fragment. J. Struct. Biol. 2014, 185, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.R.; Jones, L.N.; Cheng, N.; Wall, J.S.; Parry, D.A.D.; Steven, A.C. Cryo-electron microscopy of trichocyte (hard α-keratin) intermediate filaments reveals a low-density core. J. Struct. Biol. 2002, 137, 109–118. [Google Scholar] [CrossRef]
- Parry, D.A.D.; Steinert, P.M. Intermediate filaments: Molecular architecture, assembly, dynamics and polymorphism. Quart. Rev. Biophys. 1999, 32, 99–187. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Intermediate filament structure in fully differentiated (oxidised) trichocyte keratin. J. Struct. Biol. 2017, 200, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Parry, D.A.D. Hard α-keratin intermediate filaments: An alternative interpretation of the low-angle equatorial X-ray diffraction pattern, and the axial disposition of putative disulphide bonds in the intra- and inter-protofilamentous networks. Int. J. Biol. Macromol. 1996, 19, 207–211. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. The role of disulfide bond formation in the structural transition observed in the intermediate filaments of developing hair. J. Struct. Biol. 2012, 180, 117–124. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. The three-dimensional structure of trichocyte (hard α-) keratin intermediate filaments: Features of the molecular packing deduced from the sites of induced crosslinks. J. Struct. Biol. 2005, 151, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; MacRae, T.P. The structure of the α-keratin microfibril. Biosci. Rep. 1983, 3, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; MacRae, T.P. Intermediate filament structure. Biosci. Rep. 1985, 5, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.D.B.; Parry, D.A.D. Direct evidence supporting the existence of a helical dislocation in protofilament packing in the intermediate filaments of oxidized trichocyte keratin. J. Struct. Biol. 2018, 204, 491–497. [Google Scholar] [CrossRef]
- Fraser, R.D.B.; Parry, D.A.D. Structural Hierarchy of Trichocyte Keratin Intermediate Filaments. In The Hair Fibre: Proteins, Structure and Development; Plowman, J.D., Harland, D., Deb-Choudhury, S., Eds.; Springer: Singapore, 2018; Volume 1054, pp. 57–70. [Google Scholar]
- Fraser, R.D.B.; Parry, D.A.D. Trichocyte keratin-associated proteins (KAPs). In The Hair Fibre: Proteins, Structure and Development; Plowman, J.D., Harland, D., Deb-Choudhury, S., Eds.; Springer: Singapore, 2018; Volume 1054, pp. 71–86. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parry, D.A.D. Structures of the ß-Keratin Filaments and Keratin Intermediate Filaments in the Epidermal Appendages of Birds and Reptiles (Sauropsids). Genes 2021, 12, 591. https://doi.org/10.3390/genes12040591
Parry DAD. Structures of the ß-Keratin Filaments and Keratin Intermediate Filaments in the Epidermal Appendages of Birds and Reptiles (Sauropsids). Genes. 2021; 12(4):591. https://doi.org/10.3390/genes12040591
Chicago/Turabian StyleParry, David A.D. 2021. "Structures of the ß-Keratin Filaments and Keratin Intermediate Filaments in the Epidermal Appendages of Birds and Reptiles (Sauropsids)" Genes 12, no. 4: 591. https://doi.org/10.3390/genes12040591
APA StyleParry, D. A. D. (2021). Structures of the ß-Keratin Filaments and Keratin Intermediate Filaments in the Epidermal Appendages of Birds and Reptiles (Sauropsids). Genes, 12(4), 591. https://doi.org/10.3390/genes12040591