
The Role of Peptide Hormones Discovered in the 21st Century in the Regulation of Adipose Tissue Functions

 ,
,  , , , , , , , ,
, , , , , , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Adropin
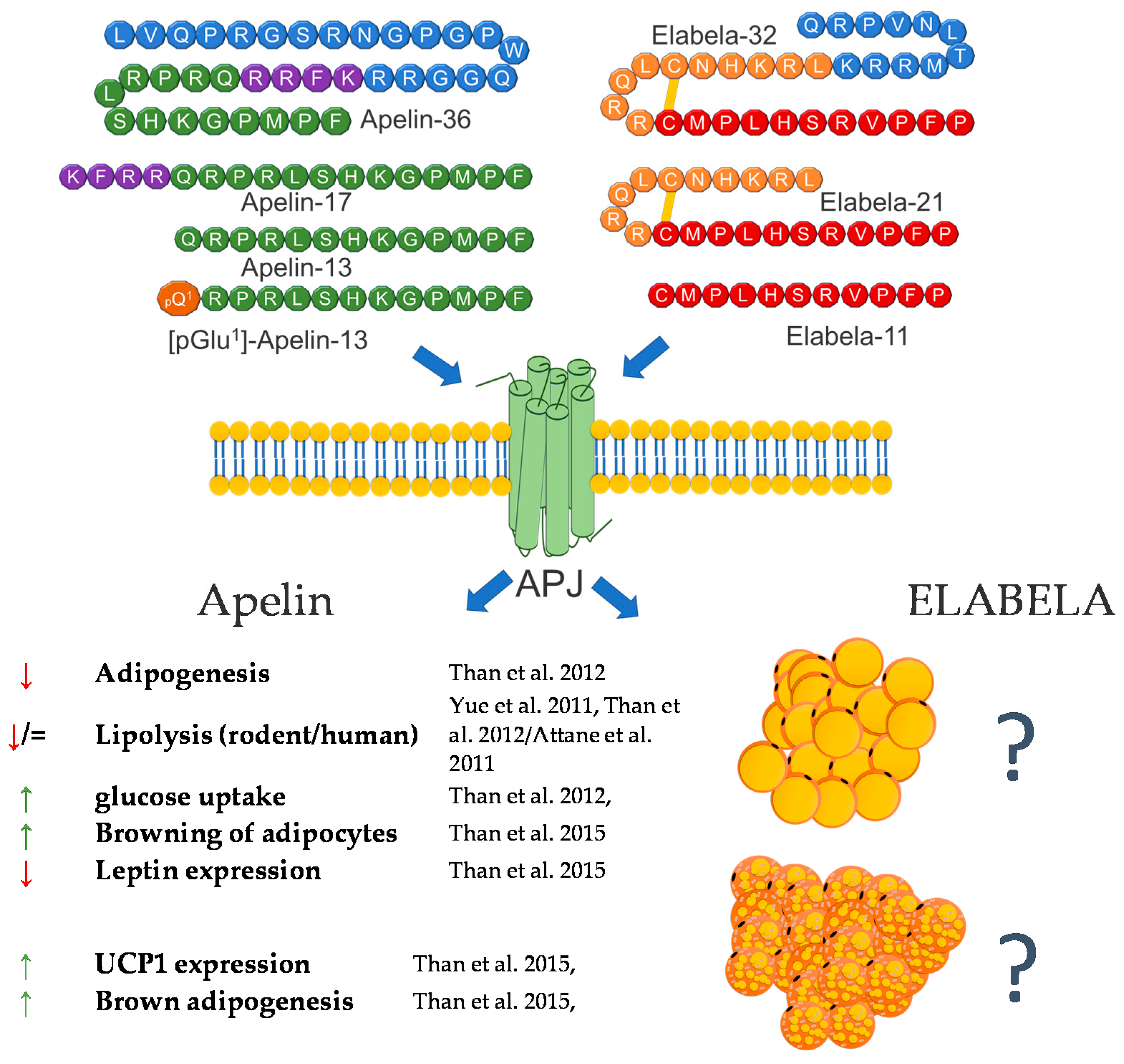
3. Apelin/Elabela
3.1. Apelin
3.2. Elabela/Toddler
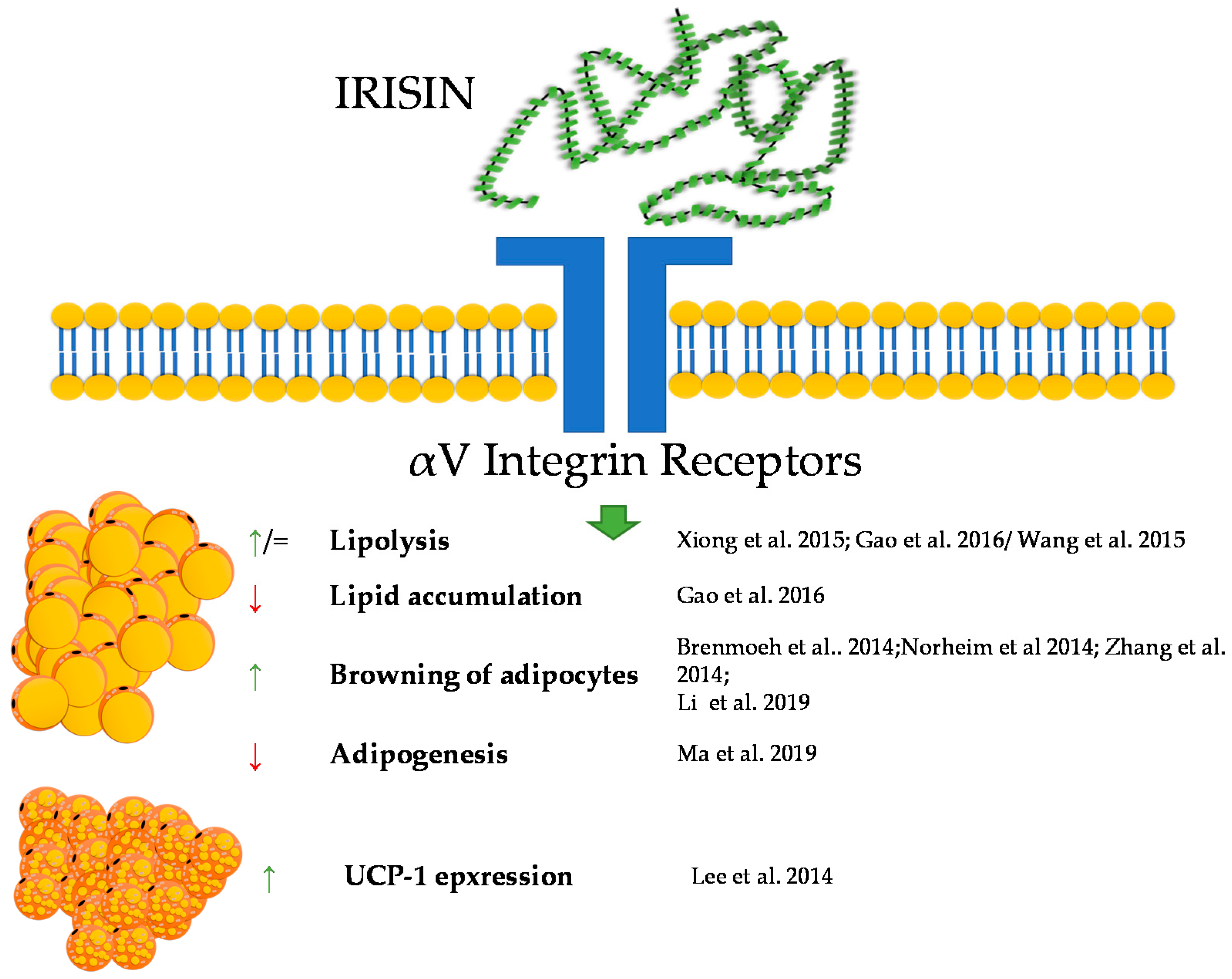
4. Irisin
5. Kisspeptin
6. Mots-C
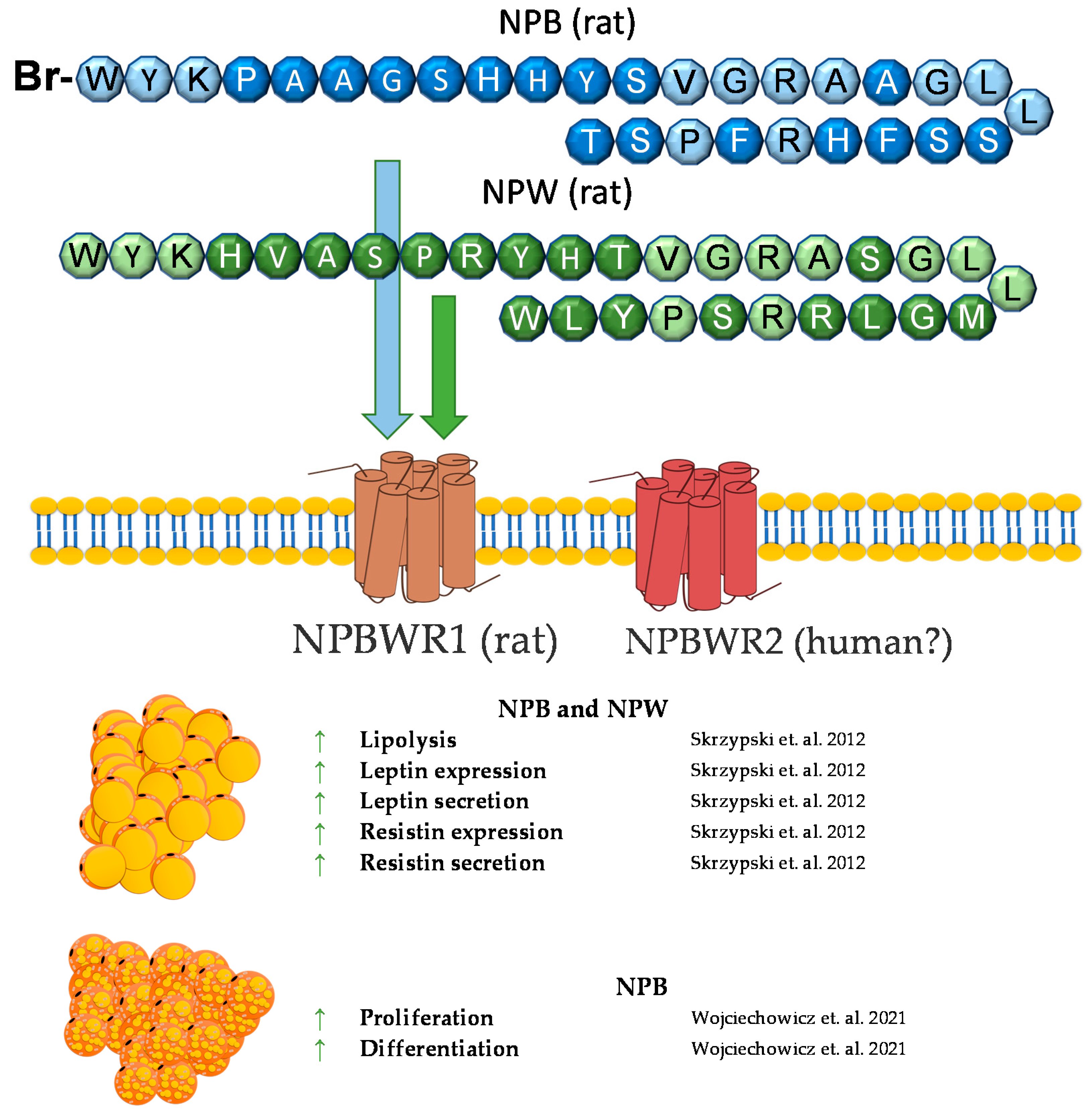
7. Neuropeptide B and Neuropeptide W
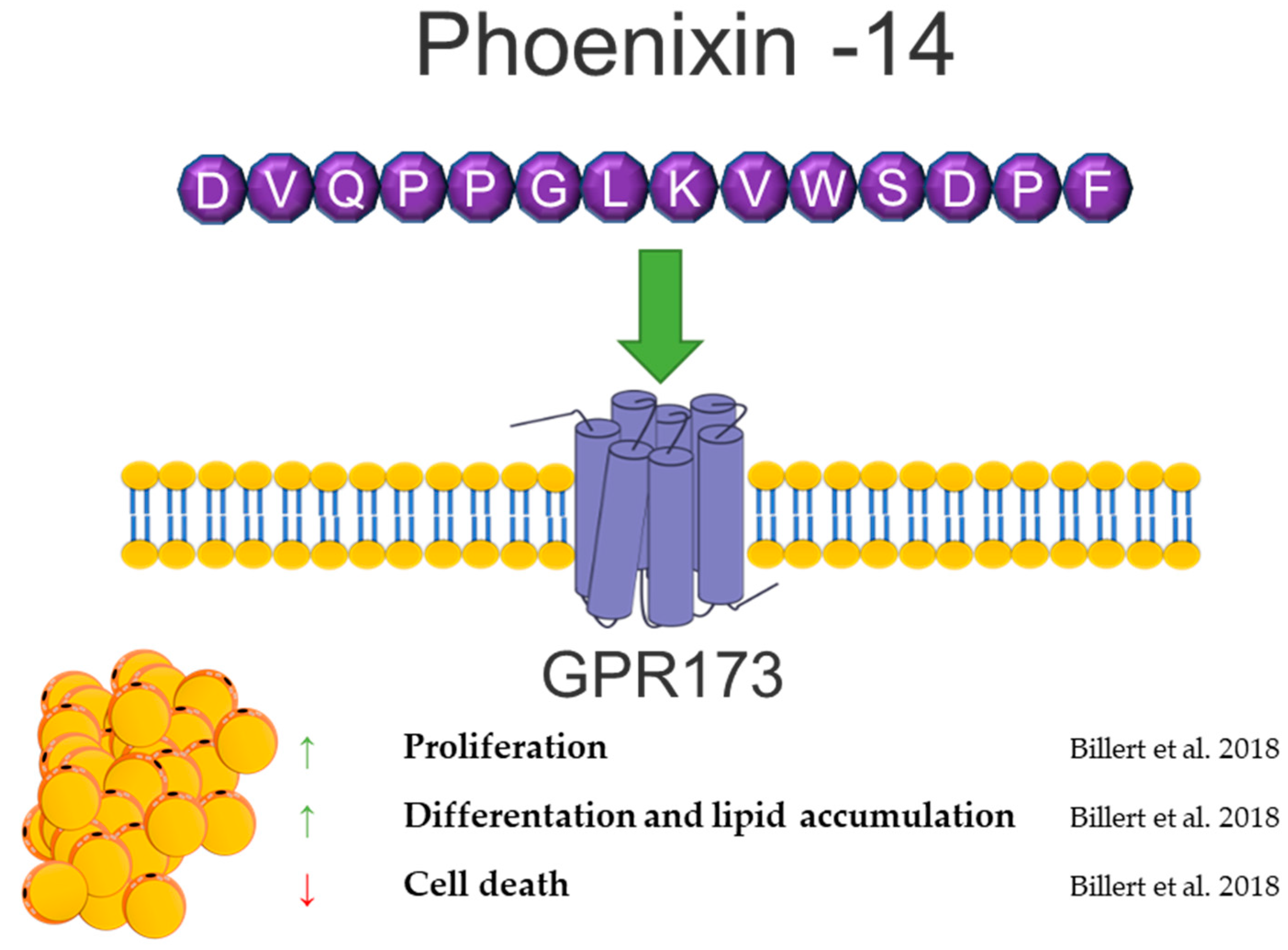
8. Phoenixin
9. Spexin
10. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Rocha, V.Z.; Libby, P. The Multiple Facets of the Fat Tissue. Thyroid 2008, 18, 175–183. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Obesity: Preventing and managing the global epidemic. In Report of a WHO Consultation; Technical Report Series; World Health Organization: Geneva, Switzerland, 2000; Volume 894, pp. 1–253. [Google Scholar]
- Tyagi, A.; Kapoor, P.; Kumar, R.; Chaudhary, K.; Gautam, A.; Raghava, G.P.S. In Silico Models for Designing and Discovering Novel Anticancer Peptides. Sci. Rep. 2013, 3, 2984. [Google Scholar] [CrossRef]
- Gao, S.; McMillan, R.P.; Zhu, Q.; Lopaschuk, G.D.; Hulver, M.W.; Butler, A.A. Therapeutic effects of adropin on glucose tolerance and substrate utilization in diet-induced obese mice with insulin resistance. Mol. Metab. 2015, 4, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.G.; Trevaskis, J.L.; Lam, D.D.; Sutton, G.M.; Koza, R.A.; Chouljenko, V.N.; Kousoulas, K.G.; Rogers, P.M.; Kesterson, R.A.; Thearle, M.; et al. Identification of Adropin as a Secreted Factor Linking Dietary Macronutrient Intake with Energy Homeostasis and Lipid Metabolism. Cell Metab. 2008, 8, 468–481. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Stevens, J.R.; Butler, A.A. Adropin—A circulating factor in metabolic control or a drop in the ocean? Expert Rev. Endocrinol. Metab. 2016, 11, 239–241. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, S.; Stevens, J.R.; Billon, C.; Girardet, C.; Sitaula, S.; Leon, A.S.; Rao, D.; Skinner, J.S.; Rankinen, T.; Bouchard, C.; et al. Adropin: An endocrine link between the biological clock and cholesterol homeostasis. Mol. Metab. 2018, 8, 51–64. [Google Scholar] [CrossRef]
- Lian, A.; Wu, K.; Liu, T.; Jiang, N.; Jiang, Q. Adropin induction of lipoprotein lipase expression in tilapia hepatocytes. J. Mol. Endocrinol. 2016, 56, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A.; Zhang, J.; Price, C.A.; Stevens, J.R.; Graham, J.L.; Stanhope, K.L.; King, S.; Krauss, R.M.; Bremer, A.A.; Havel, P.J. Low plasma adropin concentrations increase risks of weight gain and metabolic dysregulation in response to a high-sugar diet in male nonhuman primates. J. Biol. Chem. 2019, 294, 9706–9719. [Google Scholar] [CrossRef] [Green Version]
- Aydin, S.; Kuloglu, T.; Aydin, S.; Eren, M.N.; Yilmaz, M.; Kalayci, M.; Sahin, I.; Kocaman, N.; Çitil, C.; Kendir, Y. Expression of adropin in rat brain, cerebellum, kidneys, heart, liver, and pancreas in streptozotocin-induced diabetes. Mol. Cell. Biochem. 2013, 380, 73–81. [Google Scholar] [CrossRef]
- Choi, H.-N.; Yim, J.-E. Plasma Adropin as a Potential Marker Predicting Obesity and Obesity-associated Cancer in Korean Patients with Type 2 Diabetes Mellitus. J. Cancer Prev. 2018, 23, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.M.; Yosten, G.L.C.; Samson, W.K. Adropin acts in brain to inhibit water drinking: Potential interaction with the orphan G protein-coupled receptor, GPR19. Am. J. Physiol. Integr. Comp. Physiol. 2016, 310, R476–R480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.; Herr, D.R. G protein-coupled receptor GPR19 regulates E-cadherin expression and invasion of breast cancer cells. Biochim. Biophys. Acta Bioenerg. 2017, 1864, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- O’Dowd, B.F.; Nguyen, T.; Lynch, K.R.; Kolakowski, L.F.; Thompson, M.; Cheng, R.; Marchese, A.; Ng, G.; Heng, H.H.; George, S.R. A novel gene codes for a putative G protein-coupled receptor with an abundant expression in brain. FEBS Lett. 1996, 394, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-M.; Wang, Y.; Lee, J.T.H.; Huang, Z.; Wu, D.; Xu, A.; Lam, K.S.L. Adropin Is a Brain Membrane-bound Protein Regulating Physical Activity via the NB-3/Notch Signaling Pathway in Mice. J. Biol. Chem. 2014, 289, 25976–25986. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.A.; Tam, C.S.; Stanhope, K.L.; Wolfe, B.M.; Ali, M.R.; O’Keeffe, M.; St-Onge, M.-P.; Ravussin, E.; Havel, P.J. Low Circulating Adropin Concentrations with Obesity and Aging Correlate with Risk Factors for Metabolic Disease and Increase after Gastric Bypass Surgery in Humans. J. Clin. Endocrinol. Metab. 2012, 97, 3783–3791. [Google Scholar] [CrossRef] [Green Version]
- Sayın, O.; Tokgöz, Y.; Arslan, N. Investigation of adropin and leptin levels in pediatric obesity-related nonalcoholic fatty liver disease. J. Pediatr. Endocrinol. Metab. 2014, 27, 479–484. [Google Scholar] [CrossRef]
- Yu, H.-Y.; Zhao, P.; Wu, M.-C.; Liu, J.; Yin, W. Serum adropin levels are decreased in patients with acute myocardial infarction. Regul. Pept. 2014, 190–191, 46–49. [Google Scholar] [CrossRef]
- Zang, H.; Jiang, F.; Cheng, X.; Xu, H.; Hu, X. Serum adropin levels are decreased in Chinese type 2 diabetic patients and negatively correlated with body mass index. Endocr. J. 2018, 65, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Ganesh-Kumar, K.; Zhang, J.; Gao, S.; Rossi, J.; McGuinness, O.P.; Halem, H.H.; Culler, M.D.; Mynatt, R.L.; Butler, A.A. Adropin Deficiency Is Associated with Increased Adiposity and Insulin Resistance. Obesity 2012, 20, 1394–1402. [Google Scholar] [CrossRef] [Green Version]
- Jasaszwili, M.; Wojciechowicz, T.; Billert, M.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Effects of adropin on proliferation and differentiation of 3T3-L1 cells and rat primary preadipocytes. Mol. Cell. Endocrinol. 2019, 496, 110532. [Google Scholar] [CrossRef] [PubMed]
- Jasaszwili, M.; Wojciechowicz, T.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Adropin stimulates proliferation but suppresses differentiation in rat primary brown preadipocytes. Arch. Biochem. Biophys. 2020, 692, 108536. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.C.; Arneson, D.; Krishnan, K.C.; Ahn, I.S.; Diamante, G.; Cely, I.; Butte, A.J.; Lusis, A.J.; Yang, X.; Rajbhandari, P. Cold-associated mammokines preserve adipocyte identity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Nakagami, H. The Mechanism of White and Brown Adipocyte Differentiation. Diabetes Metab. J. 2013, 37, 85–90. [Google Scholar] [CrossRef] [Green Version]
- O’Dowd, B.F.; Heiber, M.; Chan, A.; Heng, H.H.; Tsui, L.-C.; Kennedy, J.L.; Shi, X.; Petronis, A.; George, S.R.; Nguyen, T. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene 1993, 136, 355–360. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y.; Wu, C.; Wan, Y.; Fang, C.; Li, J.; Fang, W.; Yi, R.; Zhu, G.; Li, J.; et al. Characterization of the Apelin/Elabela Receptors (APLNR) in Chickens, Turtles, and Zebrafish: Identification of a Novel Apelin-Specific Receptor in Teleosts. Front. Endocrinol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Pope, G.R.; Roberts, E.M.; Lolait, S.J.; O’Carroll, A.-M. Central and peripheral apelin receptor distribution in the mouse: Species differences with rat. Peptides 2012, 33, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, N.A.; Dupré, D.J.; Rainey, J.K. The apelin receptor: Physiology, pathology, cell signalling, and ligand modulation of a peptide-activated class A GPCR. Biochem. Cell Biol. 2014, 92, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Y.; Lou, Y.; Luo, M.; Lu, Y.; Li, Z.; Wang, Y.; Miao, L. Elabela, a newly discovered APJ ligand: Similarities and differences with Apelin. Peptides 2018, 109, 23–32. [Google Scholar] [CrossRef]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.-X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and Characterization of a Novel Endogenous Peptide Ligand for the Human APJ Receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Chng, S.C.; Ho, L.; Tian, J.; Reversade, B. ELABELA: A Hormone Essential for Heart Development Signals via the Apelin Receptor. Dev. Cell 2013, 27, 672–680. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.K.; Cheng, R.; Nguyen, T.; Fan, T.; Kariyawasam, A.P.; Liu, Y.; Osmond, D.H.; George, S.R.; O’Dowd, B.F. Characterization of Apelin, the Ligand for the APJ Receptor. J. Neurochem. 2001, 74, 34–41. [Google Scholar] [CrossRef]
- De Falco, M.; De Luca, L.; Onori, N.; Cavallotti, I.; Artigiano, F.; Esposito, V.; De Luca, B.; Laforgia, V.; Groeger, A.M.; De Luca, A. Apelin expression in normal human tissues. In Vivo 2002, 16, 333–336. [Google Scholar]
- Boucher, J.; Masri, B.; Daviaud, D.; Gesta, S.; Guigné, C.; Mazzucotelli, A.; Castan-Laurell, I.; Tack, I.; Knibiehler, B.; Carpéné, C.; et al. Apelin, a Newly Identified Adipokine Up-Regulated by Insulin and Obesity. Endocrinology 2005, 146, 1764–1771. [Google Scholar] [CrossRef]
- De Mota, N.; Lenkei, Z.; Llorens-Cortès, C. Cloning, Pharmacological Characterization and Brain Distribution of the Rat Apelin Receptor. Neuroendocrinology 2000, 72, 400–407. [Google Scholar] [CrossRef]
- Medhurst, A.D.; Jennings, C.A.; Robbins, M.J.; Davis, R.P.; Ellis, C.; Winborn, K.Y.; Lawrie, K.W.M.; Hervieu, G.; Riley, G.; Bolaky, J.E.; et al. Pharmacological and immunohistochemical characterization of the APJ receptor and its endogenous ligand apelin. J. Neurochem. 2003, 84, 1162–1172. [Google Scholar] [CrossRef]
- Tatemoto, K.; Takayama, K.; Zou, M.-X.; Kumaki, I.; Zhang, W.; Kumano, K.; Fujimiya, M. The novel peptide apelin lowers blood pressure via a nitric oxide-dependent mechanism. Regul. Pept. 2001, 99, 87–92. [Google Scholar] [CrossRef]
- Sunter, D.; Hewson, A.K.; Dickson, S.L. Intracerebroventricular injection of apelin-13 reduces food intake in the rat. Neurosci. Lett. 2003, 353, 1–4. [Google Scholar] [CrossRef]
- O’Shea, M.; Hansen, M.J.; Tatemoto, K.; Morris, M.J. Inhibitory Effect of Apelin-12 on Nocturnal Food Intake in the Rat. Nutr. Neurosci. 2003, 6, 163–167. [Google Scholar] [CrossRef]
- Ferrante, C.; Orlando, G.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di Nisio, C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; Vacca, M.; et al. Central apelin-13 administration modulates hypothalamic control of feeding. J. Boil. Regul. Homeost. Agents 2016, 30, 883–888. [Google Scholar]
- Valle, A.; Hoggard, N.; Adams, A.C.; Roca, P.; Speakman, J.R. Chronic central administration of apelin-13 over 10 days increases food intake, body weight, locomotor activity and body temperature in C57BL/6 mice. J. Neuroendocr. 2007, 20, 79–84. [Google Scholar] [CrossRef]
- Daviaud, D.; Boucher, J.; Gesta, S.; Dray, C.; Guigne, C.; Quilliot, D.; Ayav, A.; Ziegler, O.; Carpene, C.; Saulnier-Blache, J.-S.; et al. TNFα up-regulates apelin expression in human and mouse adipose tissue. FASEB J. 2006, 20, 1528–1530. [Google Scholar] [CrossRef]
- Wei, L.; Hou, X.; Tatemoto, K. Regulation of apelin mRNA expression by insulin and glucocorticoids in mouse 3T3-L1 adipocytes. Regul. Pept. 2005, 132, 27–32. [Google Scholar] [CrossRef]
- Geiger, K.; Muendlein, A.; Stark, N.; Saely, C.H.; Wabitsch, M.; Fraunberger, P.; Drexel, H. Hypoxia Induces Apelin Expression in Human Adipocytes. Horm. Metab. Res. 2011, 43, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Lorente-Cebrián, S.; Bustos, M.; Marti, A.; Martinez, J.A.; Moreno-Aliaga, M.J. Eicosapentaenoic acid up-regulates apelin secretion and gene expression in 3T3-L1 adipocytes. Mol. Nutr. Food Res. 2010, 54, S104–S111. [Google Scholar] [CrossRef]
- Han, S.; Wang, G.; Qi, X.; Englander, E.W.; Greeley, G.H. Involvement of a Stat3 binding site in inflammation-induced enteric apelin expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1068–G1078. [Google Scholar] [CrossRef]
- Mazzucotelli, A.; Ribet, C.; Castan-Laurell, I.; Daviaud, D.; Guigné, C.; Langin, D.; Valet, P. The transcriptional co-activator PGC-1α up regulates apelin in human and mouse adipocytes. Regul. Pept. 2008, 150, 33–37. [Google Scholar] [CrossRef]
- Hung, W.-W.; Hsieh, T.-J.; Lin, T.; Chou, P.-C.; Hsiao, P.-J.; Lin, K.-D.; Shin, S.-J. Blockade of the Renin-Angiotensin System Ameliorates Apelin Production in 3T3-L1 Adipocytes. Cardiovasc. Drugs Ther. 2010, 25, 3–12. [Google Scholar] [CrossRef]
- Yue, P.; Jin, H.; Xu, S.; Aillaud, M.; Deng, A.C.; Azuma, J.; Kundu, R.K.; Reaven, G.M.; Quertermous, T.; Tsao, P.S. Apelin Decreases Lipolysis via Gq, Gi, and AMPK-Dependent Mechanisms. Endocrinology 2011, 152, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Than, A.; Cheng, Y.; Foh, L.-C.; Leow, M.K.-S.; Lim, S.C.; Chuah, Y.J.; Kang, Y.; Chen, P. Apelin inhibits adipogenesis and lipolysis through distinct molecular pathways. Mol. Cell. Endocrinol. 2012, 362, 227–241. [Google Scholar] [CrossRef]
- Zhu, S.; Sun, F.; Li, W.; Cao, Y.; Wang, C.; Wang, Y.; Liang, N.; Zhang, R.; Zhang, S.; Wang, H.; et al. Apelin stimulates glucose uptake through the PI3K/Akt pathway and improves insulin resistance in 3T3-L1 adipocytes. Mol. Cell. Biochem. 2011, 353, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Attane, C.; Daviaud, D.; Dray, C.; Dusaulcy, R.; Masseboeuf, M.; Prévot, D.; Carpéné, C.; Castan-Laurell, I.; Valet, P. Apelin stimulates glucose uptake but not lipolysis in human adipose tissue ex vivo. J. Mol. Endocrinol. 2010, 46, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Than, A.; He, H.L.; Chua, S.H.; Xu, D.; Sun, L.; Leow, M.K.-S.; Chen, P. Apelin Enhances Brown Adipogenesis and Browning of White Adipocytes. J. Biol. Chem. 2015, 290, 14679–14691. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Miao, X.; Wang, S.; Liu, Y.; Sun, J.; Liu, Q.; Cai, L.; Wang, Y. Elabela may regulate SIRT3-mediated inhibition of oxidative stress through Foxo3a deacetylation preventing diabetic-induced myocardial injury. J. Cell. Mol. Med. 2021, 25, 323–332. [Google Scholar] [CrossRef]
- Onalan, E.; Doğan, Y.; Onalan, E.; Gozel, N.; Buran, I.; Donder, E. Elabela levels in patients with type 2 diabetes: Can it be a marker for diabetic nephropathy? Afr. Health Sci. 2020, 20, 833–840. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, D.; Ni, L.; Shi, L.; Xu, W.; Shi, M.; Chen, J.; Ai, Y.; Zhang, X. Serum Elabela/Toddler Levels Are Associated with Albuminuria in Patients with Type 2 Diabetes. Cell. Physiol. Biochem. 2018, 48, 1347–1354. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Panati, K.; Narala, V.R.; Narasimha, V.R.; Derangula, M.; Tatireddigari, V.R.A.; Yeguvapalli, S. Expression, purification and biological characterisation of recombinant human irisin (12.5 kDa). J. Genet. Eng. Biotechnol. 2018, 16, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Guerra, E.; Pardo, G.; Tinahones, F.; Ricart, W.; Fernández-Real, J.M. Irisin Is Expressed and Produced by Human Muscle and Adipose Tissue in Association with Obesity and Insulin Resistance. J. Clin. Endocrinol. Metab. 2013, 98, E769–E778. [Google Scholar] [CrossRef]
- Kim, H.; Wrann, C.D.; Jedrychowski, M.; Vidoni, S.; Kitase, Y.; Nagano, K.; Zhou, C.; Chou, J.; Parkman, V.-J.A.; Novick, S.J.; et al. Irisin Mediates Effects on Bone and Fat via αV Integrin Receptors. Cell 2018, 175, 1756–1768.e17. [Google Scholar] [CrossRef] [Green Version]
- Brenmoehl, J.; Albrecht, E.; Komolka, K.; Schering, L.; Langhammer, M.; Hoeflich, A.; Maak, S. Irisin Is Elevated in Skeletal Muscle and Serum of Mice Immediately after Acute Exercise. Int. J. Biol. Sci. 2014, 10, 338–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norheim, F.; Langleite, T.M.; Hjorth, M.; Holen, T.; Kielland, A.; Stadheim, H.K.; Gulseth, H.L.; Birkeland, K.I.; Jensen, J.; Drevon, C.A. The effects of acute and chronic exercise on PGC-1α, irisin and browning of subcutaneous adipose tissue in humans. FEBS J. 2013, 281, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Elbelt, U.; Stengel, A. Irisin as a muscle-derived hormone stimulating thermogenesis—A critical update. Peptides 2014, 54, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Y.; Wang, F.; Donelan, W.; Zona, M.C.; Li, S.; Reeves, W.; Ding, Y.; Tang, D.; Yang, L. Effects of irisin on the differentiation and browning of human visceral white adipocytes. Am. J. Transl. Res. 2019, 11, 7410–7421. [Google Scholar] [PubMed]
- Zhang, Y.; Li, R.; Meng, Y.; Li, S.; Donelan, W.; Zhao, Y.; Qi, L.; Zhang, M.; Wang, X.; Cui, T.; et al. Irisin Stimulates Browning of White Adipocytes through Mitogen-Activated Protein Kinase p38 MAP Kinase and ERK MAP Kinase Signaling. Diabetes 2014, 63, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xie, C.; Wang, H.; Foss, R.M.; Clare, M.; George, E.V.; Li, S.; Katz, A.; Cheng, H.; Ding, Y.; et al. Irisin exerts dual effects on browning and adipogenesis of human white adipocytes. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E530–E541. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Linderman, J.D.; Smith, S.; Brychta, R.J.; Wang, J.; Idelson, C.; Perron, R.M.; Werner, C.D.; Phan, G.Q.; Kammula, U.S.; et al. Irisin and FGF21 Are Cold-Induced Endocrine Activators of Brown Fat Function in Humans. Cell Metab. 2014, 19, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, J.Y.; Dincer, F.; Mesfum, E.; Mantzoros, C.S. Irisin stimulates muscle growth-related genes and regulates adipocyte differentiation and metabolism in humans. Int. J. Obes. 2014, 38, 1538–1544. [Google Scholar] [CrossRef]
- Vaughan, R.A.; Gannon, N.P.; Barberena, M.A.; Garcia-Smith, R.; Bisoffi, M.; Mermier, C.M.; Conn, C.A.; Trujillo, K.A. Characterization of the metabolic effects of irisin on skeletal muscle in vitro. Diabetes Obes. Metab. 2014, 16, 711–718. [Google Scholar] [CrossRef]
- Xiong, X.-Q.; Chen, D.; Sun, H.-J.; Ding, L.; Wang, J.-J.; Chen, Q.; Li, Y.-H.; Zhou, Y.-B.; Han, Y.; Zhang, F.; et al. FNDC5 overexpression and irisin ameliorate glucose/lipid metabolic derangements and enhance lipolysis in obesity. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1867–1875. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, L.; Li, W.; Yan, F.; Tian, M.; Wu, C.; Qi, L.; Wang, X.; Song, J.; Hou, X.; et al. Irisin has no effect on lipolysis in 3T3-L1 adipocytes or fatty acid metabolism in HepG2 hepatocytes. Endocrine 2014, 49, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.B.; Sahar, N.E.; Jeong, M.; Huh, J.Y. Irisin Exerts Inhibitory Effect on Adipogenesis through Regulation of Wnt Signaling. Front. Physiol. 2019, 10, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Li, F.; Li, H.; Huang, Y.; Liu, Y.; Chen, Y. Effects and Molecular Mechanism of GST-Irisin on Lipolysis and Autocrine Function in 3T3-L1 Adipocytes. PLoS ONE 2016, 11, e0147480. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Yu, R.; Liu, S.; Huwatibieke, B.; Li, Z.; Zhang, W. Irisin Inhibits Hepatic Cholesterol Synthesis via AMPK-SREBP2 Signaling. EBioMedicine 2016, 6, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Tekin, S.; Erden, Y.; Ozyalin, F.; Cigremis, Y.; Colak, C.; Sandal, S. The effects of intracerebroventricular infusion of irisin on feeding behaviour in rats. Neurosci. Lett. 2017, 645, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin improves fatty acid oxidation and glucose utilization in type 2 diabetes by regulating the AMPK signaling pathway. Int. J. Obes. 2016, 40, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Raschke, S.; Elsen, M.; Gassenhuber, H.; Sommerfeld, M.; Schwahn, U.; Brockmann, B.; Jung, R.; Wisløff, U.; Tjønna, A.E.; Raastad, T.; et al. Evidence against a Beneficial Effect of Irisin in Humans. PLoS ONE 2013, 8, e73680. [Google Scholar] [CrossRef] [Green Version]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.-M.; Le Poul, E.; Brézillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The Metastasis Suppressor Gene KiSS-1 Encodes Kisspeptins, the Natural Ligands of the Orphan G Protein-coupled Receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [Green Version]
- Michelle, L. Gottsch et al From KISS1 to Kisspeptins: An historical Perspective and Suggested Nomenclature. Peptides 2009, 30, 4–9. [Google Scholar] [CrossRef]
- Trevisan, C.M.; Montagna, E.; De Oliveira, R.; Christofolini, D.M.; Barbosa, C.P.; Crandall, K.A.; Bianco, B. Kisspeptin/GPR54 System: What Do We Know about Its Role in Human Reproduction? Cell. Physiol. Biochem. 2018, 49, 1259–1276. [Google Scholar] [CrossRef]
- Herbison, A.E.; De Tassigny, X.D.; Doran, J.; Colledge, W.H. Distribution and Postnatal Development of Gpr54 Gene Expression in Mouse Brain and Gonadotropin-Releasing Hormone Neurons. Endocrinology 2010, 151, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, A.S.; Park, J.H.; McPhie-Lalmansingh, A.A.; Gottsch, M.L.; Bodo, C.; Hohmann, J.G.; Pavlova, M.N.; Rohde, A.D.; Clifton, D.K.; Steiner, R.A.; et al. The Kisspeptin Receptor GPR54 Is Required for Sexual Differentiation of the Brain and Behavior. J. Neurosci. 2007, 27, 8826–8835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudek, M.; Kołodziejski, P.; Pruszyńska-Oszmałek, E.; Sassek, M.; Ziarniak, K.; Nowak, K.; Sliwowska, J. Effects of high-fat diet-induced obesity and diabetes on Kiss1 and GPR54 expression in the hypothalamic–pituitary–gonadal (HPG) axis and peripheral organs (fat, pancreas and liver) in male rats. Neuropeptides 2016, 56, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cui, X.; Xie, L.; Xing, R.; You, P.; Zhao, Y.; Yang, Y.; Xu, Y.; Zeng, L.; Chen, H.; et al. Kisspeptin Receptor GPR54 Promotes Adipocyte Differentiation and Fat Accumulation in Mice. Front. Physiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pruszyńska-Oszmałek, E.; Kołodziejski, P.A.; Sassek, M.; Sliwowska, J.H. Kisspeptin-10 inhibits proliferation and regulates lipolysis and lipogenesis processes in 3T3-L1 cells and isolated rat adipocytes. Endocrinology 2017, 56, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Song, W.J.; Mondal, P.; Wolfe, A.; Alonso, L.C.; Stamateris, R.; Ong, B.W.; Lim, O.C.; Yang, K.S.; Radovick, S.; Novaira, H.J.; et al. Glucagon regulates hepatic kisspeptin1 to impair insulin secretion. Cell Metab. 2014, 19, 667–681. [Google Scholar] [CrossRef] [Green Version]
- Fontana, R.; Della Torre, S. The Deep Correlation between Energy Metabolism and Reproduction: A View on the Effects of Nutrition for Women Fertility. Nutrients 2016, 8, 87. [Google Scholar] [CrossRef]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Molecules 2018, 23, 3071. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.; Imran, S.; Ur, E.; Wilkinson, M. KiSS-1 mRNA in adipose tissue is regulated by sex hormones and food intake. Mol. Cell. Endocrinol. 2008, 281, 64–72. [Google Scholar] [CrossRef]
- Wahab, F.; Riaz, T.; Shahab, M. Study on the Effect of Peripheral Kisspeptin Administration on Basal and Glucose-induced Insulin Secretion under Fed and Fasting Conditions in the Adult Male Rhesus Monkey (Macaca mulatta). Horm. Metab. Res. 2010, 43, 37–42. [Google Scholar] [CrossRef]
- Tolson, K.P.; Marooki, N.; De Bond, J.P.; Walenta, E.; Stephens, S.B.; Liaw, R.B.; Savur, R.; Wolfe, A.; Oh, D.Y.; Smith, J.T.; et al. Conditional knockout of kisspeptin signaling in brown adipose tissue increases metabolic rate and body temperature and lowers body weight. FASEB J. 2020, 34, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Halvorson, C.; De Bond, J.; Maloney, S.; Smith, J. Thermoneutral conditions correct the obese phenotype in female, but not male, Kiss1r knockout mice. J. Therm. Biol. 2020, 90, 102592. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.S.; Vu, J.P.; Oh, S.; Sanford, D.; Pisegna, J.R.; Germano, P. Intraperitoneal Treatment of Kisspeptin Suppresses Appetite and Energy Expenditure and Alters Gastrointestinal Hormones in Mice. Dig. Dis. Sci. 2019, 65, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Wang, L.; Goebel-Stengel, M.; Taché, Y. Centrally injected kisspeptin reduces food intake by increasing meal intervals in mice. NeuroReport 2011, 22, 253–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, Z.; Ozcan, M.; Ozkaya, A.; Canpolat, S.; Kutlu, S.; Kelestimur, H. Percentages of serum, liver and adipose tissue fatty acids and body weight are affected in female rats by long-term Central kisspeptin treatments. Arch. Physiol. Biochem. 2020, 1–9. [Google Scholar] [CrossRef]
- Zhai, L.; Zhao, J.; Zhu, Y.; Liu, Q.; Niu, W.; Liu, C.; Wang, Y. Downregulation of leptin receptor and kisspeptin/GPR54 in the murine hypothalamus contributes to male hypogonadism caused by high-fat diet-induced obesity. Endocrine 2018, 62, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, H.; Yang, S.; Li, Y.; Wang, B.; Chen, Y.; Wu, X. High-fat diet decreases the expression of Kiss1 mRNA and kisspeptin in the ovary, and increases ovulatory dysfunction in postpubertal female rats. Reprod. Biol. Endocrinol. 2014, 12, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kołodziejski, P.A.; Pruszyńska-Oszmałek, E.; Korek, E.; Sassek, M.; Szczepankiewicz, D.; Kaczmarek, P.; Nogowski, L.; Maćkowiak, P.; Nowak, K.W.; Krauss, H.; et al. Serum Levels of Spexin and Kisspeptin Negatively Correlate with Obesity and Insulin Resistance in Women. Physiol. Res. 2018, 67, 45–56. [Google Scholar] [CrossRef]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.-J.; Mehta, H.; Hevener, A.L.; De Cabo, R.; et al. The Mitochondrial-Derived Peptide MOTS-c Promotes Metabolic Homeostasis and Reduces Obesity and Insulin Resistance. Cell Metab. 2015, 21, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Mehta, H.H.; Wan, J.; Kuehnemann, C.; Chen, J.; Hu, J.-F.; Hoffman, A.R.; Cohen, P. Mitochondrial peptides modulate mitochondrial function during cellular senescence. Aging 2018, 10, 1239–1256. [Google Scholar] [CrossRef]
- Lu, H.; Tang, S.; Xue, C.; Liu, Y.; Wang, J.; Zhang, W.; Luo, W.; Chen, J. Mitochondrial-Derived Peptide MOTS-c Increases Adipose Thermogenic Activation to Promote Cold Adaptation. Int. J. Mol. Sci. 2019, 20, 2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Wei, M.; Zhai, Y.; Li, Q.; Ye, Z.; Wang, L.; Luo, W.; Chen, J.; Lu, Z. MOTS-c peptide regulates adipose homeostasis to prevent ovariectomy-induced metabolic dysfunction. J. Mol. Med. 2019, 97, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, H.; Hu, G.; Ye, Z.; Zhai, D.; Yan, Z.; Wang, L.; Xiang, A.; Lu, Z. Earlier changes in mice after D-galactose treatment were improved by mitochondria derived small peptide MOTS-c. Biochem. Biophys. Res. Commun. 2019, 513, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Ramanjaneya, M.; Jerobin, J.; Bettahi, I.; Bensila, M.; Aye, M.; Siveen, K.S.; Sathyapalan, T.; Skarulis, M.; Abou-Samra, A.; Atkin, S.L. Lipids and insulin regulate mitochondrial-derived peptide (MOTS-c) in PCOS and healthy subjects. Clin. Endocrinol. 2019, 91, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Fujii, R.; Yoshida, H.; Fukusumi, S.; Habata, Y.; Hosoya, M.; Kawamata, Y.; Yano, T.; Hinuma, S.; Kitada, C.; Asami, T.; et al. Identification of a Neuropeptide Modified with Bromine as an Endogenous Ligand for GPR7. J. Biol. Chem. 2002, 277, 34010–34016. [Google Scholar] [CrossRef] [Green Version]
- Brezillon, S.; Lannoy, V.; Franssen, J.-D.; Le Poul, E.; Dupriez, V.; Lucchetti, J.; Detheux, M.; Parmentier, M. Identification of Natural Ligands for the Orphan G Protein-coupled Receptors GPR7 and GPR8. J. Biol. Chem. 2003, 278, 776–783. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Yoshida, T.; Miyamoto, N.; Motoike, T.; Kurosu, H.; Shibata, K.; Yamanaka, A.; Williams, S.C.; Richardson, J.A.; Tsujino, N.; et al. Characterization of a family of endogenous neuropeptide ligands for the G protein-coupled receptors GPR7 and GPR8. Proc. Natl. Acad. Sci. USA 2003, 100, 6251–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, G.; Lin, D.; Cui, L.; Huang, L.; Lv, C.; Huang, S.; Wan, Y.; Fang, C.; Li, J.; Wang, Y. Characterization of Neuropeptide B (NPB), Neuropeptide W (NPW), and Their Receptors in Chickens: Evidence for NPW Being a Novel Inhibitor of Pituitary GH and Prolactin Secretion. Endocrinology 2016, 157, 3562–3576. [Google Scholar] [CrossRef]
- Takenoya, F.; Kageyama, H.; Shiba, K.; Date, Y.; Nakazato, M.; Shioda, S. Neuropeptide W: A key player in the homeostatic regulation of feeding and energy metabolism? Ann. N. Y. Acad. Sci. 2010, 1200, 162–169. [Google Scholar] [CrossRef]
- Shimomura, Y.; Harada, M.; Goto, M.; Sugo, T.; Matsumoto, Y.; Abe, M.; Watanabe, T.; Asami, T.; Kitada, C.; Mori, M.; et al. Identification of Neuropeptide W as the Endogenous Ligand for Orphan G-protein-coupled Receptors GPR7 and GPR8. J. Biol. Chem. 2002, 277, 35826–35832. [Google Scholar] [CrossRef] [Green Version]
- Takenoya, F.; Kageyama, H.; Hirako, S.; Ota, E.; Wada, N.; Ryushi, T.; Shioda, S. Neuropeptide W. Front. Endocrinol. 2012, 3, 171. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, Y.; Tanaka, H.; Motoike, T.; Ishii, M.; Williams, S.C.; Yanagisawa, M.; Sakurai, T. Distribution of neuropeptide W immunoreactivity and mRNA in adult rat brain. Brain Res. 2006, 1093, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Su, J.; Zheng, L.; Jin, M.; Hou, Y.; Ma, Z.; Guo, T.; Zhu, S.; Ma, X.; Ahmed, E.; et al. Cloning and distribution of neuropeptide W and its receptors in pigs. Res. Vet. Sci. 2015, 101, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Davenport, A.P. Neuropeptide B and W: Neurotransmitters in an emerging G-protein-coupled receptor system. Br. J. Pharmacol. 2006, 148, 1033–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.J.; Samson, W.K.; Ferguson, A.V. Neuropeptide W has cell phenotype-specific effects on the excitability of different subpopulations of paraventricular nucleus neurones. J. Neuroendocr. 2009, 21, 850–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzocchi, G.; Rebuffat, P.; Ziolkowska, A.; Rossi, G.P.; Malendowicz, L.K.; Nussdorfer, G.G. G Protein Receptors 7 and 8 Are Expressed in Human Adrenocortical Cells, and Their Endogenous Ligands Neuropeptides B and W Enhance Cortisol Secretion by Activating Adenylate Cyclase- and Phospholipase C-Dependent Signaling Cascades. J. Clin. Endocrinol. Metab. 2005, 90, 3466–3471. [Google Scholar] [CrossRef] [Green Version]
- Billert, M.; Sassek, M.; Wojciechowicz, T.; Jasaszwili, M.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Neuropeptide B stimulates insulin secretion and expression but not proliferation in rat insulin-producing INS-1E cells. Mol. Med. Rep. 2019, 20, 2030–2038. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowicz, T.; Billert, M.; Dhandapani, P.; Szczepankiewicz, D.; Wasielewski, O.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Neuropeptide B promotes proliferation and differentiation of rat brown primary preadipocytes. FEBS Open Bio 2021, 11, 1153–1164. [Google Scholar] [CrossRef]
- Dun, S.L.; Brailoiu, G.C.; Mizuo, K.; Yang, J.; Chang, J.K.; Dun, N.J. Neuropeptide B immunoreactivity in the central nervous system of the rat. Brain Res. 2005, 1045, 157–163. [Google Scholar] [CrossRef]
- Dvorakova, M.C. Distribution and Function of Neuropeptides W/B Signaling System. Front. Physiol. 2018, 9, 981. [Google Scholar] [CrossRef]
- Hochol, A.; Belloni, A.S.; Rucinski, M.; Ziolkowska, A.; Di Liddo, R.; Nussdorfer, G.G.; Malendowicz, L.K. Expression of neuropeptides B and W and their receptors in endocrine glands of the rat. Int. J. Mol. Med. 2006, 18, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Tuma, Z.; Peroni, E.; Monasson, O.; Papini, A.M.; Dvorakova, M.C. Identification of NPB, NPW and Their Receptor in the Rat Heart. Int. J. Mol. Sci. 2020, 21, 7827. [Google Scholar] [CrossRef]
- Kelly, M.A.; Beuckmann, C.T.; Williams, S.C.; Sinton, C.M.; Motoike, T.; Richardson, J.A.; Hammer, R.E.; Garry, M.G.; Yanagisawa, M. Neuropeptide B-deficient mice demonstrate hyperalgesia in response to inflammatory pain. Proc. Natl. Acad. Sci. USA 2005, 102, 9942–9947. [Google Scholar] [CrossRef] [Green Version]
- Samson, W.K.; Baker, J.R.; Samson, H.W.; Taylor, M.M. Central Neuropeptide B Administration Activates Stress Hormone Secretion and Stimulates Feeding in Male Rats. J. Neuroendocr. 2004, 16, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Hochol, A.; Albertin, G.; Nussdorfer, G.G.; Spinazzi, R.; Ziolkowska, A.; Rucinski, M.; Malendowicz, L.K. Effects of neuropeptides B and W on the secretion and growth of rat adrenocortical cells. Int. J. Mol. Med. 2004, 14, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ma, Z.; Suo, C.; Cheng, L.; Su, J.; Lei, Z.; Zhihai, L. Cloning and mRNA expression of NPB and its effect on hormone secretion of the reproductive cells in the pig. Gen. Comp. Endocrinol. 2018, 261, 97–103. [Google Scholar] [CrossRef]
- Dun, S.L.; Brailoiu, G.; Yang, J.; Chang, J.K.; Dun, N.J. Neuropeptide W-immunoreactivity in the hypothalamus and pituitary of the rat. Neurosci. Lett. 2003, 349, 71–74. [Google Scholar] [CrossRef]
- Takenoya, F.; Yagi, M.; Kageyama, H.; Shiba, K.; Endo, K.; Nonaka, N.; Date, Y.; Nakazato, M.; Shioda, S. Distribution of neuropeptide W in the rat brain. Neuropeptides 2010, 44, 99–106. [Google Scholar] [CrossRef]
- Skrzypski, M.; Pruszyńska-Oszmałek, E.; Rucinski, M.; Szczepankiewicz, D.; Sassek, M.; Wojciechowicz, T.; Kaczmarek, P.; Kołodziejski, P.A.; Strowski, M.Z.; Malendowicz, L.K.; et al. Neuropeptide B and W regulate leptin and resistin secretion, and stimulate lipolysis in isolated rat adipocytes. Regul. Pept. 2012, 176, 51–56. [Google Scholar] [CrossRef]
- Rucinski, M.; Nowak, K.W.; Chmielewska, J.; Ziolkowska, A.; Malendowicz, L.K. Neuropeptide W exerts a potent suppressive effect on blood leptin and insulin concentrations in the rat. Int. J. Mol. Med. 2007, 19, 401–405. [Google Scholar] [CrossRef]
- Mondal, M.S.; Yamaguchi, H.; Date, Y.; Shimbara, T.; Toshinai, K.; Shimomura, Y.; Mori, M.; Nakazato, M. A Role for Neuropeptide W in the Regulation of Feeding Behavior. Endocrinology 2003, 144, 4729–4733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Wada, M.; Irukayama-Tomobe, Y.; Ogata, Y.; Tsujino, N.; Suzuki, M.; Furutani, N.; Sakurai, T.; Yamamoto, M. A Single Nucleotide Polymorphism of the Neuropeptide B/W Receptor-1 Gene Influences the Evaluation of Facial Expressions. PLoS ONE 2012, 7, e35390. [Google Scholar] [CrossRef]
- Ishii, M.; Fei, H.; Friedman, J.M. Targeted disruption of GPR7, the endogenous receptor for neuropeptides B and W, leads to metabolic defects and adult-onset obesity. Proc. Natl. Acad. Sci. USA 2003, 100, 10540–10545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamaluddin, S.; Weakley, S.M.; Yao, Q.; Chen, C. Resistin: Functional roles and therapeutic considerations for cardiovascular disease. Br. J. Pharmacol. 2012, 165, 622–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriori, P.J.; Evans, A.E.; Sinnayah, P.; Cowley, M.A. Leptin Resistance and Obesity. Obesity 2006, 14, 254S–258S. [Google Scholar] [CrossRef]
- Yosten, G.L.C.; Lyu, R.-M.; Hsueh, A.J.; Avsian-Kretchmer, O.; Chang, J.-K.; Tullock, C.W.; Dun, S.L.; Dun, N.; Samson, W.K. A Novel Reproductive Peptide, Phoenixin. J. Neuroendocr. 2012, 25, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Cowan, A.; Lyu, R.-M.; Chen, Y.-H.; Dun, S.L.; Chang, J.-K.; Dun, N.J. Phoenixin: A candidate pruritogen in the mouse. Neuroscience 2015, 310, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Stein, L.M.; Haddock, C.J.; Samson, W.K.; Kolar, G.R.; Yosten, G.L. The phoenixins: From discovery of the hormone to identification of the receptor and potential physiologic actions. Peptides 2018, 106, 45–48. [Google Scholar] [CrossRef]
- McIlwraith, E.K.; Belsham, D.D. Phoenixin: Uncovering its receptor, signaling and functions. Acta Pharmacol. Sin. 2018, 39, 774–778. [Google Scholar] [CrossRef]
- Stein, L.M.; Tullock, C.W.; Mathews, S.K.; Garcia-Galiano, D.; Elias, C.F.; Samson, W.K.; Yosten, G.L.C. Hypothalamic action of phoenixin to control reproductive hormone secretion in females: Importance of the orphan G protein-coupled receptor Gpr173. Am. J. Physiol. Integr. Comp. Physiol. 2016, 311, R489–R496. [Google Scholar] [CrossRef] [Green Version]
- Yosten, G.L.C.; Kolar, G.R.; Redlinger, L.J.; Samson, W.K. Evidence for an interaction between proinsulin C-peptide and GPR146. J. Endocrinol. 2013, 218, B1–B8. [Google Scholar] [CrossRef] [PubMed]
- Treen, A.K.; Luo, V.; Belsham, D.D. Phoenixin Activates Immortalized GnRH and Kisspeptin Neurons through the Novel Receptor GPR173. Mol. Endocrinol. 2016, 30, 872–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, X.P.; Nakamura, T.; Osuka, S.; Bayasula, B.; Nakanishi, N.; Kasahara, Y.; Muraoka, A.; Hayashi, S.; Nagai, T.; Murase, T.; et al. Effect of the neuropeptide phoenixin and its receptor GPR173 during folliculogenesis. Reproduction 2019, 158, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ullah, K.; Rahman, T.U.; Wu, D.-D.; Lin, X.-H.; Liu, Y.; Guo, X.-Y.; Leung, P.C.; Zhang, R.-J.; Huang, H.-F.; Sheng, J.-Z. Phoenixin-14 concentrations are increased in association with luteinizing hormone and nesfatin-1 concentrations in women with polycystic ovary syndrome. Clin. Chim. Acta 2017, 471, 243–247. [Google Scholar] [CrossRef]
- Yuruyen, M.; Gultekin, G.; Batun, G.C.; Yavuzer, H.; Akcan, F.E.; Doventas, A.; Emul, M. Does plasma phoenixin level associate with cognition? Comparison between subjective memory complaint, mild cognitive impairment, and mild Alzheimer’s disease. Int. Psychogeriatr. 2017, 29, 1543–1550. [Google Scholar] [CrossRef]
- Billert, M.; Wojciechowicz, T.; Jasaszwili, M.; Szczepankiewicz, D.; Waśko, J.; Kaźmierczak, S.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Phoenixin-14 stimulates differentiation of 3T3-L1 preadipocytes via cAMP/Epac-dependent mechanism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1449–1457. [Google Scholar] [CrossRef]
- Kalamon, N.; Błaszczyk, K.; Szlaga, A.; Billert, M.; Skrzypski, M.; Pawlicki, P.; Górowska–Wójtowicz, E.; Kotula–Balak, M.; Błasiak, A.; Rak, A. Levels of the neuropeptide phoenixin-14 and its receptor GRP173 in the hypothalamus, ovary and periovarian adipose tissue in rat model of polycystic ovary syndrome. Biochem. Biophys. Res. Commun. 2020, 528, 628–635. [Google Scholar] [CrossRef]
- Delitala, A.P.; Capobianco, G.; Delitala, G.; Cherchi, P.L.; Dessole, S. Polycystic ovary syndrome, adipose tissue and metabolic syndrome. Arch. Gynecol. Obstet. 2017, 296, 405–419. [Google Scholar] [CrossRef]
- Mirabeau, O.; Perlas, E.; Severini, C.; Audero, E.; Gascuel, O.; Possenti, R.; Birney, E.; Rosenthal, N.; Gross, C. Identification of novel peptide hormones in the human proteome by hidden Markov model screening. Genome Res. 2007, 17, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-K.; Yun, S.; Son, G.H.; Hwang, J.-I.; Park, C.R.; Kim, J.I.; Kim, K.; Vaudry, H.; Seong, J.Y. Coevolution of the Spexin/Galanin/Kisspeptin Family: Spexin Activates Galanin Receptor Type II and III. Endocrinology 2014, 155, 1864–1873. [Google Scholar] [CrossRef] [Green Version]
- Porzionato, A.; Rucinski, M.; Macchi, V.; Stecco, C.; Malendowicz, L.K.; De Caro, R. Spexin Expression in Normal Rat Tissues. J. Histochem. Cytochem. 2010, 58, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kołodziejski, P.; Pruszyńska-Oszmałek, E.; Hejdysz, M.; Sassek, M.; Leciejewska, N.; Ziarniak, K.; Bień, J.; Ślósarz, P.; Kubiś, M.; Kaczmarek, S. Effects of Fasting on the Spexin System in Broiler Chickens. Animals 2021, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejski, P.A.; Pruszynska-Oszmalek, E.; Micker, M.; Skrzypski, M.; Wojciechowicz, T.; Szwarckopf, P.; Skieresz-Szewczyk, K.; Nowak, K.W.; Strowski, M.Z. Spexin: A novel regulator of adipogenesis and fat tissue metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1228–1236. [Google Scholar] [CrossRef]
- Zheng, B.; Li, S.; Liu, Y.; Li, Y.; Chen, H.; Tang, H.; Liu, X.; Lin, H.; Zhang, Y.; Cheng, C.H.K. Spexin Suppress Food Intake in Zebrafish: Evidence from Gene Knockout Study. Sci. Rep. 2017, 7, 14643. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Ma, Y.; Gu, M.; Zhang, Y.; Yan, S.; Li, N.; Wang, Y.; Ding, X.; Yin, J.; Fan, N.; et al. Spexin peptide is expressed in human endocrine and epithelial tissues and reduced after glucose load in type 2 diabetes. Peptides 2015, 71, 232–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.-Y.; Zhou, Y.-C.; Zhang, X.-M.; Chen, W.-D.; Wang, Y.-D. Emerging Roles of NPQ/Spexin in Physiology and Pathology. Front. Pharmacol. 2019, 10, 457. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.K.H.; Sze, K.H.; Chen, T.; Cho, C.K.; Law, H.C.H.; Chu, I.K.; Wong, A.O.L. Goldfish spexin: Solution structure and novel function as a satiety factor in feeding control. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E348–E366. [Google Scholar] [CrossRef] [Green Version]
- Walewski, J.L.; Ge, F.; Iv, H.L.; Levin, N.; Schwartz, G.J.; Vasselli, J.R.; Pomp, A.; Dakin, G.; Berk, P.D. Spexin is a novel human peptide that reduces adipocyte uptake of long chain fatty acids and causes weight loss in rodents with diet-induced obesity. Obesity 2014, 22, 1643–1652. [Google Scholar] [CrossRef] [Green Version]
- Karaca, A.; Bakar-Ates, F.; Ersoz-Gulcelik, N. Decreased Spexin Levels in Patients with Type 1 and Type 2 Diabetes. Med. Princ. Pract. 2018, 27, 549–554. [Google Scholar] [CrossRef]
- Behrooz, M.; Vaghef-Mehrabany, E.; Ostadrahimi, A. Different spexin level in obese vs normal weight children and its relationship with obesity related risk factors. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 674–682. [Google Scholar] [CrossRef]
- Behrooz, M.; Vaghef-Mehrabany, E.; Moludi, J.; Ostadrahimi, A. Are spexin levels associated with metabolic syndrome, dietary intakes and body composition in children? Diabetes Res. Clin. Pract. 2021, 172, 108634. [Google Scholar] [CrossRef] [PubMed]
- Pruszynska-Oszmalek, E.; Sassek, M.; Szczepankiewicz, D.; Nowak, K.W.; Kolodziejski, P.A. Short-term administration of spexin in rats reduces obesity by affecting lipolysis and lipogenesis: An in vivo and in vitro study. Gen. Comp. Endocrinol. 2020, 299, 113615. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.F.; Walewski, J.L.; Anglade, D.; Berk, P.D. Regulation of Hepatocellular Fatty Acid Uptake in Mouse Models of Fatty Liver Disease with and without Functional Leptin Signaling: Roles of NfKB and SREBP-1C and the Effects of Spexin. Semin. Liver Dis. 2016, 36, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Gambaro, S.E.; Zubiría, M.G.; Giordano, A.P.; Portales, A.E.; Alzamendi, A.; Rumbo, M.; Giovambattista, A. Spexin improves adipose tissue inflammation and macrophage recruitment in obese mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158700. [Google Scholar] [CrossRef] [PubMed]
- Said, M.A.; Nafeh, N.Y.; Abdallah, H.A. Spexin alleviates hypertension, hyperuricaemia, dyslipidemia and insulin resistance in high fructose diet induced metabolic syndrome in rats via enhancing PPAR-ɣ and AMPK and inhibiting IL-6 and TNF-α. Arch. Physiol. Biochem. 2021, 1–6. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kołodziejski, P.A.; Pruszyńska-Oszmałek, E.; Wojciechowicz, T.; Sassek, M.; Leciejewska, N.; Jasaszwili, M.; Billert, M.; Małek, E.; Szczepankiewicz, D.; Misiewicz-Mielnik, M.; et al. The Role of Peptide Hormones Discovered in the 21st Century in the Regulation of Adipose Tissue Functions. Genes 2021, 12, 756. https://doi.org/10.3390/genes12050756
Kołodziejski PA, Pruszyńska-Oszmałek E, Wojciechowicz T, Sassek M, Leciejewska N, Jasaszwili M, Billert M, Małek E, Szczepankiewicz D, Misiewicz-Mielnik M, et al. The Role of Peptide Hormones Discovered in the 21st Century in the Regulation of Adipose Tissue Functions. Genes. 2021; 12(5):756. https://doi.org/10.3390/genes12050756
Chicago/Turabian StyleKołodziejski, Paweł A., Ewa Pruszyńska-Oszmałek, Tatiana Wojciechowicz, Maciej Sassek, Natalia Leciejewska, Mariami Jasaszwili, Maria Billert, Emilian Małek, Dawid Szczepankiewicz, Magdalena Misiewicz-Mielnik, and et al. 2021. "The Role of Peptide Hormones Discovered in the 21st Century in the Regulation of Adipose Tissue Functions" Genes 12, no. 5: 756. https://doi.org/10.3390/genes12050756
APA StyleKołodziejski, P. A., Pruszyńska-Oszmałek, E., Wojciechowicz, T., Sassek, M., Leciejewska, N., Jasaszwili, M., Billert, M., Małek, E., Szczepankiewicz, D., Misiewicz-Mielnik, M., Hertig, I., Nogowski, L., Nowak, K. W., Strowski, M. Z., & Skrzypski, M. (2021). The Role of Peptide Hormones Discovered in the 21st Century in the Regulation of Adipose Tissue Functions. Genes, 12(5), 756. https://doi.org/10.3390/genes12050756