
The Trichohyalin-Like Protein Scaffoldin Is Expressed in the Multilayered Periderm during Development of Avian Beak and Egg Tooth

 ,
, 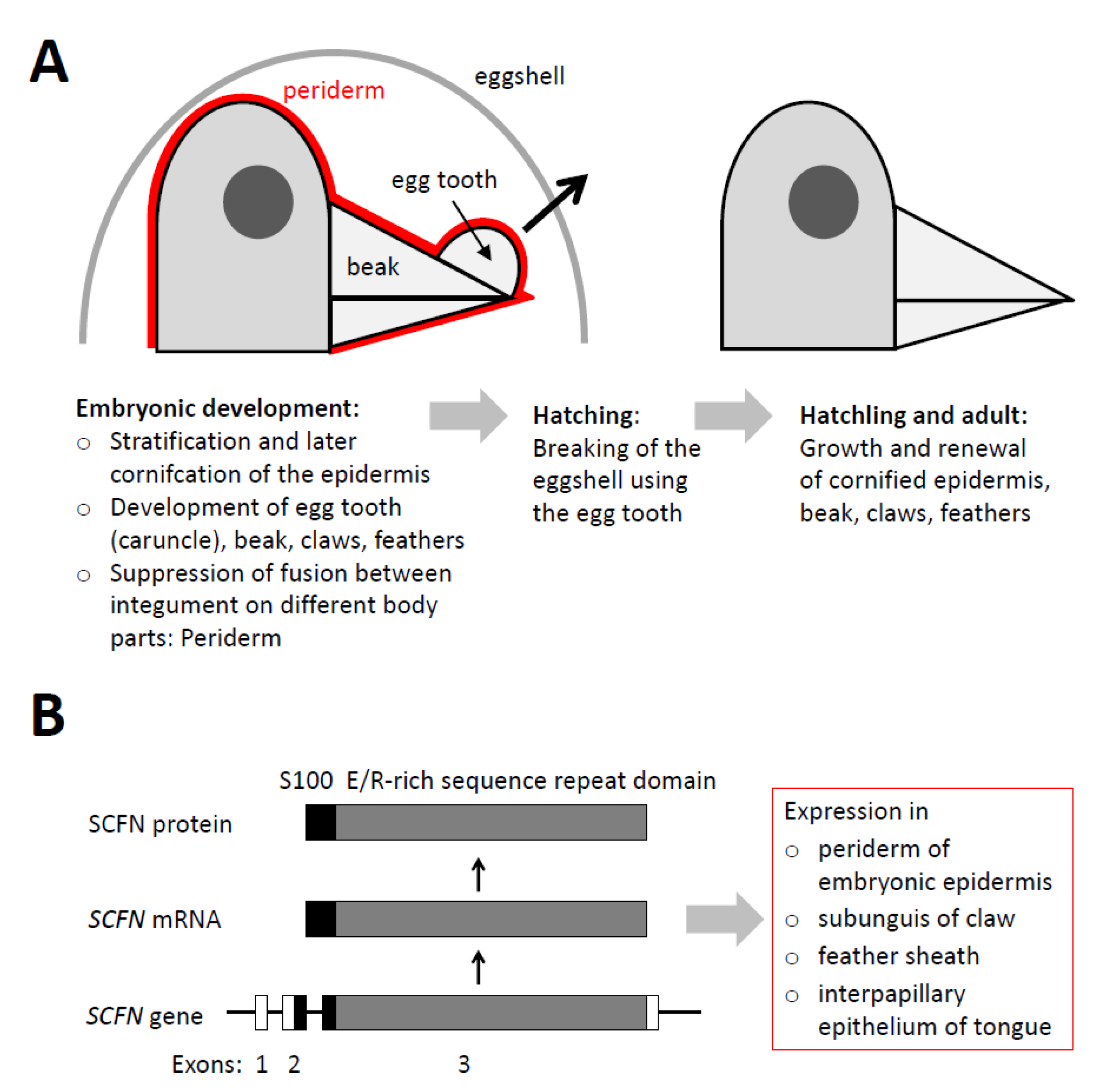{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. RNA Extraction, cDNA Synthesis and Real-Time PCR
2.3. Histological Staining and Immunohistochemistry
2.4. In Situ Hybridization
3. Results
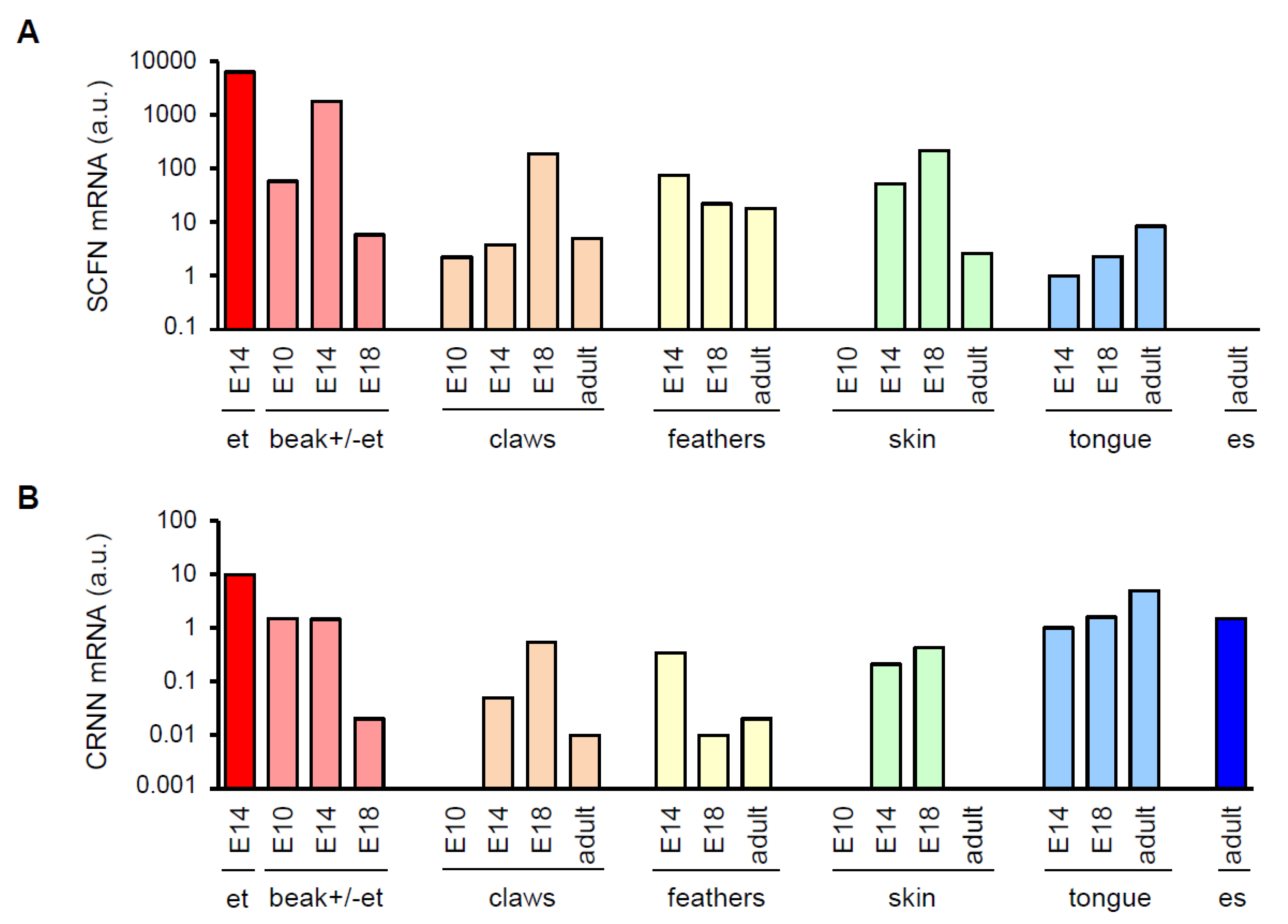
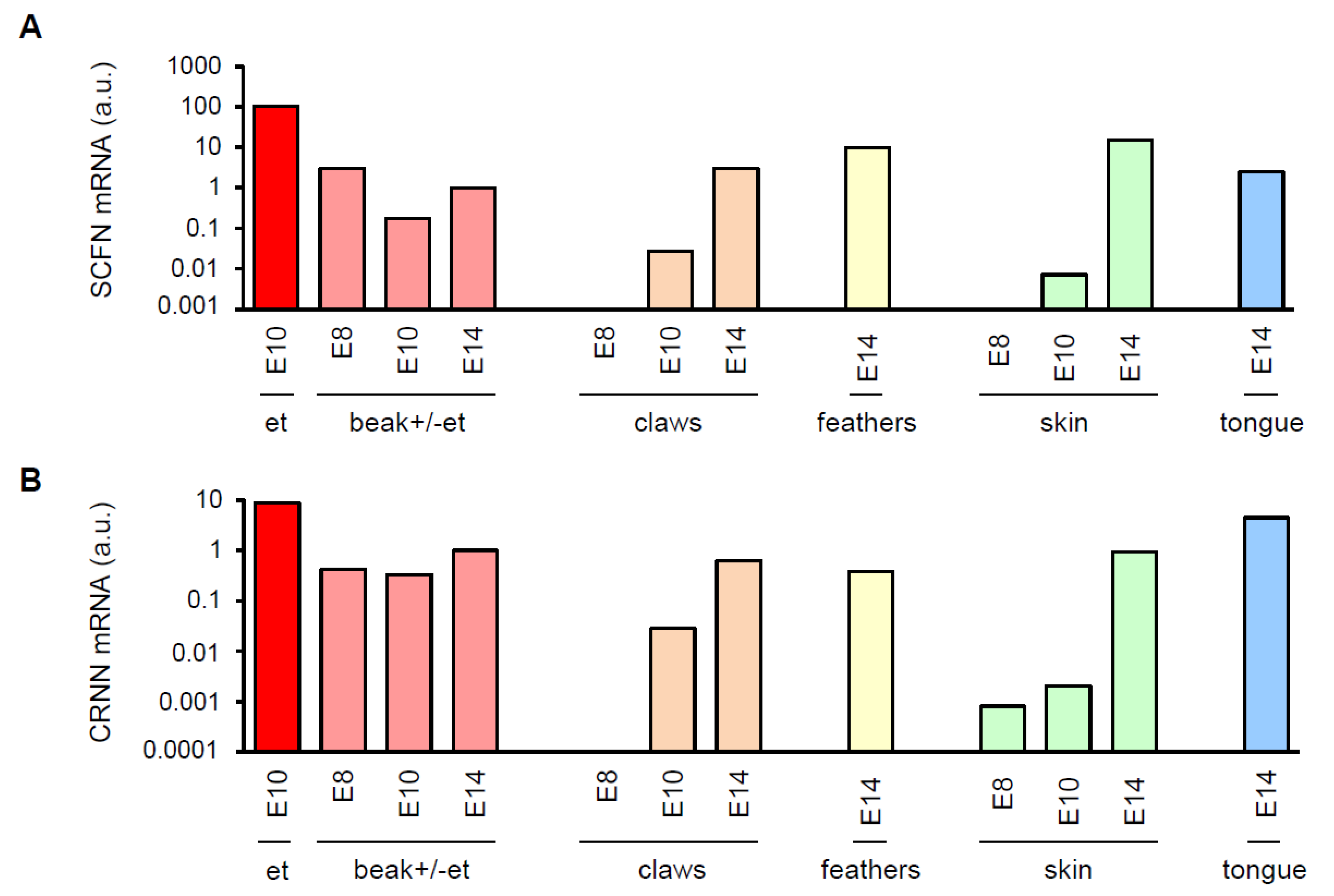
3.1. Avian SFTPs Are Expressed during Embryonic Beak Development of Chicken and Quail
3.2. SCFN Forms Granules in Multilayered Periderm on the Beak
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fons, J.M.; Gaete, M.; Zahradnicek, O.; Landova, M.; Bandali, H.; Khannoon, E.R.; Richman, J.M.; Buchtová, M.; Tucker, A.S.; Landova, M. Getting out of an egg: Merging of tooth germs to create an egg tooth in the snake. Dev. Dyn. 2020, 249, 199–208. [Google Scholar] [CrossRef]
- Lynch, V.J.; Wagner, G.P. Did egg-laying boas break Dollo’s law? Phylogenetic evidence for reversal to oviparity in sand boas (Eryx, Boidae). Evolution 2010, 64, 207–216. [Google Scholar] [CrossRef]
- Kingsbury, J.W.; Allen, V.G.; Rotheram, B.A. The histological structure of the beak in the chick. Anat. Rec. 1953, 116, 95–115. [Google Scholar] [CrossRef]
- Alibardi, L. Cell proliferation, adhesion, and differentiation of keratinocytes in the developing beak and egg-tooth of the turtle Emydura macquarii. Protoplasma 2020, 257, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- Shames, R.B.; Knapp, L.W.; Carver, W.E.; Sawyer, R.H. Region-specific expression of scutate scale type β keratins in the developing chick beak. J. Exp. Zool. 1991, 260, 258–266. [Google Scholar] [CrossRef]
- Alibardi, L. Immunodetection of type I acidic keratins associated to periderm granules during the transition of cornification from embryonic to definitive chick epidermis. Micron 2014, 65, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Ng, C.S.; Yan, J.; Lai, Y.-C.; Chen, C.-K.; Lai, Y.-T.; Wu, S.-M.; Chen, J.-J.; Luo, W.; Widelitz, R.B.; et al. Topographical mapping of α- and β-keratins on developing chicken skin integuments, Functional interaction and evolutionary perspectives. Proc. Natl. Acad. Sci. USA 2015, 112, E6770–E6779. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Stiegler, J.; Wu, P.; Chuong, C.M.; Hu, D.; Balanoff, A.; Zhou, Y.; Xu, X. Heterochronic truncation of odontogenesis in theropod dinosaurs provides insight into the macroevolution of avian beaks. Proc. Natl. Acad. Sci. USA 2017, 114, 10930–10935. [Google Scholar] [CrossRef] [Green Version]
- Nimmagadda, S.; Buchtová, M.; Fu, K.; Geetha-Loganathan, P.; Hosseini-Farahabadi, S.; Trachtenberg, A.J.; Kuo, W.P.; Vesela, I.; Richman, J.M. Identification and functional analysis of novel facial patterning genes in the duplicated beak chicken embryo. Dev. Biol. 2015, 407, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, R.H.; Knapp, L.W.; O’Guin, M.W. The Skin of Birds. Biology of the Integument, Vertebrates. In Epidermis, DERMIS and Appendages; Bereither-Hahn, J., Matoltsy, A.G., Sylvia-Richards, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; Volume 2, pp. 194–237. [Google Scholar]
- Wu, P.; Hou, L.; Plikus, M.; Hughes, M.; Scehnet, J.; Suksaweang, S.; Widelitz, R.; Jiang, T.X.; Chuong, C.M. Evo-Devo of amniote integuments and appendages. Int. J. Dev. Biol. 2004, 48, 249–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibardi, L. Embryonic keratinization in vertebrates in relation to land colonization. Acta Zool. 2009, 90, 1–17. [Google Scholar] [CrossRef]
- Henry, J.; Toulza, E.; Hsu, C.Y.; Pellerin, L.; Balica, S.; Mazereeuw-Hautier, J.; Paul, C.; Serre, G.; Jonca, N.; Simon, M. Update on the epidermal differentiation complex. Front. Biosci. 2012, 17, 1517–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex, cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef]
- Greenwold, M.J.; Sawyer, R.H. Genomic organization and molecular phylogenies of the β (β) keratin multigene family in the chicken (Gallus gallus) and zebra finch (Taeniopygia guttata), implications for feather evolution. BMC Evol. Biol. 2010, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Greenwold, M.J.; Bao, W.; Jarvis, E.D.; Hu, H.; Li, C.; Gilbert, M.T.P.; Zhang, G.; Sawyer, R.H. Dynamic evolution of the α (α) and β (β) keratins has accompanied integument diversification and the adaptation of birds into novel lifestyles. BMC Evol. Biol. 2014, 14, 249. [Google Scholar] [CrossRef] [Green Version]
- Strasser, B.; Mlitz, V.; Hermann, M.; Rice, R.H.; Eigenheer, R.A.; Alibardi, L.; Tschachler, E.; Eckhart, L. Evolutionary origin and diversification of epidermal barrier proteins in amniotes. Mol. Biol. Evol. 2014, 31, 3194–3205. [Google Scholar] [CrossRef] [Green Version]
- Strasser, B.; Mlitz, V.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich proteins in feathers and hair. BMC Evol. Biol. 2015, 15, 82. [Google Scholar] [CrossRef] [Green Version]
- Alibardi, L.; Holthaus, K.B.; Sukseree, S.; Hermann, M.; Tschachler, E.; Eckhart, L. Immunolocalization of a histidine-rich epidermal differentiation protein in the chicken supports the hypothesis of an evolutionary developmental link between the embryonic subperiderm and feather barbs and barbules. PLoS ONE 2016, 11, e0167789. [Google Scholar] [CrossRef]
- Lachner, J.; Ehrlich, F.; Mlitz, V.; Hermann, M.; Alibardi, L.; Tschachler, E.; Eckhart, L. Immunolocalization and phylogenetic profiling of the feather protein with the highest cysteine content. Protoplasma 2019, 256, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Steinert, P.M. A model for the hierarchical structure of the human epidermal cornified cell envelope. Cell Death Differ. 1995, 2, 33–40. [Google Scholar]
- Mildner, M.; Jin, J.; Eckhart, L.; Kezic, S.; Gruber, F.; Barresi, C.; Stremnitzer, C.; Buchberger, M.; Mlitz, V.; Ballaun, C.; et al. Knockdown of filaggrin impairs diffusion barrier function and increases UV sensitivity in a human skin model. J. Investig. Dermatol. 2010, 130, 2286–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlitz, V.; Latreille, J.; Gardinier, S.; Jdid, R.; Drouault, Y.; Hufnagl, P.; Eckhart, L.; Guinot, C.; Tschachler, E. Impact of filaggrin mutations on Raman spectra and biophysical properties of the stratum corneum in mild to moderate atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 983–990. [Google Scholar] [CrossRef] [PubMed]
- McLean, W.H. Filaggrin failure—From ichthyosis vulgaris to atopic eczema and beyond. Br. J. Dermatol. 2016, 175 (Suppl. S2), 4–7. [Google Scholar] [CrossRef] [Green Version]
- Contzler, R.; Favre, B.; Huber, M.; Hohl, D. Cornulin, a new member of the “fused gene” family, is expressed during epidermal differentiation. J. Investig. Dermatol. 2005, 124, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, E.J.; Hamilton, E.H.; Lee, S.C.; Steinert, P. Trichohyalin, a structural protein of hair, tongue, nail, and epidermis. J. Invest. Dermatol. 1993, 101, 65S–71S. [Google Scholar]
- Steinert, P.M.; Parry, D.A.; Marekov, L.N. Trichohyalin mechanically strengthens the hair follicle: Multiple cross-bridging roles in the inner root shealth. J. Biol. Chem. 2003, 278, 41409–41419. [Google Scholar] [CrossRef]
- Basmanav, F.B.Ü.; Cau, L.; Tafazzoli, A.; Méchin, M.C.; Wolf, S.; Romano, M.T.; Valentin, F.; Wiegmann, H.; Huchenq, A.; Kandil, R.; et al. Mutations in three genes encoding proteins involved in hair shaft formation cause uncombable hair syndrome. Am. J. Hum. Genet. 2016, 99, 1292–1304. [Google Scholar] [CrossRef] [Green Version]
- Mlitz, V.; Strasser, B.; Jaeger, K.; Hermann, M.; Ghannadan, M.; Buchberger, M.; Alibardi, L.; Tschachler, E.; Eckhart, L. Trichohyalin-like proteins have evolutionarily conserved roles in the morphogenesis of skin appendages. J. Investig. Dermatol. 2014, 134, 2685–2692. [Google Scholar] [CrossRef] [Green Version]
- Mlitz, V.; Hussain, T.; Tschachler, E.; Eckhart, L. Filaggrin has evolved from an “S100 fused-type protein” (SFTP) gene present in a common ancestor of amphibians and mammals. Exp. Dermatol. 2017, 26, 955–957. [Google Scholar] [CrossRef] [Green Version]
- Holthaus, K.B.; Strasser, B.; Sipos, W.; Schmidt, H.A.; Mlitz, V.; Sukseree, S.; Weissenbacher, A.; Tschachler, E.; Alibardi, L.; Eckhart, L. Comparative genomics identifies epidermal proteins associated with the evolution of the turtle shell. Mol. Biol. Evol. 2016, 33, 726–737. [Google Scholar] [CrossRef]
- Holthaus, K.B.; Mlitz, V.; Strasser, B.; Tschachler, E.; Alibardi, L.; Eckhart, L. Identification and comparative analysis of the epidermal differentiation complex in snakes. Sci. Rep. 2017, 7, 45338. [Google Scholar] [CrossRef] [Green Version]
- Holthaus, K.B.; Alibardi, L.; Tschachler, E.; Eckhart, L. Identification of epidermal differentiation genes of the tuatara provides insights into the early evolution of lepidosaurian skin. Sci. Rep. 2020, 10, 12844. [Google Scholar] [CrossRef]
- Feng, S.; Stiller, J.; Deng, Y.; Armstrong, J.; Fang, Q.; Reeve, A.H.; Xie, D.; Chen, G.; Guo, C.; Faircloth, B.C.; et al. Dense sampling of bird diversity increases power of comparative genomics. Nature 2020, 587, 252–257. [Google Scholar] [CrossRef]
- Nikolay, B.; Plieschnig, J.A.; Šubik, D.; Schneider, J.D.; Schneider, W.J.; Hermann, M. A novel estrogen-regulated avian apolipoprotein. Biochimie 2013, 95, 2445–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeau, N.J.; Minvielle, F.; Ito, S.; Inoue-Murayama, M.; Gourichon, D.; Follett, S.A.; Burke, T.; Mundy, N.I. Characterization of Japanese quail yellow as a genomic deletion upstream of the avian homolog of the mammalian ASIP (agouti) gene. Genetics 2008, 178, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Fischer, H.; Eckhart, L.; Mildner, M.; Jaeger, K.; Buchberger, M.; Ghannadan, M.; Tschachler, E. DNase1L2 degrades nuclear DNA during corneocyte formation. J. Investig. Dermatol. 2007, 127, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Mori, T. Peridermal granules of the chick embryo, histochemical, ultrastructural and immuno-electron microscopical study. Tohoku J. Exp. Med. 1987, 151, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibardi, L.; Mlitz, V.; Eckhart, L. Immunolocalization of scaffoldin, a trichohyalin-like protein, in the epidermis of the chicken embryo. Anat. Rec. 2015, 298, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.A., Jr. Occurrence and timing of egg teeth in birds. Wilson Bull. 1961, 73, 268–278. [Google Scholar]
- Richardson, R.J.; Dixon, J.; Jiang, R.; Dixon, M.J. Integration of IRF6 and Jagged2 signalling is essential for controlling palatal adhesion and fusion competence. Hum. Mol. Genet. 2009, 18, 2632–2642. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Duncan, K.; Helverson, A.; Kumari, P.; Mumm, C.; Xiao, Y.; Carlson, J.C.; Darbellay, F.; Visel, A.; Leslie, E.; et al. Analysis of zebrafish periderm enhancers facilitates identification of a regulatory variant near human KRT8/18. Elife 2020, 9, e51325. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, R.H.; Rogers, L.; Washington, L.; Glenn, T.C.; Knapp, L.W. Evolutionary origin of the feather epidermis. Dev. Dyn. 2005, 232, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Takase, T.; Hirai, Y. Identification of the C-terminal tail domain of AHF/trichohyalin as the critical site for modulation of the keratin filamentous meshwork in the keratinocyte. J. Dermatol. Sci. 2012, 65, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto-Tanaka, M.; Makino, T.; Motoyama, A.; Miyai, M.; Tsuboi, R.; Hibino, T. Multiple pathways are involved in DNA degradation during keratinocyte terminal differentiation. Cell Death Dis. 2014, 5, e1181. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlitz, V.; Hermann, M.; Buchberger, M.; Tschachler, E.; Eckhart, L. The Trichohyalin-Like Protein Scaffoldin Is Expressed in the Multilayered Periderm during Development of Avian Beak and Egg Tooth. Genes 2021, 12, 248. https://doi.org/10.3390/genes12020248
Mlitz V, Hermann M, Buchberger M, Tschachler E, Eckhart L. The Trichohyalin-Like Protein Scaffoldin Is Expressed in the Multilayered Periderm during Development of Avian Beak and Egg Tooth. Genes. 2021; 12(2):248. https://doi.org/10.3390/genes12020248
Chicago/Turabian StyleMlitz, Veronika, Marcela Hermann, Maria Buchberger, Erwin Tschachler, and Leopold Eckhart. 2021. "The Trichohyalin-Like Protein Scaffoldin Is Expressed in the Multilayered Periderm during Development of Avian Beak and Egg Tooth" Genes 12, no. 2: 248. https://doi.org/10.3390/genes12020248
APA StyleMlitz, V., Hermann, M., Buchberger, M., Tschachler, E., & Eckhart, L. (2021). The Trichohyalin-Like Protein Scaffoldin Is Expressed in the Multilayered Periderm during Development of Avian Beak and Egg Tooth. Genes, 12(2), 248. https://doi.org/10.3390/genes12020248