
Silver Nanoparticles as Modulators of Myogenesis-Related Gene Expression in Chicken Embryos

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticles
2.2. Experimental Design
2.3. Gene Expression Analysis Using Real-Time PCR
2.4. Western Blot Analysis
2.5. Histological Examination of Breast Muscle Tissue
2.6. Statistical Analysis
3. Results
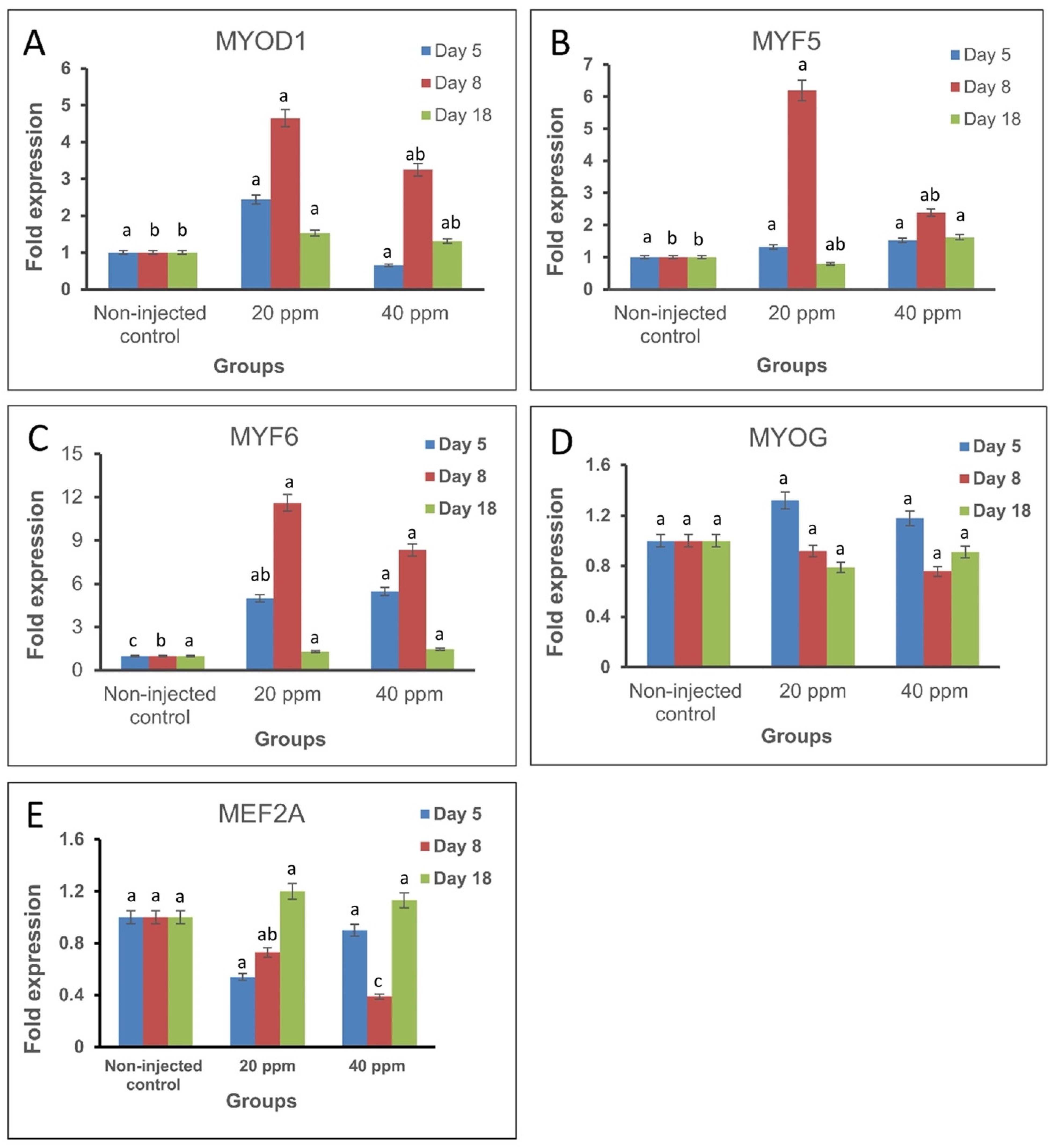
3.1. Gene Expression Analysis
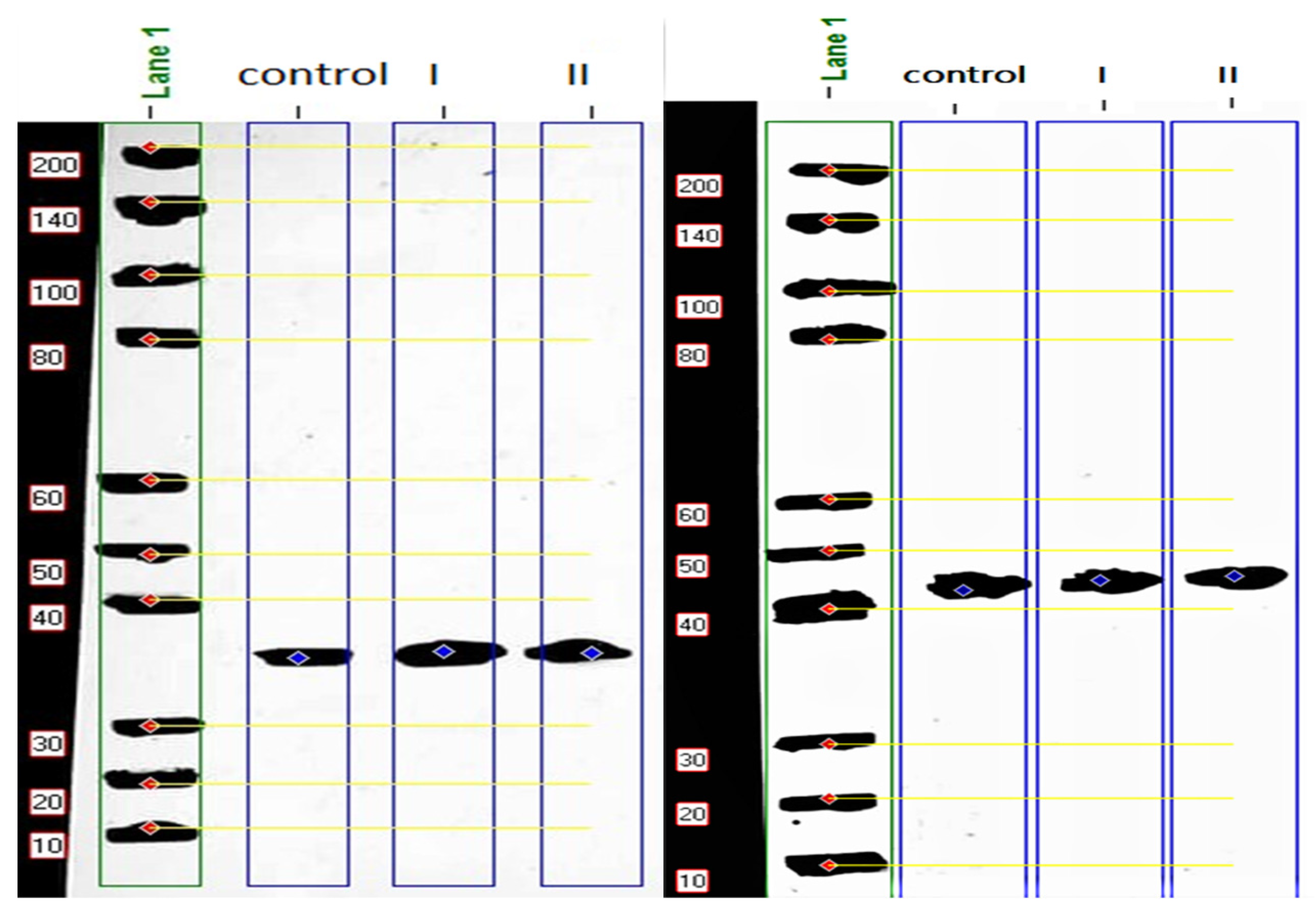
3.2. Western Blot Analysis
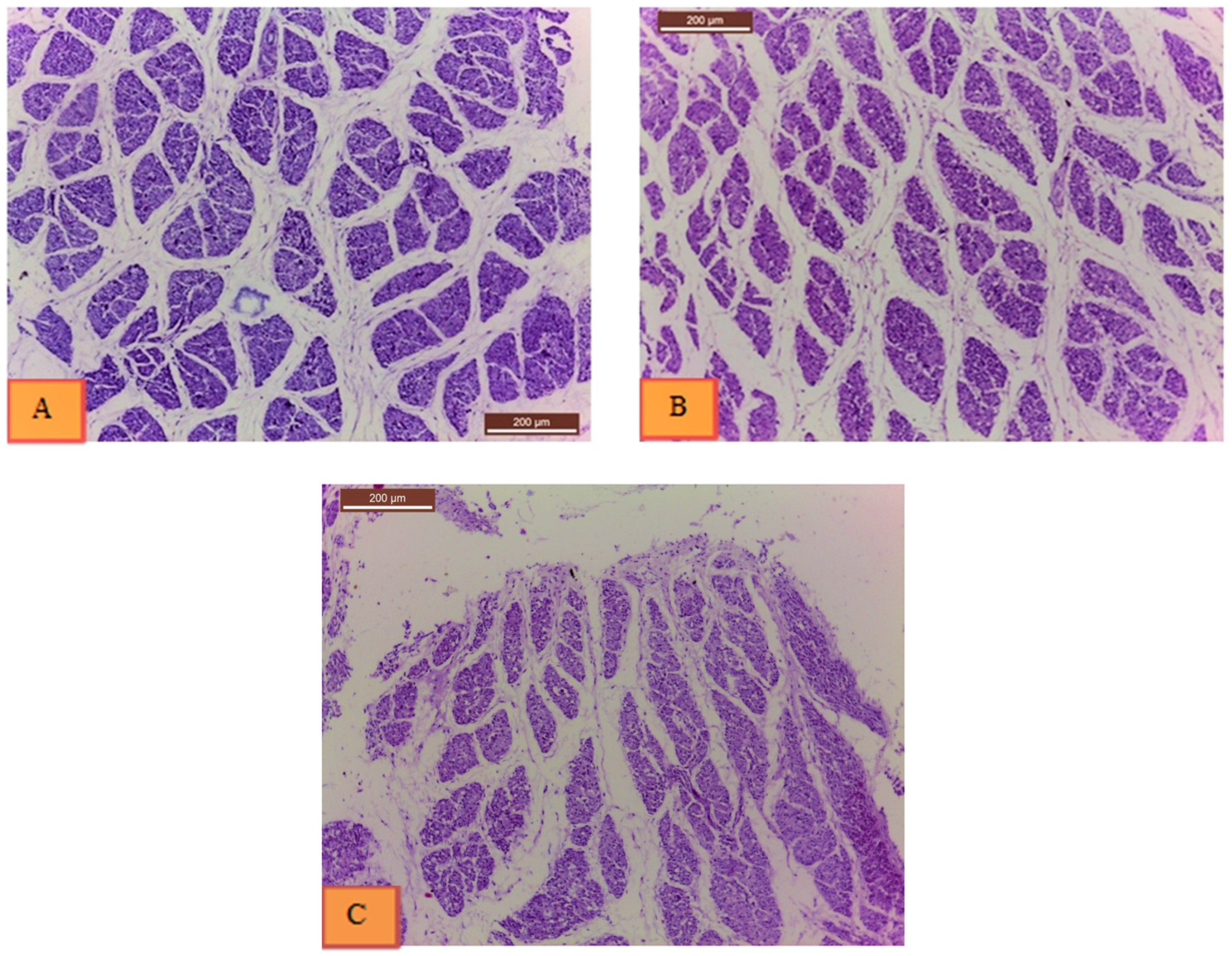
3.3. Histological Examination
3.4. Hatchability Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sailor, M.J.; Park, J.H. Hybrid nanoparticles for detection and treatment of cancer. Adv. Mater. 2012, 24, 3779–3802. [Google Scholar] [CrossRef]
- Abdelsalam, M.; Al-Homidan, I.; Ebeid, T.; Abou-Emera, O.; Mostafa, M.; El-Razik, A.; Shehab-El-Deen, M.; Abdel Ghani, S.; Fathi, M. Effect of silver nanoparticle administration on productive performance, blood parameters, antioxidative status, and silver residues in growing rabbits under hot climate. Animals 2019, 9, 845. [Google Scholar] [CrossRef] [Green Version]
- Hassanen, E.I.; Morsy, E.A.; Hussien, A.M.; Ibrahim, M.A.; Farroh, K.Y. The effect of different concentrations of gold nanoparticles on growth performance, toxicopathological and immunological parameters of broiler chickens. Biosci. Rep. 2020, 40, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Z. Effect of chromium nanoparticle on growth performance, carcass characteristics, pork quality and tissue chromium in finishing pigs. Asian-Australas J. Anim. Sci. 2004, 17, 1118–1122. [Google Scholar] [CrossRef]
- Vadalasetty, K.P.; Lauridsen, C.; Engberg, R.M.; Vadalasetty, R.; Kutwin, M.; Chwalibog, A.; Sawosz, E. Influence of silver nanoparticles on growth and health of broiler chickens after infection with Campylobacter jejuni. BMC Vet. Res. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pineda, L.; Sawosz, E.; Hotowy, A.; Elnif, J.; Sawosz, F.; Ali, A.; Chwalibog, A. Effect of nanoparticles of silver and gold on metabolic rate and development of broiler and layer embryos. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 161, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Manshian, B.; Jenkins, G.J.; Griffiths, S.M.; Williams, P.M.; Maffeis, T.G.; Wright, C.J.; Doak, S.H. Nanogenotoxicology: The DNA damaging potential of engineered nanomaterials. Biomaterials 2009, 30, 3891–3914. [Google Scholar] [CrossRef]
- Sawosz, E.; Grodzik, M.; Lisowski, P.; Zwierzchowski, L.; Niemiec, T.; Zielińska, M.; Szmidt, M.; Chwalibog, A. Influence of hydrocolloids of Ag, Au, and Ag/Cu alloy nanoparticles on the inflammatory state at transcriptional level. Bull. Vet. Inst. Pulawy 2010, 54, 81–85. [Google Scholar]
- Sawosz, F.; Pineda, L.M.; Hotowy, A.M.; Hyttel, P.; Sawosz, E.; Szmidt, M.; Niemiec, T.; Chwalibog, A. Nano-nutrition of chicken embryos. The effect of silver nanoparticles and glutamine on molecular responses, and the morphology of pectoral muscle. Balt. J. Comp. Clin. Syst. Biol. 2012, 2, 29–45. [Google Scholar] [CrossRef]
- Sawosz, F.; Pineda, L.; Hotowy, A.; Jaworski, S.; Prasek, M.; Sawosz, E.; Chwalibog, A. Nano-nutrition of chicken embryos. The effect of silver nanoparticles and ATP on expression of chosen genes involved in myogenesis. Arch. Anim. Nutr. 2013, 67, 347–355. [Google Scholar] [CrossRef]
- Pineda, L.; Sawosz, E.; Lauridsen, C.; Engberg, R.M.; Elnif, J.; Hotowy, A.; Sawosz, F.; Chwalibog, A. Influence of in ovo injection and subsequent provision of silver nanoparticles on growth performance, microbial profile, and immune status of broiler chickens. Open Access Anim. Physiol. 2012, 4, 1–8. [Google Scholar]
- Sweetman, D. The myogenic regulatory factors: Critical determinants of muscle identity in development, growth and regeneration. In Skeletal Muscle-From Myogenesis to Clinical Relations; IntechOpen: London, UK, 2012. [Google Scholar]
- Bajard, L.; Relaix, F.; Lagha, M.; Rocancourt, D.; Daubas, P.; Buckingham, M.E. A novel genetic hierarchy functions during hypaxial myogenesis: PAX3 directly activates MYF5 in muscle progenitor cells in the limb. Genes Dev. 2006, 20, 2450–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, T.; Bober, E.; Rudnicki, M.A.; Jaenisch, R.; Arnold, H.-H. MYOD expression marks the onset of skeletal myogenesis in MYF-5 mutant mice. Development 1994, 120, 3083–3092. [Google Scholar] [PubMed]
- Buckingham, M.; Mayeuf, A. Skeletal muscle development. In Muscle; Elsevier: Amsterdam, The Netherlands, 2012; pp. 749–762. [Google Scholar]
- Nabeshima, Y.; Hanaoka, K.; Hayasaka, M.; Esuml, E.; Li, S.; Nonaka, I.; Nabeshima, Y.-i. Myogenin gene disruption results in perinatal lethality because of severe muscle defect. Nature 1993, 364, 532. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L.; Olson, E.N. Transcriptional control of muscle development by myocyte enhancer factor-2 (MEF2) proteins. Annu. Rev. Cell Dev. Biol. 1998, 14, 167–196. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L.; Molkentin, J.D.; Olson, E.N. Multiple roles for the MYOD basic region in transmission of transcriptional activation signals and interaction with MEF2. Mol. Cell. Biol. 1998, 18, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.; Bober, E.; Buschhausen-Denker, G.; Kohtz, S.; Grzeschik, K.-H.; Arnold, H.H.; Kotz, S. Differential expression of myogenic determination genes in muscle cells: Possible autoactivation by the MYF gene products. EMBO J. 1989, 8, 3617–3625. [Google Scholar] [CrossRef]
- Hotowy, A.; Sawosz, E.; Pineda, L.; Sawosz, F.; Grodzik, M.; Chwalibog, A. Silver nanoparticles administered to chicken affect VEGFA and FGF2 gene expression in breast muscle and heart. Nanoscale Res. Lett. 2012, 7, 418. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.B.; Lam, K.; Burrell, R.E. Wound management in an era of increasing bacterial antibiotic resistance: A role for topical silver treatment. Am. J. Infect. Control 1998, 26, 572–577. [Google Scholar] [CrossRef]
- Chwalibog, A.; Sawosz, E.; Hotowy, A.; Szeliga, J.; Mitura, S.; Mitura, K.; Grodzik, M.; Orlowski, P.; Sokolowska, A. Visualization of interaction between inorganic nanoparticles and bacteria or fungi. Int. J. Nanomed. 2010, 5, 1085. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Salmanzadeh, M.; Ebrahimnezhad, Y.; Shahryar, H.A.; Lotfi, A. The effects of in ovo injection of L-threonine in broiler breeder eggs on characters of hatching and growth performance broiler chickens. Eur. J. Exp. Biol. 2011, 1, 164–168. [Google Scholar]
- Sawosz, E.; Grodzik, M.; Zielińska, M.; Niemiec, T.; Olszańska, B.; Chwalibog, A. Nanoparticles of silver do not affect growth, development and DNA oxidative damage in chicken embryos. Eur. Poult. Sci. 2009, 73, 208–213. [Google Scholar]
- Chen, H.; Weiss, J.; Shahidi, F. Nanotechnology in nutraceuticals and functional foods. Food Technol. 2006, 60, 30–36. [Google Scholar]
- Mahmoud, U.T. Silver nanoparticles in poultry production. J. Adv. Vet. Res. 2012, 2, 303–306. [Google Scholar]
- Csete, M.; Walikonis, J.; Slawny, N.; Wei, Y.; Korsnes, S.; Doyle, J.C.; Wold, B. Oxygen-mediated regulation of skeletal muscle satellite cell proliferation and adipogenesis in culture. J. Cell. Physiol. 2001, 189, 189–196. [Google Scholar] [CrossRef]
- Zielinska, M.; Sawosz, E.; Grodzik, M.; Balcerak, M.; Wierzbicki, M.; Skomial, J.; Sawosz, F.; Chwalibog, A. Effect of taurine and gold nanoparticles on the morphological and molecular characteristics of muscle development during chicken embryogenesis. Arch. Anim. Nutr. 2012, 66, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, O.; Fujisawa-Sehara, A.; Nabeshima, Y.-i.; Periasamy, M. Expression of myogenic factors in denervated chicken breast muscle: Isolation of the chicken MYF5 gene. Nucleic Acids Res. 1993, 21, 2503–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, J.; Perry, R.L.; Asakura, A.; Rudnicki, M.A. MYOD induces myogenic differentiation through cooperation of its NH2-and COOH-terminal regions. J. Cell Biol. 2005, 171, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef]
- Fong, A.P.; Tapscott, S.J. Skeletal muscle programming and re-programming. Curr. Opin. Genet. Dev. 2013, 23, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Behringer, R.R.; Olson, E.N. Inactivation of the myogenic bHLH gene MRF4 results in up-regulation of myogenin and rib anomalies. Genes Dev. 1995, 9, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.-O.; Bober, E.; Lyons, G.; Arnold, H.; Buckingham, M. Early expression of the myogenic regulatory gene, MYF-5, in precursor cells of skeletal muscle in the mouse embryo. Development 1991, 111, 1097–1107. [Google Scholar]
- Moretti, I.; Ciciliot, S.; Dyar, K.A.; Abraham, R.; Murgia, M.; Agatea, L.; Akimoto, T.; Bicciato, S.; Forcato, M.; Pierre, P. MRF4 negatively regulates adult skeletal muscle growth by repressing MEF2 activity. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, A.; Goldfine, I.D. Role of myogenin in myoblast differentiation and its regulation by fibroblast growth factor. J. Biol. Chem. 1990, 265, 5960–5963. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene * | Accession Number | Primer Sequence * | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| MYOD1 | NM_204214.2 | FP: 5′ GAATCACCAAATGACCCAAAG 3′ RP: 5′ CTCCACTGTCACTCAGGTTTC 3′ | 185 | This study |
| MYF5 | NM_001030363 | FP: 5′ AGGAGGCTGAAGAAAGTGAACC 3′ RP: 5′ TAGTTCTCCACCTGTTCCCTCA 3′ | 155 | This study |
| MYF6 | NM_001030746 | FP: 5′ CCCCTTCAGCTTCAGCCC 3′ RP: 5′ CTCATTTCTCCACCGCCTCTTC 3′ | 242 | This study |
| MYOG | N M_204184 | FP: 5′ AATCCTTTCCCACTCCTCTCCA 3′ RP: 5′ TTGGTCGAAGAGCAACTTGG 3′ | 176 | This study |
| MEF2A | NM_204864 | FP: 5′ TCGGTGCGAAGTTTTCCTCT 3′ RP: 5′ CTGTTCCGTTCGTCCATTATTC 3′ | 250 | This study |
| ACTB | 396526 | FP: 5′ GTCCACCTTCCAGCAGATGT 3′ RP: 5′ ATAAAGCCATGCCAATCTCG 3′ | 169 | [20] |
| Groups | MYOD1 * | MYF5 * | MYF6 * | MYOG * | MEF2A * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D 5 | D 8 | D 18 | D 5 | D 8 | D 18 | D 5 | D 8 | D 18 | D 5 | D 8 | D 18 | D 5 | D 8 | D 18 | |
| Non-injected control | 1.00 a ± 0.00 | 1.00 b ± 0.00 | 1.00 b ± 0.00 | 1.00 a ± 0.00 | 1.00 b ± 0.00 | 1.00 ab ± 0.00 | 1.00 c ± 0.00 | 1.00 b ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 | 1.00 a ± 0.00 |
| 20 ppm | 2.44 a ± 1.16 | 4.65 a ± 0.90 | 1.53 a ± 0.18 | 1.32 a ± 0.96 | 6.19 a ± 2.37 | 0.79 ab ± 0.18 | 5.00 ab ± 0.47 | 11.61 a ± 1.13 | 1.30 a ± 0.16 | 1.32 a ± 0.18 | 0.92 a ± 0.14 | 0.79 a ± 0.09 | 0.54 a ± 0.48 | 0.73 ab ± 0.29 | 1.20 a ± 0.22 |
| 40 ppm | 0.65 a ± 0.08 | 3.25 ab ± 1.19 | 1.31 ab ± 0.19 | 1.52 a ± 0.50 | 2.39 ab ± 0.21 | 1.62 a ± 0.39 | 5.49 a ± 0.70 | 8.33 a ± 0.84 | 1.46 a ± 0.08 | 1.18 a ± 0.31 | 0.76 a ± 0.09 | 0.91 a ± 0.17 | 0.90 a ± 0.14 | 0.39 bc ± 0.05 | 1.13 a ± 0.38 |
| Marker | β -Actin | Non-Injected Control | Treatment I | Treatment II | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Band No | Lane % | MW (kDa) | Band No | Lane % | MW (kDa) | Band No | Lane % | MW (kDa) | Band No | Lane % | MW (kDa) | Band No | Lane % | MW (kDa) |
| 1 | 9.21 | 200.000 | 1 | 98.90 | 44.47 | 1 | 49.98 | 34.230 | 1 | 71.31 | 34.148 | 1 | 68.43 | 34.314 |
| Group | Count * | Cross-Sectional Area * (µm2) | Average Size* (µm2) | % Area * |
|---|---|---|---|---|
| Non-injected control | 3283.22 b ± 300.08 | 30,584.33 a ± 1978.87 | 10.16 a ± 1.31 | 36.31 a ± 2.30 |
| 20 ppm | 4314.11 a ± 387.06 | 26,722.33 ab ± 994.7 | 6.51 b ± 0.62 | 30.73 b ± 1.19 |
| 40 ppm | 2216.56 c ± 213.99 | 24,214.33 b ± 1798.54 | 12.45 a ± 1.74 | 30.16 b ± 1.50 |
| Groups | Egg Transported to Hatcher (n) | Infertile Eggs (n) | Unhatched Chicks (n) | Hatched Chicks (n) | Scientific Hatchability % |
|---|---|---|---|---|---|
| Injected control | 25 | 3 | 0 | 22 | 22/25 = 88.0 |
| Treatment I (20 ppm) | 24 | 3 | 1 | 20 | 20/21 = 95.23 |
| Treatment II (40 ppm) | 27 | 0 | 3 | 24 | 24/27 = 88.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Husseiny, W.A.; Hassanin, A.A.I.; El Nabtiti, A.A.S.; Khalil, K.; Elaswad, A. Silver Nanoparticles as Modulators of Myogenesis-Related Gene Expression in Chicken Embryos. Genes 2021, 12, 629. https://doi.org/10.3390/genes12050629
Husseiny WA, Hassanin AAI, El Nabtiti AAS, Khalil K, Elaswad A. Silver Nanoparticles as Modulators of Myogenesis-Related Gene Expression in Chicken Embryos. Genes. 2021; 12(5):629. https://doi.org/10.3390/genes12050629
Chicago/Turabian StyleHusseiny, Walaa A., Abeer A. I. Hassanin, Adel A. S. El Nabtiti, Karim Khalil, and Ahmed Elaswad. 2021. "Silver Nanoparticles as Modulators of Myogenesis-Related Gene Expression in Chicken Embryos" Genes 12, no. 5: 629. https://doi.org/10.3390/genes12050629
APA StyleHusseiny, W. A., Hassanin, A. A. I., El Nabtiti, A. A. S., Khalil, K., & Elaswad, A. (2021). Silver Nanoparticles as Modulators of Myogenesis-Related Gene Expression in Chicken Embryos. Genes, 12(5), 629. https://doi.org/10.3390/genes12050629