
Identification of Potential Key lncRNAs in the Context of Mouse Myeloid Differentiation by Systematic Transcriptomics Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA-Seq Data Collection and Data Analysis
2.2. Differential Expression Analysis
2.3. Gene Co-Expression Network Analysis
2.4. GO Enrichment Analysis
2.5. Cell Culture
2.6. Total RNA Extraction
2.7. Reverse Transcription and Real-Time qPCR
2.8. RNA-Seq
2.9. Gene Knockdown Using ASOs
2.10. CRISPRi
2.11. Flow Cytometry Analysis
2.12. Western Blotting
2.13. Antibodies Used for Flow Cytometry Analysis and Western Blot
3. Results
3.1. Gene Co-Expression Network Analysis and Potential Key lncRNAs in the Context of Myeloid Differentiation
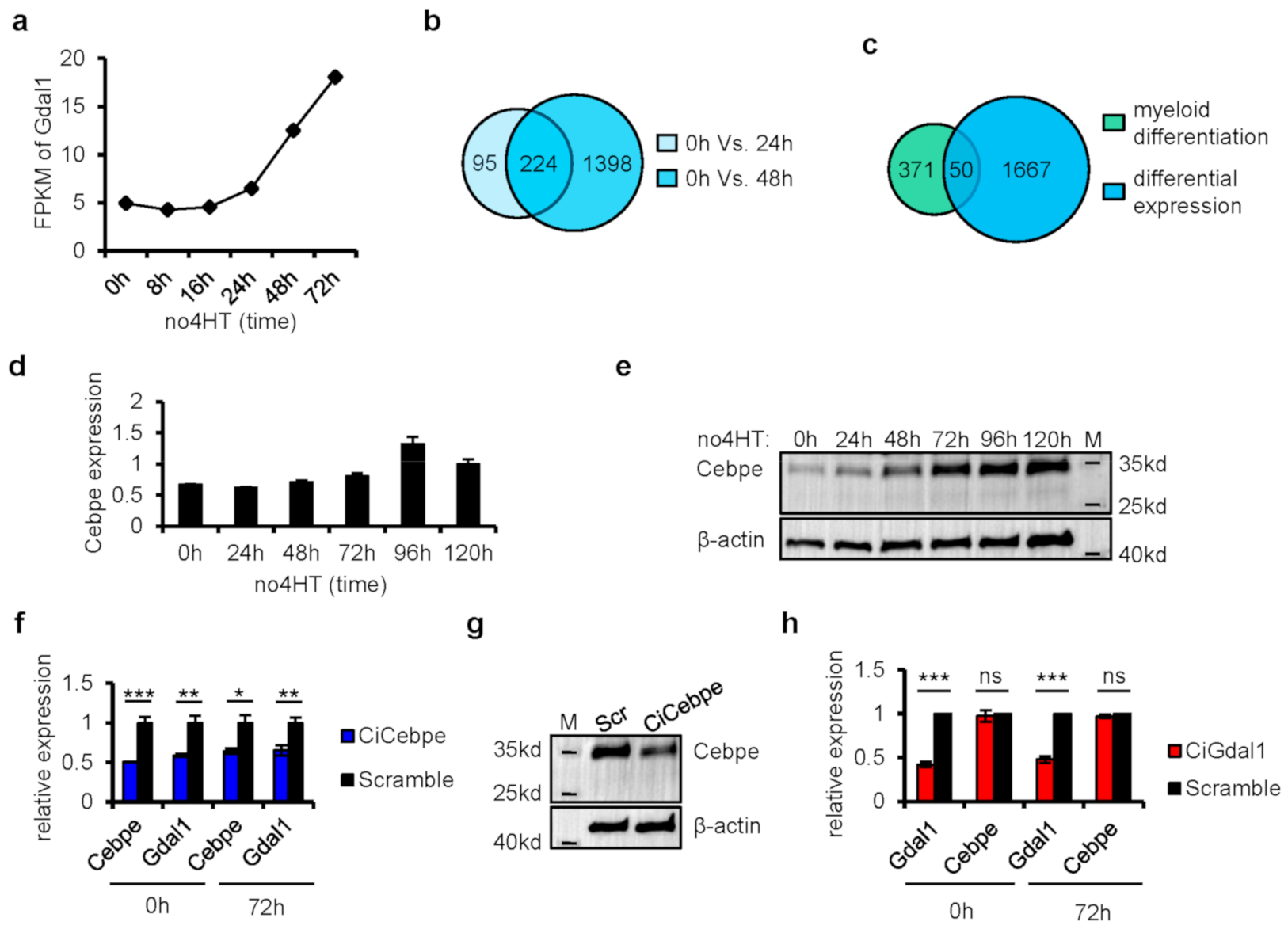
3.2. Gdal1 Was Required for Granulocytic Differentiation
3.3. Knockdown of Cebpe Led to Down-Regulation of Gdal1, but Not Vice Versa
3.4. Transcriptome Analysis in Gdal1 Knockdown CSH3 Cells Further Confirmed the Involvement of Gdal1 in Myeloid Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rieger, M.A.; Schroeder, T. Hematopoiesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008250. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Bresnick, E.H. Transcriptional control of erythropoiesis: Emerging mechanisms and principles. Oncogene 2007, 26, 6777–6794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nutt, S.L.; Kee, B.L. The transcriptional regulation of B cell lineage commitment. Immunity 2007, 26, 715–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbauer, F.; Tenen, D.G. Transcription factors in myeloid development: Balancing differentiation with transformation. Nat. Rev. Immunol. 2007, 7, 105–117. [Google Scholar] [CrossRef]
- Zhang, D.E.; Zhang, P.; Wang, N.D.; Hetherington, C.J.; Darlington, G.J.; Tenen, D.G. Absence of granulocyte colony-stimulating factor signaling and neutrophil development in CCAAT enhancer binding protein α-deficient mice. Proc. Natl. Acad. Sci. USA 1997, 94, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Iwasaki-Arai, J.; Iwasaki, H.; Fenyus, M.L.; Dayaram, T.; Owens, B.M.; Shigematsu, H.; Levantini, E.; Huettner, C.S.; Lekstrom-Himes, J.A.; et al. Enhancement of hematopoietic stem cell repopulating capacity and self-renewal in the absence of the transcription factor C/EBP α. Immunity 2004, 21, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, R.; Barlow, C.; Lekstrom-Himes, J.; Castilla, L.H.; Liu, P.P.; Eckhaus, M.; Decker, T.; Wynshaw-Boris, A.; Xanthopoulos, K.G. Impaired granulopoiesis, myelodysplasia, and early lethality in CCAAT/enhancer binding protein epsilon-deficient mice. Proc. Natl. Acad. Sci. USA 1997, 94, 13187–13192. [Google Scholar] [CrossRef] [Green Version]
- Halene, S.; Gaines, P.; Sun, H.; Zibello, T.; Lin, S.; Khanna-Gupta, A.; Williams, S.C.; Perkins, A.; Krause, D.; Berliner, N. C/EBPepsilon directs granulocytic-vs-monocytic lineage determination and confers chemotactic function via Hlx. Exp. Hematol. 2010, 38, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Tenen, D.G. Disruption of differentiation in human cancer: AML shows the way. Nat. Rev. Cancer 2003, 3, 89–101. [Google Scholar] [CrossRef]
- Yang, W.; Ernst, P. SET/MLL family proteins in hematopoiesis and leukemia. Int. J. Hematol. 2017, 105, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Zeisig, B.B.; Milne, T.; García-Cuéllar, M.P.; Schreiner, S.; Martin, M.E.; Fuchs, U.; Borkhardt, A.; Chanda, S.K.; Walker, J.; Soden, R.; et al. Hoxa9 and Meis1 are key targets for MLL-ENL-mediated cellular immortalization. Mol. Cell. Biol. 2004, 24, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Cuellar, M.P.; Büttner, C.; Bartenhagen, C.; Dugas, M.; Slany, R.K. Leukemogenic MLL-ENL Fusions Induce Alternative Chromatin States to Drive a Functionally Dichotomous Group of Target Genes. Cell Rep. 2016, 15, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Sun, Y.; Wang, J.; Jiang, H.; Muntean, A.G. Differential regulation of the c-Myc/Lin28 axis discriminates subclasses of rearranged MLL leukemia. Oncotarget 2016, 7, 25208–25223. [Google Scholar] [CrossRef] [Green Version]
- Ugale, A.; Norddahl, G.L.; Wahlestedt, M.; Säwén, P.; Jaako, P.; Pronk, C.J.; Soneji, S.; Cammenga, J.; Bryder, D. Hematopoietic stem cells are intrinsically protected against MLL-ENL-mediated transformation. Cell Rep. 2014, 9, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Jeong, M.; Sun, D.; Park, H.J.; Rodriguez, B.A.; Xia, Z.; Yang, L.; Zhang, X.; Sheng, K.; Darlington, G.J.; et al. Long non-coding RNAs control hematopoietic stem cell function. Cell Stem Cell 2015, 16, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Lian, Z.; Padden, C.; Gerstein, M.B.; Rozowsky, J.; Snyder, M.; Gingeras, T.R.; Kapranov, P.; Weissman, S.M.; Newburger, P.E. A myelopoiesis-associated regulatory intergenic noncoding RNA transcript within the human HOXA cluster. Blood 2009, 113, 2526–2534. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Weissman, S.M.; Newburger, P.E. Long intergenic non-coding RNA HOTAIRM1 regulates cell cycle progression during myeloid maturation in NB4 human promyelocytic leukemia cells. RNA Biol. 2014, 11, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.T.; Lin, H.S.; Shen, C.; Ma, Y.N.; Wang, F.; Zhao, H.L.; Yu, J.; Zhang, J.W. PU.1-Regulated Long Noncoding RNA lnc-MC Controls Human Monocyte/Macrophage Differentiation through Interaction with MicroRNA 199a-5p. Mol. Cell. Biol. 2015, 35, 3212–3224. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Xue, Y.; Han, Y.; Lin, L.; Wu, C.; Xu, S.; Jiang, Z.; Xu, J.; Liu, Q.; Cao, X. The STAT3-binding long noncoding RNA lnc-DC controls human dendritic cell differentiation. Science 2014, 344, 310–313. [Google Scholar] [CrossRef]
- Wagner, L.A.; Christensen, C.J.; Dunn, D.M.; Spangrude, G.J.; Georgelas, A.; Kelley, L.; Esplin, M.S.; Weiss, R.B.; Gleich, G.J. EGO, a novel, noncoding RNA gene, regulates eosinophil granule protein transcript expression. Blood 2007, 109, 5191–5198. [Google Scholar] [CrossRef]
- Delás, M.J.; Sabin, L.R.; Dolzhenko, E.; Knott, S.R.; Maravilla, E.M.; Jackson, B.T.; Wild, S.A.; Kovacevic, T.; Stork, E.M.; Zhou, M.; et al. lncRNA requirements for mouse acute myeloid leukemia and normal differentiation. eLife 2017, 6, e25607. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Kang, Q.; Zhu, X.; Chen, Q.; Wang, X.; Chen, Y.; Ouyang, J.; Zhang, L.; Tan, H.; Chen, R.; et al. A long noncoding RNA critically regulates Bcr-Abl-mediated cellular transformation by acting as a competitive endogenous RNA. Oncogene 2015, 34, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, W.; Cao, L.; Li, Z.; Wang, X. Long Non-Coding RNA CCAT1 Acts as a Competing Endogenous RNA to Regulate Cell Growth and Differentiation in Acute Myeloid Leukemia. Mol. Cells 2016, 39, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; Xu, Y.; Xu, L.; Yu, X.; Cheng, J.; Yang, L.; Chen, S.; Li, Y. Inhibition of long non-coding RNA NEAT1 impairs myeloid differentiation in acute promyelocytic leukemia cells. BMC Cancer 2014, 14, 693. [Google Scholar] [CrossRef] [Green Version]
- Frankish, A.; Diekhans, M.; Ferreira, A.M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 2019, 47, D766–D773. [Google Scholar] [CrossRef] [Green Version]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution-trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.H. Emerging properties of animal gene regulatory networks. Nature 2010, 468, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Qian, P.; He, X.C.; Paulson, A.; Li, Z.; Tao, F.; Perry, J.M.; Guo, F.; Zhao, M.; Zhi, L.; Venkatraman, A.; et al. The Dlk1-Gtl2 Locus Preserves LT-HSC Function by Inhibiting the PI3K-mTOR Pathway to Restrict Mitochondrial Metabolism. Cell Stem Cell 2016, 18, 214–228. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Shen, W.; Zhang, J.; Yang, B.; Liu, Y.N.; Qi, H.; Yu, X.; Lu, S.Y.; Chen, Y.; Xu, Y.Z.; et al. CRISPR interference-based specific and efficient gene inactivation in the brain. Nat. Neurosci. 2018, 21, 447–454. [Google Scholar] [CrossRef]
- Peng, L.; Zhang, Y.; Xin, H. lncRNA SNHG3 facilitates acute myeloid leukemia cell growth via the regulation of miR-758-3p/SRGN axis. J. Cell. Biochem. 2020, 121, 1023–1031. [Google Scholar] [CrossRef]
- Friedman, A.D. Transcriptional control of granulocyte and monocyte development. Oncogene 2007, 26, 6816–6828. [Google Scholar] [CrossRef] [Green Version]
- Lekstrom-Himes, J.A. The role of C/EBP(epsilon) in the terminal stages of granulocyte differentiation. Stem Cells 2001, 19, 125–133. [Google Scholar] [CrossRef]
- Avellino, R.; Delwel, R. Expression and regulation of C/EBPα in normal myelopoiesis and in malignant transformation. Blood 2017, 129, 2083–2091. [Google Scholar] [CrossRef]
- Friedman, A.D. C/EBPα in normal and malignant myelopoiesis. Int. J. Hematol. 2015, 101, 330–341. [Google Scholar] [CrossRef] [Green Version]
- Aoki, N.; Kimura, Y.; Kimura, S.; Nagato, T.; Azumi, M.; Kobayashi, H.; Sato, K.; Tateno, M. Expression and functional role of MDL-1 (CLEC5A) in mouse myeloid lineage cells. J. Leukoc. Biol. 2009, 85, 508–517. [Google Scholar] [CrossRef]
- Bakker, A.B.; Baker, E.; Sutherland, G.R.; Phillips, J.H.; Lanier, L.L. Myeloid DAP12-associating lectin (MDL)-1 is a cell surface receptor involved in the activation of myeloid cells. Proc. Natl. Acad. Sci. USA 1999, 96, 9792–9796. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.M.; Civin, C.I.; Rorth, P.; Friedman, A.D. A novel temporal expression pattern of three C/EBP family members in differentiating myelomonocytic cells. Blood 1992, 80, 1725–1735. [Google Scholar] [CrossRef]
- Bagger, F.O.; Kinalis, S.; Rapin, N. BloodSpot: A database of healthy and malignant haematopoiesis updated with purified and single cell mRNA sequencing profiles. Nucleic Acids Res. 2019, 47, D881–D885. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.E.; Radomska, H.S.; Wu, B.; Zhang, P.; Winnay, J.N.; Bajnok, L.; Wright, W.S.; Schaufele, F.; Tenen, D.G.; MacDougald, O.A. Phosphorylation of C/EBPalpha inhibits granulopoiesis. Mol. Cell. Biol. 2004, 24, 675–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ensemble ID | Gene Name | Location | Scaled Degree | Enriched In 1 | p-Value |
|---|---|---|---|---|---|
| ENSMUSG00000118061 | Rbfaos | chr18 | 0.771 | Diff. arrest | 8.45 × 10−13 |
| ENSMUSG00000104184 | Gm37818 | chr11 | 0.833 | Diff. arrest | 1.17 × 10−11 |
| ENSMUSG00000086425 | Gdal1 | chr2 | 0.754 | Normal Diff. | 1.73 × 10−8 |
| ENSMUSG00000084085 | Gm16140 | chr11 | 0.751 | Diff. arrest | 2.77 × 10−5 |
| ENSMUSG00000109799 | Gm45515 | chr7 | 0.821 | Diff. arrest | 4.23 × 10−5 |
| ENSMUSG00000027196 | Alkbh3os1 | chr2 | 0.834 | Diff. arrest | 6.86 × 10−5 |
| ENSMUSG00000087026 | A230103J11Rik | chr8 | 0.772 | Diff. arrest | 3.00 × 10−4 |
| ENSMUSG00000097057 | Gm17638 | chr15 | 0.844 | Diff. arrest | 5.28 × 10−4 |
| ENSMUSG00000107320 | Gm42549 | chr6 | 0.780 | Diff. arrest | 4.61 × 10−3 |
| ENSMUSG00000107480 | Gm44165 | chr7 | 0.757 | Normal Diff. | 8.90 × 10−3 |
| ENSMUSG00000113184 | Gm49654 | chr12 | 0.836 | Normal Diff. | 1.75 × 10−2 |
| ENSMUSG00000108402 | 9430064I24Rik | chr7 | 0.817 | Diff. arrest | 2.28 × 10−2 |
| ENSMUSG00000078308 | Gm47854 | chr9 | 0.905 | Normal Diff. | 3.85 × 10−2 |
| ENSMUSG00000097772 | 5430416N02Rik | chr5 | 0.960 | Diff. arrest | 3.85 × 10−2 |
| Ensemble ID | Gene Name | Correlation Coefficient | p-Value (t-Test) |
|---|---|---|---|
| ENSMUSG00000086425 | Gdal1 | 1.000 | 0.00 |
| ENSMUSG00000052435 | Cebpe | 0.991 | 1.12 × 10−4 |
| ENSMUSG00000020160 | Meis1 | −0.991 | 1.22 × 10−4 |
| ENSMUSG00000005583 | Mef2c | −0.964 | 1.96 × 10−3 |
| ENSMUSG00000044220 | Nkx2-3 | −0.962 | 2.13 × 10−3 |
| ENSMUSG00000076431 | Sox4 | −0.961 | 2.26 × 10−3 |
| ENSMUSG00000038236 | Hoxa7 | −0.953 | 3.29 × 10−3 |
| ENSMUSG00000055148 | Klf2 | 0.947 | 4.08 × 10−3 |
| ENSMUSG00000022508 | Bcl6 | 0.946 | 4.37 × 10−3 |
| ENSMUSG00000038227 | Hoxa9 | −0.941 | 5.17 × 10−3 |
| ENSMUSG00000034957 | Cebpa | 0.939 | 5.39 × 10−3 |
| ENSMUSG00000021025 | Nfkbia | 0.935 | 6.15 × 10−3 |
| ENSMUSG00000021356 | Irf4 | 0.933 | 6.62 × 10−3 |
| ENSMUSG00000038253 | Hoxa5 | −0.896 | 1.55 × 10−2 |
| ENSMUSG00000041515 | Irf8 | −0.875 | 2.25 × 10−2 |
| ENSMUSG00000020644 | Id2 | 0.814 | 4.87 × 10−2 |
| ENSMUSG00000037465 | Klf10 | 0.784 | 6.48 × 10−2 |
| ENSMUSG00000015053 | Gata2 | 0.645 | 1.66 × 10−1 |
| ENSMUSG00000052684 | Jun | 0.507 | 3.05 × 10−1 |
| ENSMUSG00000022528 | Hes1 | 0.293 | 5.73 × 10−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, Y.; Li, M.; Mi, S. Identification of Potential Key lncRNAs in the Context of Mouse Myeloid Differentiation by Systematic Transcriptomics Analysis. Genes 2021, 12, 630. https://doi.org/10.3390/genes12050630
Lan Y, Li M, Mi S. Identification of Potential Key lncRNAs in the Context of Mouse Myeloid Differentiation by Systematic Transcriptomics Analysis. Genes. 2021; 12(5):630. https://doi.org/10.3390/genes12050630
Chicago/Turabian StyleLan, Yongqing, Meng Li, and Shuangli Mi. 2021. "Identification of Potential Key lncRNAs in the Context of Mouse Myeloid Differentiation by Systematic Transcriptomics Analysis" Genes 12, no. 5: 630. https://doi.org/10.3390/genes12050630
APA StyleLan, Y., Li, M., & Mi, S. (2021). Identification of Potential Key lncRNAs in the Context of Mouse Myeloid Differentiation by Systematic Transcriptomics Analysis. Genes, 12(5), 630. https://doi.org/10.3390/genes12050630