
Accurate Quantification of AAV Vector Genomes by Quantitative PCR

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
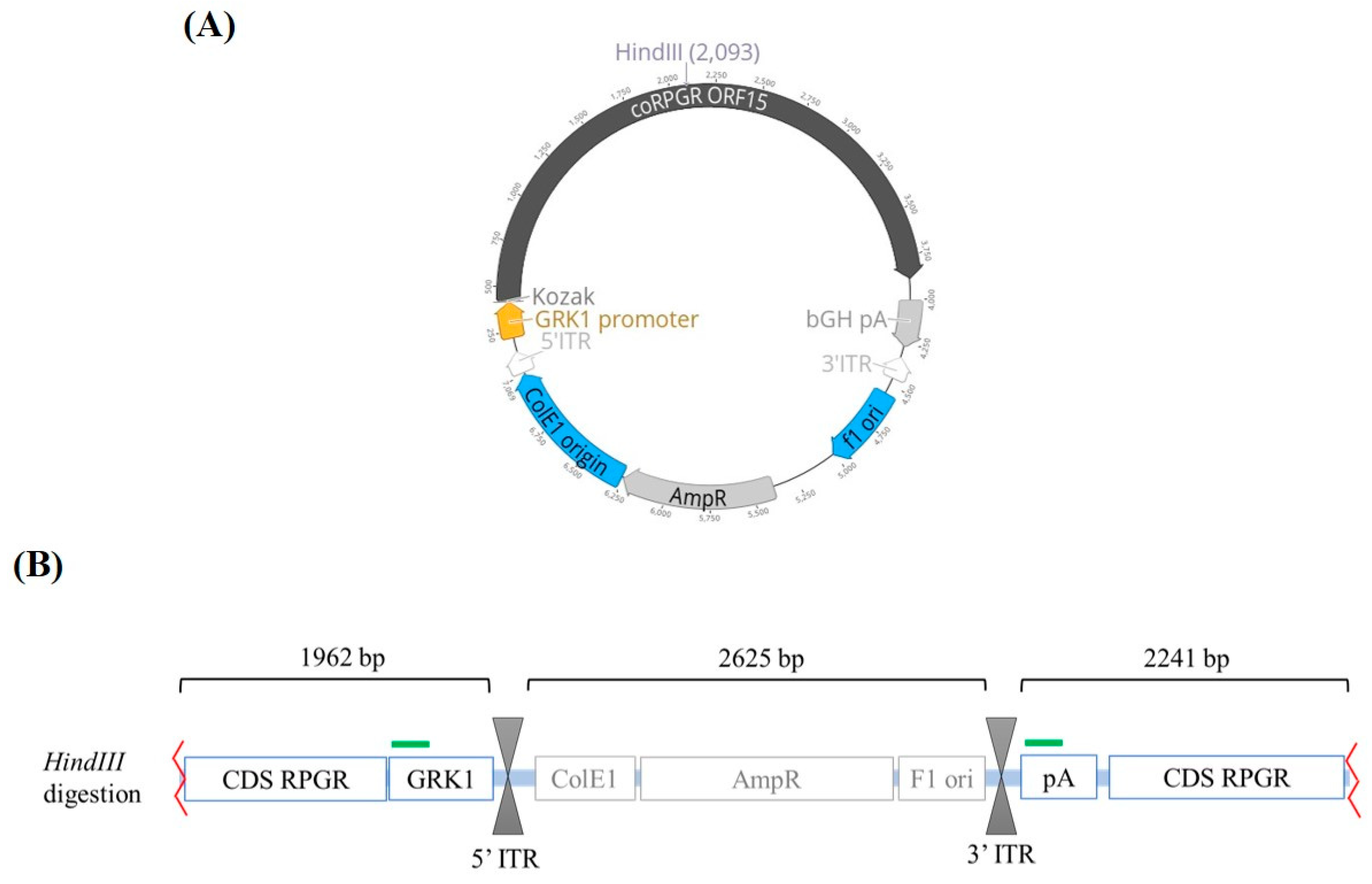
2.1. Preparation of Viral Vector
2.2. AAV Sample Processing
2.3. Analytical Standards for qPCR
2.4. Quantitative PCR Analysis
2.5. Statistical Analysis
3. Results
3.1. Supercoiled DNA Generates Higher Ct Values Than Linearized DNA
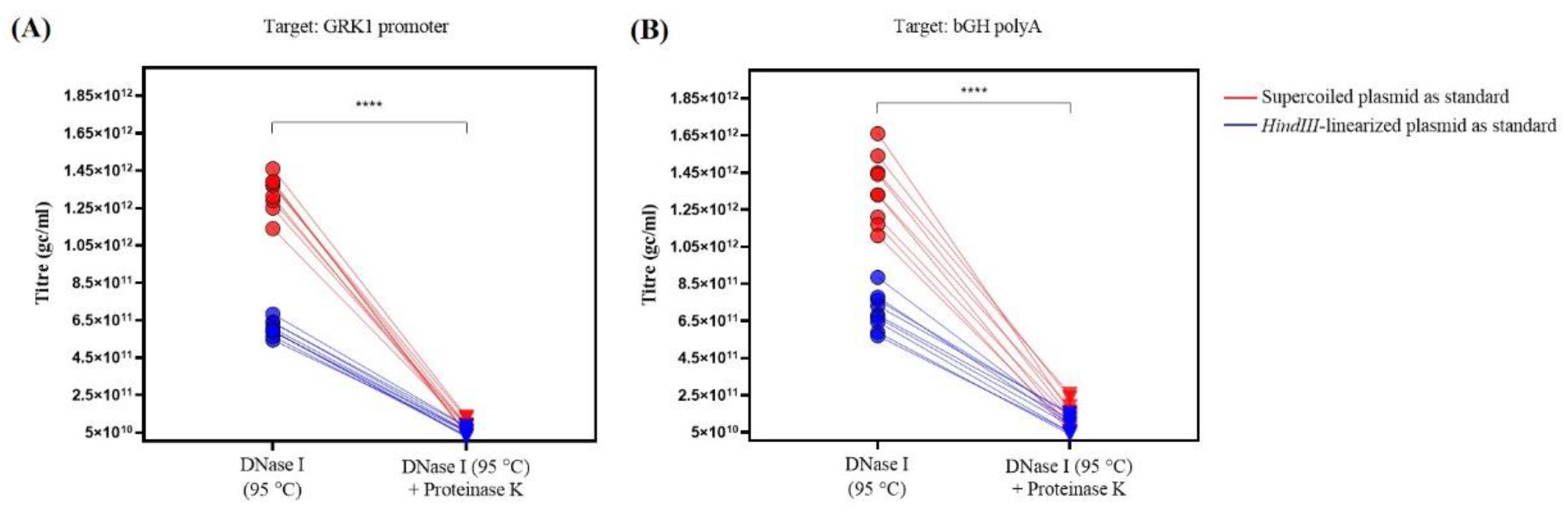
3.2. Additional Treatment with Proteinase K Significantly Influences AAV Titre
3.3. The Conformation of the Reference DNA Influences the AAV Titre
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balakrishnan, B.; Jayandharan, G.R. Basic biology of adeno-associated virus (AAV) vectors used in gene therapy. Curr. Gene Ther. 2014, 14, 86–100. [Google Scholar] [CrossRef]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [Green Version]
- McClements, M.E.; MacLaren, R.E. Adeno-associated Virus (AAV) Dual Vector Strategies for Gene Therapy Encoding Large Transgenes. Yale J. Biol. Med. 2017, 90, 611–623. [Google Scholar]
- Vance, M.A.; Mitchell, A.; Samulski, R.J. AAV Biology, Infectivity and Therapeutic Use from Bench to Clinic. In Gene Therapy—Principles and Challenges; IntechOpen Ltd.: London, UK, 2015. [Google Scholar]
- Balaggan, K.S.; Duran, Y.; Georgiadis, A.; Thaung, C.; Barker, S.E.; Buch, P.K.; MacNeil, A.; Robbie, S.; Bainbridge, J.W.; Smith, A.J.; et al. Absence of ocular malignant transformation after sub-retinal delivery of rAAV2/2 or integrating lentiviral vectors in p53-deficient mice. Gene Ther. 2012, 19, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surace, E.M.; Auricchio, A. Versatility of AAV vectors for retinal gene transfer. Vis. Res. 2008, 48, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLaren, R.E.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Seymour, L.; Clark, K.R.; During, M.J.; Cremers, F.P.M.; Black, G.C.M.; et al. Retinal gene therapy in patients with choroideremia: Initial findings from a phase 1/2 clinical trial. Lancet 2014, 383, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Xue, K.; Jolly, J.K.; Barnard, A.R.; Rudenko, A.; Salvetti, A.P.; Patricio, M.I.; Edwards, T.L.; Groppe, M.; Orlans, H.O.; Tolmachova, T.; et al. Beneficial effects on vision in patients undergoing retinal gene therapy for choroideremia. Nat. Med. 2018, 24, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Cehajic-Kapetanovic, J.; Xue, K.; Martinez-Fernandez de la Camara, C.; Nanda, A.; Davies, A.; Wood, L.J.; Salvetti, A.P.; Fischer, M.D.; Aylward, J.W.; Barnard, A.R.; et al. Initial results from a first-in-human gene therapy trial on X-linked retinitis pigmentosa caused by mutations in RPGR. Nat. Med. 2020, 26, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Patel, U.; Boucher, M.; de Leseleuc, L.; Visintini, S. Voretigene Neparvovec: An Emerging Gene Therapy for the Treatment of Inherited Blindness. In CADTH Issues in Emerging Health Technologies; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2016; pp. 1–11. [Google Scholar]
- MacLaren, R.E. Gene Therapy for Retinal Disease: What Lies Ahead. Ophthalmologica 2015, 234, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fernandez De La Camara, C.; Nanda, A.; Salvetti, A.P.; Fischer, M.D.; MacLaren, R.E. Gene therapy for the treatment of X-linked retinitis pigmentosa. Expert Opin. Orphan Drugs 2018, 6, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.F. Manufacturing and characterizing AAV-based vectors for use in clinical studies. Gene Ther. 2008, 15, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, A.; Leuchs, B.; Katus, H.A.; Rommelaere, J.; Muller, O.J. Protocol for efficient generation and characterization of adeno-associated viral (AAV) vectors. Hum. Gene Ther. Methods 2017, 28, 235–246. [Google Scholar] [CrossRef]
- Aurnhammer, C.; Haase, M.; Muether, N.; Hausl, M.; Rauschhuber, C.; Huber, I.; Nitschko, H.; Busch, U.; Sing, A.; Ehrhardt, A.; et al. Universal real-time PCR for the detection and quantification of adeno-associated virus serotype 2-derived inverted terminal repeat sequences. Hum. Gene Ther. Methods 2012, 23, 18–28. [Google Scholar] [CrossRef]
- Dobnik, D.; Kogovsek, P.; Jakomin, T.; Kosir, N.; Tusek Znidaric, M.; Leskovec, M.; Kaminsky, S.M.; Mostrom, J.; Lee, H.; Ravnikar, M. Accurate Quantification and Characterization of Adeno-Associated Viral Vectors. Front. Microbiol. 2019, 10, 1570. [Google Scholar] [CrossRef] [Green Version]
- Francois, A.; Bouzelha, M.; Lecomte, E.; Broucque, F.; Penaud-Budloo, M.; Adjali, O.; Moullier, P.; Blouin, V.; Ayuso, E. Accurate Titration of Infectious AAV Particles Requires Measurement of Biologically Active Vector Genomes and Suitable Controls. Mol. Ther. Methods Clin. Dev. 2018, 10, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Salvetti, A.P.; Patricio, M.I.; Barnard, A.R.; Orlans, H.O.; Hickey, D.G.; MacLaren, R.E. Impact of Vital Dyes on Cell Viability and Transduction Efficiency of AAV Vectors Used in Retinal Gene Therapy Surgery: An In Vitro and In Vivo Analysis. Transl. Vis. Sci. Technol. 2017, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Karlen, Y.; McNair, A.; Perseguers, S.; Mazza, C.; Mermod, N. Statistical significance of quantitative PCR. BMC Bioinform. 2007, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.D.; McClements, M.E.; Martinez-Fernandez de la Camara, C.; Bellingrath, J.S.; Dauletbekov, D.; Ramsden, S.C.; Hickey, D.G.; Barnard, A.R.; MacLaren, R.E. Codon-Optimized RPGR Improves Stability and Efficacy of AAV8 Gene Therapy in Two Mouse Models of X-Linked Retinitis Pigmentosa. Mol. Ther. 2017, 25, 1854–1865. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Liu, Y.; Noor, A.F.; Tran, J.; Li, R. Characteristics and advantages of adeno-associated virus vector-mediated gene therapy for neurodegenerative diseases. Neural Regen. Res. 2019, 14, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.A.; Shalaev, E.; Karami, T.K.; Cunningham, J.; Slater, N.K.H.; Rivers, H.M. Pharmaceutical Development of AAV-Based Gene Therapy Products for the Eye. Pharm. Res. 2018, 36, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdera, H.C.; Kuranda, K.; Mingozzi, F. AAV Vector Immunogenicity in Humans: A Long Journey to Successful Gene Transfer. Mol. Ther. 2020, 28, 723–746. [Google Scholar] [CrossRef]
- Bainbridge, J.W.; Mehat, M.S.; Sundaram, V.; Robbie, S.J.; Barker, S.E.; Ripamonti, C.; Georgiadis, A.; Mowat, F.M.; Beattie, S.G.; Gardner, P.J.; et al. Long-term effect of gene therapy on Leber’s congenital amaurosis. N. Engl. J. Med. 2015, 372, 1887–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichel, F.F.; Dauletbekov, D.L.; Klein, R.; Peters, T.; Ochakovski, G.A.; Seitz, I.P.; Wilhelm, B.; Ueffing, M.; Biel, M.; Wissinger, B.; et al. AAV8 Can Induce Innate and Adaptive Immune Response in the Primate Eye. Mol. Ther. 2017, 25, 2648–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Farina, I.; Di Scala, M.; Vanrell, L.; Olague, C.; Vales, A.; High, K.A.; Prieto, J.; Mingozzi, F.; Gonzalez-Aseguinolaza, G. IL12-mediated liver inflammation reduces the formation of AAV transcriptionally active forms but has no effect over preexisting AAV transgene expression. PLoS ONE 2013, 8, e67748. [Google Scholar] [CrossRef]
- Xiong, W.; Wu, D.M.; Xue, Y.; Wang, S.K.; Chung, M.J.; Ji, X.; Rana, P.; Zhao, S.R.; Mai, S.; Cepko, C.L. AAV cis-regulatory sequences are correlated with ocular toxicity. Proc. Natl. Acad. Sci. USA 2019, 116, 5785–5794. [Google Scholar] [CrossRef] [Green Version]
- Khabou, H.; Cordeau, C.; Pacot, L.; Fisson, S.; Dalkara, D. Dosage Thresholds and Influence of Transgene Cassette in Adeno-Associated Virus-Related Toxicity. Hum. Gene Ther. 2018, 29, 1235–1241. [Google Scholar] [CrossRef]
- Bennett, A.; Patel, S.; Mietzsch, M.; Jose, A.; Lins-Austin, B.; Yu, J.C.; Bothner, B.; McKenna, R.; Agbandje-McKenna, M. Thermal Stability as a Determinant of AAV Serotype Identity. Mol. Ther. Methods Clin. Dev. 2017, 6, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Rayaprolu, V.; Kruse, S.; Kant, R.; Venkatakrishnan, B.; Movahed, N.; Brooke, D.; Lins, B.; Bennett, A.; Potter, T.; McKenna, R.; et al. Comparative analysis of adeno-associated virus capsid stability and dynamics. J. Virol. 2013, 87, 13150–13160. [Google Scholar] [CrossRef] [Green Version]
- Werling, N.J.; Satkunanathan, S.; Thorpe, R.; Zhao, Y. Systematic Comparison and Validation of Quantitative Real-Time PCR Methods for the Quantitation of Adeno-Associated Viral Products. Hum. Gene Ther. Methods 2015, 26, 82–92. [Google Scholar] [CrossRef]
- Hou, Y.; Zhang, H.; Miranda, L.; Lin, S. Serious overestimation in quantitative PCR by circular (supercoiled) plasmid standard: Microalgal pcna as the model gene. PLoS ONE 2010, 5, e9545. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Chen, Y.C.; Pan, T.M. Quantification bias caused by plasmid DNA conformation in quantitative real-time PCR assay. PLoS ONE 2011, 6, e29101. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target | Label | Sequence (5′→3′) |
|---|---|---|
| GRK1 promoter | Forward primer | TCTCTTAAGGTAGCCCCGG |
| Reverse primer | ATCCGATTAGATCATTCTGCCC | |
| Taqman Probe (FAM) | CCTCACTTTTCCCCTGAGAAGGACA | |
| bGH polyA | Forward primer | CTCGACTGTGCCTTCTAGTTG |
| Reverse primer | ACCTACTCAGACAATGCGATG | |
| Taqman Probe (FAM) | TGCCACTCCCACTGTCCTTTCC |
| GRK1 Promoter | bGH polyA | |||
|---|---|---|---|---|
| Supercoiled | HindIII-Linearized | Supercoiled | HindIII-Linearized | |
| Ct range | 12.66–28.87 | 10.72–28.91 | 13.21–28.91 | 10.81–28.53 |
| R square | 0.9953 | 0.9993 | 0.9923 | 0.9989 |
| Slope | −3.153 | −3.607 | −3.080 | −3.487 |
| Intercept | 39.13 | 41.31 | 38.81 | 40.42 |
| Efficiency | 107.6% | 89.4% | 110.2% | 93.5% |
| Sample Treatment | Target | AAV Titre (gc/mL) | Significance (p Value) | Ratio (Supercoiled/Linearized) | |
|---|---|---|---|---|---|
| Supercoiled Plasmid as Standard | HindIII-Linearized Plasmid as Standard | ||||
| DNase I (95 °C) | GRK1 promoter | 1.33 ± 0.03 × 1012 (7.1%) | 6.06 ± 0.14 × 1011 (7.0%) | **** (p < 0.0001) | 2.20 ± 0.03 |
| bGH polyA | 1.36 ± 0.06 × 1012 (13.3%) | 7.02 ±0.33 × 1011 (14.2%) | **** (p < 0.0001) | 1.94 ± 0.03 | |
| DNase I (95 °C) +Proteinase K | GRK1 promoter | 8.21 ± 1.11 × 1010 (40.6%) | 5.42 ± 0.68 × 1010 (37.7%) | ns (p = 0.063) | 1.50 ± 0.02 |
| bGH polyA | 1.44 ± 0.24 × 1011 (50.0%) | 9.51 ± 1.37 × 1010 (43.3%) | ns (p = 0.162) | 1.48 ± 0.04 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Fernandez de la Camara, C.; McClements, M.E.; MacLaren, R.E. Accurate Quantification of AAV Vector Genomes by Quantitative PCR. Genes 2021, 12, 601. https://doi.org/10.3390/genes12040601
Martinez-Fernandez de la Camara C, McClements ME, MacLaren RE. Accurate Quantification of AAV Vector Genomes by Quantitative PCR. Genes. 2021; 12(4):601. https://doi.org/10.3390/genes12040601
Chicago/Turabian StyleMartinez-Fernandez de la Camara, Cristina, Michelle E. McClements, and Robert E. MacLaren. 2021. "Accurate Quantification of AAV Vector Genomes by Quantitative PCR" Genes 12, no. 4: 601. https://doi.org/10.3390/genes12040601
APA StyleMartinez-Fernandez de la Camara, C., McClements, M. E., & MacLaren, R. E. (2021). Accurate Quantification of AAV Vector Genomes by Quantitative PCR. Genes, 12(4), 601. https://doi.org/10.3390/genes12040601