
Evolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
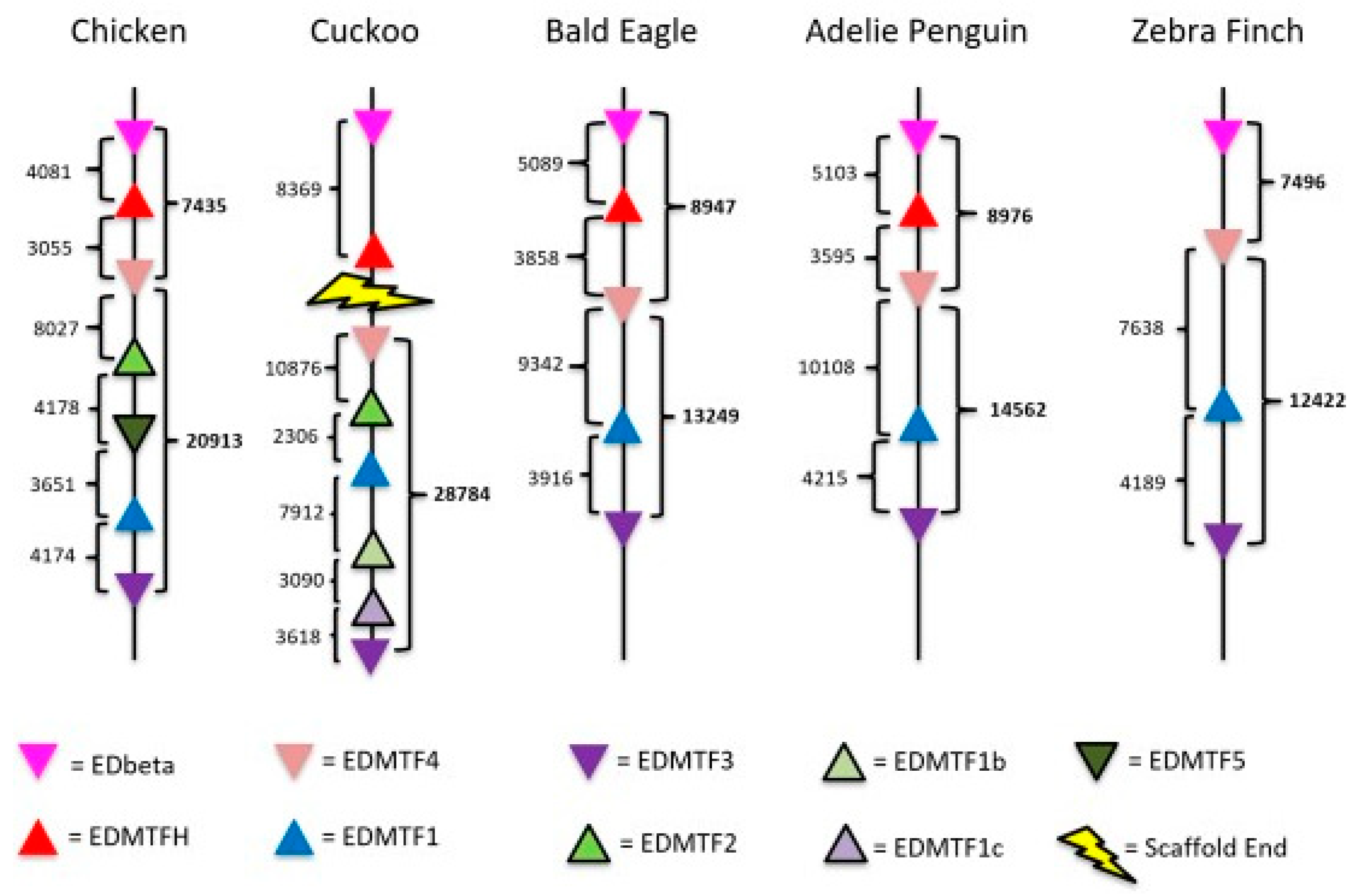
3.1. The EDAA/EDMTF Gene Family Is Conserved in the Avian EDC
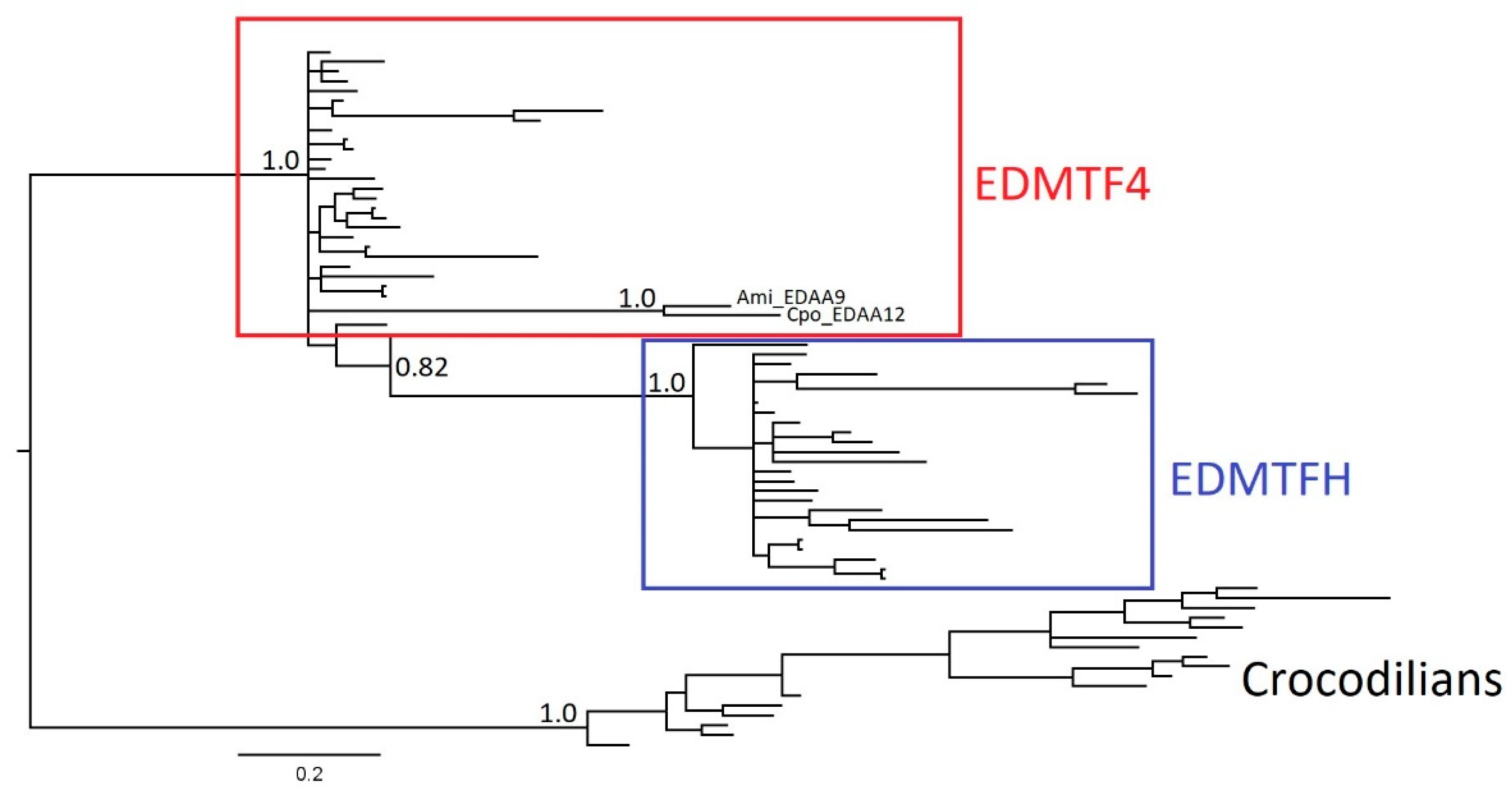
3.2. The EDAA/EDMTF Gene Family Originated in a Common Archosaur Ancestor
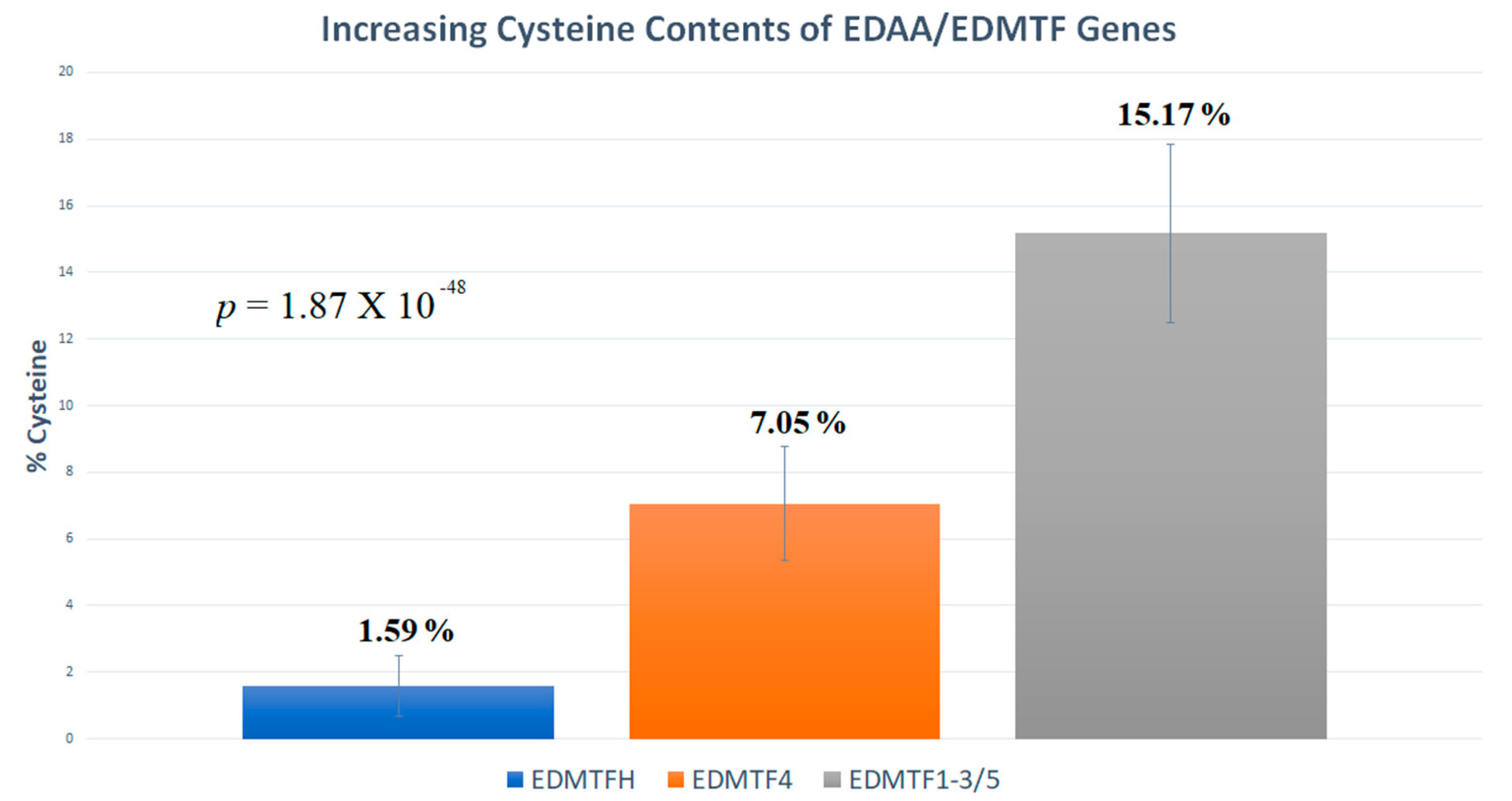
3.3. EDAA/EDMTF Genes Contain Amino Acid Contents Indicative of Epidermal Development Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sawyer, R.H.; Knapp, L.W. Avian skin development and the evolutionary origin of feathers. J. Exp. Zool. Part B Mol. Dev. Evol. 2003, 298B, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Adaptation to the land: The skin of reptiles in comparison to that of amphibians and endotherm amniotes. J. Exp. Zool. Part B Mol. Dev. Evol. 2003, 298B, 12–41. [Google Scholar] [CrossRef] [PubMed]
- Prum, R.O. Evolution of the Morphological Innovations of Feathers. J. Exp. Zool. Part B Mol. Dev. Evol. 2005, 304B, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Chuong, C.M.; Nickoloff, B.J.; Elias, P.M.; Goldsmith, L.A.; Macher, E.; Maderson, P.A.; Sundberg, J.P.; Tagami, H.; Plonka, P.M.; Thestrup-pederson, K.; et al. What is the ‘true’ function of skin? Exp. Dermatol. 2002, 11, 159–187. [Google Scholar]
- Haake, A.R.; Konig, G.; Sawyer, R.H. Avian Feather Development: Relationships between Morphogenesis and Keratinization. Dev. Biol. 1984, 106, 406–413. [Google Scholar] [CrossRef]
- Alibardi, L.; Holthaus, K.B.; Sukseree, S.; Hermann, M.; Tschachler, E.; Eckhart, L. Immunolocalization of a histidine-rich epidermal differentiation protein in the chicken supports the hypothesis of an evolutionary developmental link between the embryonic subperiderm and feather barbs and barbules. PLoS ONE 2016, 11, e0167789. [Google Scholar] [CrossRef]
- Talbot, D.; Lorgin, J.; Schorle, H. Spatiotemporal Expression Pattern of Keratins in Skin of AP-2a-deficient Mice. J. Investig. Dermatol. 1999, 113, 816–820. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, R.H.; Craig, K.F. Avian Scale Development. Absence of an “epidermal placode” in reticulate scale morphogenesis. J. Morphol. 1977, 154, 83–93. [Google Scholar] [CrossRef]
- Strasser, B.; Miltz, V.; Hermann, M.; Rice, R.G.; Eigenheer, R.A.; Alibardi, L.; Tschaler, E.; Eckhart, L. Evolutionary origin and diversification of epidermal barrier proteins in amniotes. Mol. Biol. Evol. 2014, 31, 3194–3205. [Google Scholar] [CrossRef] [Green Version]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–664. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Strasser, B.; Sipos, W.; Schmidt, H.A.; Mlitz, V.; Sukseree, S.; Weissenbacher, A.; Tschaler, E.; Alibardi, L.; Eckhart, L. Comparative genomics Identifies Epidermal Proteins Associated with the Evolution of the Turtle Shell. Mol. Biol. Evol. 2015, 33, 726–737. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Miltz, V.; Strasser, B.; Tchachler, E.; Alibardi, L.; Eckhart, L. Identification and comparative analysis of the epidermal differentiation complex in snakes. Nat. Sci. Rep. 2017, 7, 45338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holthaus, K.B.; Strasser, B.; Lacher, J.; Sukseree, S.; Sipos, W.; Weissenbacher, A.; Tshcachler, E.; ALibardi, L.; Eckhart, L. Comparative analysis of epidermal differentiation genes of crocodilians suggests new models for evolutionary origin of avian feather proteins. Genome Biol. Evol. 2018, 10, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Segre, J.A. Epidermal barrier formation and recovery in skin disorders. J. Clin. Invest. 2006, 116, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, L.; Lippens, S.; Tschachler, E.; Declercq, W. Cell Death by Cornification. Biochem. Biophys. Acta 2013, 1833, 3471–3480. [Google Scholar] [CrossRef]
- Velasco, M.V.R.; de Sa Dias, T.C.; de Freitas, A.Z.; Junior, N.D.V.; de Oliveira Pinto, C.A.S.; Kaneko, T.M.; Baby, A.R. Hair fiber characteristics and methods to evaluate hair physical and mechanical properties. Braz. J. Pharm. Sci. 2009, 45, 153–162. [Google Scholar] [CrossRef]
- Hynes, R.; Destree, A. Extensive disulfide bonding at the mammalian cell surface. Proc. Natl. Acad. Sci USA 1977, 74, 2855–2859. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.; Koblyakova, Y.; Latendorf, T.; Wu, Z.; Meyer-Hoffert, U. Cross-Linking of SPINK6 by Transflutaminases protects from epidermal proteases. J. Investig. Dermatol. 2013, 133, 1170–1177. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Takahisa, T.; Kadono, N.; Maekubo, K.; Hirai, Y. Krtap11-1, a hair keratin-associated protein, as a possible crucial element for the physical properties of hair shafts. J. Dermatol. Sci. 2014, 74, 39–47. [Google Scholar] [CrossRef]
- Strasser, B.; Miltz, V.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich proteins in feathers and hairs. BMC Evol. Biol. 2015, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachner, J.; Ehrlich, F.; Miltz, V.; Hermann, M.; ALibardi, L.; Tschaler, E.; Eckhart, L. Immunolocalization and phylogenetic profiling of the feather protein with the highest cysteine content. Protoplasma 2019, 256, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, A.C.; Greenwold, M.J.; Sawyer, R.H. Complex Gene Loss and Duplication Events Have Facilitated the Evolution of Multiple Loricrin Genes in Diverse Bird Species. Genome Biol. Evol. 2019, 11, 984–1001. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.; Li, J.; Kong, L.; Hu, H.; Pan, H.; Xu, L.; Deng, Y.; Li, Q.; Jin, L.; et al. Two Antarctic penguin genomes reveal insights into their evolutionary history and molecular changes related to the Antarctic environment. Gigascience 2014, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.; Mugal, C.; Nabholz, B.; Scheilzeth, H.; Wolf, J.B.W.; Backstrom, N.; Kunstner, A.; Balakrishnan, C.N.; Heher, A.; Ponting, C.P.; et al. Molecular evolution of genes in avian genomes. Genome Biol. Evol. 2010, 11, R68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwold, M.J.; Bao, W.; Jarvis, E.D.; Hu, H.; Li, C.; Gilbert, M.T.P.; Zhang, G.; Sawyer, R.H. Dynamic evolution of the alpha (α) and beta (β) keratins has accompanied integument diversification the adaptation of birds into novel lifestyles. BMC Evol Biol. 2014, 14, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presland, R.B.; Gregg, K.; Molloy, P.L.; Morris, C.P.; Crocker, L.A.; Rogers, G.E. Avian keratin genes, I. A molecular analysis of the structure and expression of a group of feather keratin genes. J. Mol. Biol 1989, 209, 549–560. [Google Scholar] [CrossRef]
- Presland, R.B.; Whitbread, L.A.; Rogers, G.E. Avian keratin genes, II. Chromosomal arrangement and close linkage of three gene families. J. Mol. Biol. 1989, 209, 561–576. [Google Scholar] [CrossRef]
- Greenwold, M.J.; Sawyer, R.H. Genomic organization and molecular phylogenies of the beta (β) keratin multigene family in the chicken (Gallus gallus) and zebra finch (Taeniopygia guttata): Implications for feather evolution. BMC Evol. Biol. 2010, 10, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Ng, C.S.; Yan, J.; Lai, Y.; Chen, C.; Lai, Y.; Wu, S.; Chen, J.; Luo, W.; Widelitz, R.B.; et al. Topographical mapping of α- and β-keratins on developing chicken skin integuments: Functional interaction and evolutionary perspectives. Proc. Natl. Acad. Sci. USA 2015, 112, E6770–E6779. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gish, W.; States, D.J. Identification of protein coding regions by database similarity search. Nat. Genet. 1993, 3, 266–272. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence lignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsonbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post- analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Phylogenetic Viewing Software. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 February 2019).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Smeds, L.; Qvarnstrom, A.; Ellegren, H. Direct estimate of the rate of germline mutation in a bird. Genome Res. 2016, 26, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Axelsson, E.; Smith, N.G.C.; Sundstrom, H.; Berlin, S.; Ellegren, H. Male-biased mutation rate and divergence in autosomal, z-linked and w-linked introns of chicken and turkey. Mol. Biol. Evol. 2004, 21, 1538–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.-H. Molecular Evolution; Sinauers Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; John, M.W., Ed.; Humana Press: Totowa, NJ, USA, 2002; pp. 571–607. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Smyth, G.K. Limma: Linear models for microarray data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- Gerbrands, J.J. On the Relationships Between SVD, KLT and PCA. Pattern Recognit. 1981, 14, 375–381. [Google Scholar] [CrossRef]
- Hron, T.; Pajer, P.; Paces, J.; Bartunek, P.; Elleder, D. Hidden genes in birds. Genome Biol. 2015, 16, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, G.L.; Sawyer, R.H. Histidine-rich protein B of embryonic feathers present in the transient embryonic layers of scutate scales. J. Exp. Zool. Part B Mol. Dev. Evol. 1995, 271, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. Species richness and morphological diversity of passerine birds. Proc. Natl. Acad. Sci. USA 2012, 109, 14482–14487. [Google Scholar] [CrossRef] [Green Version]
- Oliveros, C.H.; Field, D.J.; Ksepka, D.T.; Barker, F.K.; Aleixo, A. Earth History and the Passerine Superradiation. Proc. Natl. Acad. Sci. USA 2019, 116, 7917–7925. [Google Scholar] [CrossRef] [Green Version]
- Steinert, P.; Mack, J.; Korge, B.; Gan, S.Q.; Haynes, S.; Steven, A. Glycine Loops in Proteins: Their occurrence in certain intermediate filament chains, loricrins and single-stranded RNA binding protiens. Int. J. Biol. Macromol. 1991, 13, 130–139. [Google Scholar] [CrossRef]
- Alibardi, L.; Dalla Valle, L.; Nardi, A.; Toni, M. Evolution of hard proteins in the sauropsid integument in relation to the cornification of skin derivatives in amniotes. J. Anat. 2009, 214, 560–586. [Google Scholar] [CrossRef]
- Leyvraz, C.; Charles, R.P.; Rubera, I.; Guitard, M.; Rotman, S.; Breiden, B.; Sandhoff, K.; Hummler, E. The epidermal barrier function is dependent on the serine protease CAP1/Prss8. J. Cell Bio. 2005, 170, 487–496. [Google Scholar] [CrossRef]
- Greenwold, M.J.; Sawyer, R.H. Linking the molecular evolution of avian beta (β) keratins to the evolution of feathers. J. Exp. Zool. Part B Mol. Dev. Evol. 2011, 316, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, R.H.; Glenn, T.; French, J.O.; Mays, B.; Shames, R.B.; Barnes, G.L.; Rhodes, W.; Ishikawa, Y. The Expression of Beta (β) keratins in the epidermal appendages of reptiles and birds. Am. Zool. 2000, 40, 530–539. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, A.; Greenwold, M.J. Evolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds. Genes 2021, 12, 767. https://doi.org/10.3390/genes12050767
Davis A, Greenwold MJ. Evolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds. Genes. 2021; 12(5):767. https://doi.org/10.3390/genes12050767
Chicago/Turabian StyleDavis, Anthony, and Matthew J. Greenwold. 2021. "Evolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds" Genes 12, no. 5: 767. https://doi.org/10.3390/genes12050767
APA StyleDavis, A., & Greenwold, M. J. (2021). Evolution of an Epidermal Differentiation Complex (EDC) Gene Family in Birds. Genes, 12(5), 767. https://doi.org/10.3390/genes12050767