
Natural Antisense Transcript PEBP1P3 Regulates the RNA Expression, DNA Methylation and Histone Modification of CD45 Gene


Abstract
:1. Introduction
2. Materials and Methods
2.1. Database and Software
2.2. Cell Culture
2.3. RNAi Assay
2.4. RNA Extraction and RT-qPCR
2.5. Bisulfite Sequencing PCR for DNA Methylation Analysis
2.6. Chromatin Immunoprecipitation
3. Results
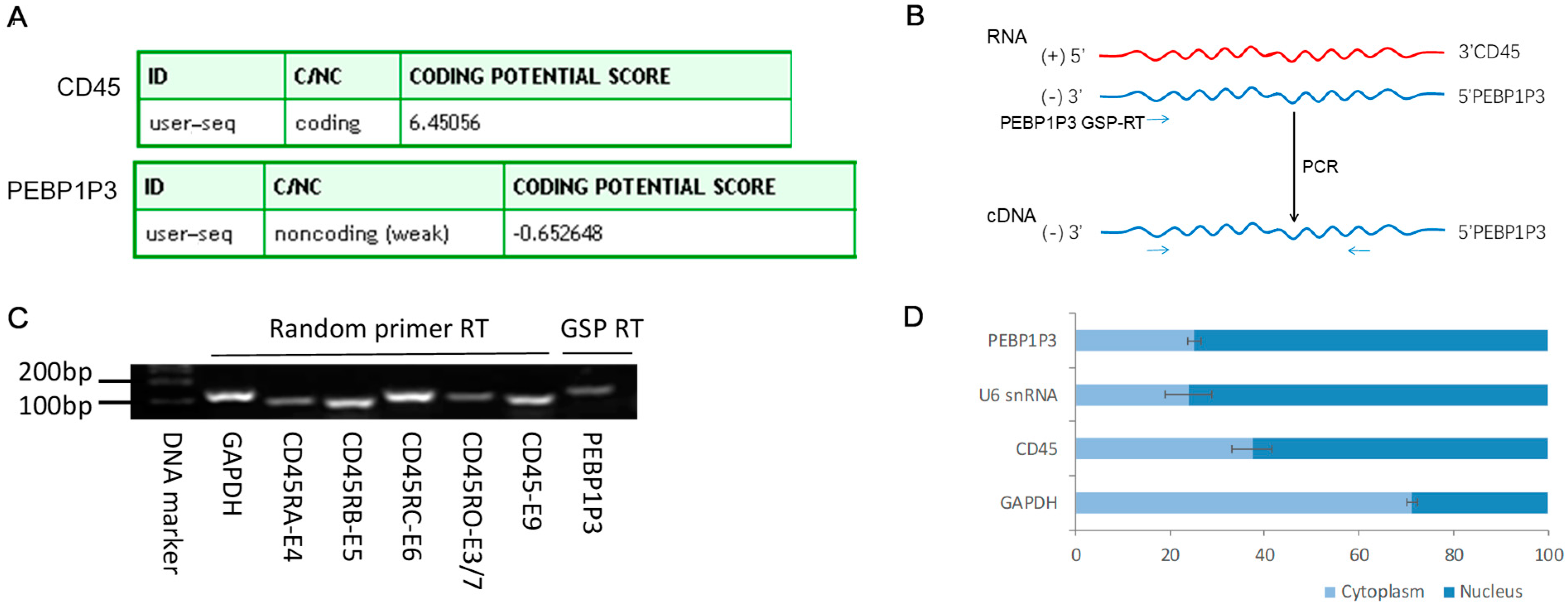
3.1. The Expression of CD45 RNA Was Positively Correlated with the Level of PEBP1P3 RNA in Multiple Human Tissues
3.2. The Expression and Subcellular Distribution of PEBP1P3 in Lymphocytes
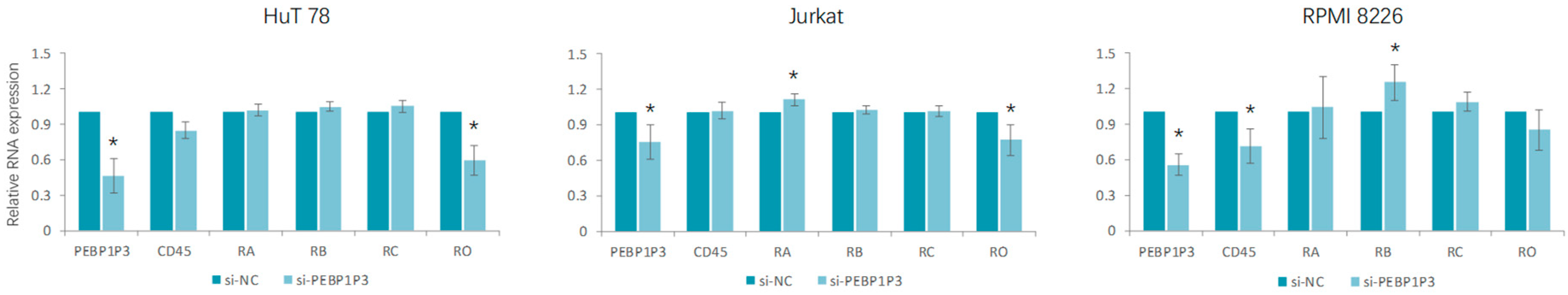
3.3. Knockdown of PEBP1P3 Altered the Expression of CD45 Isoforms
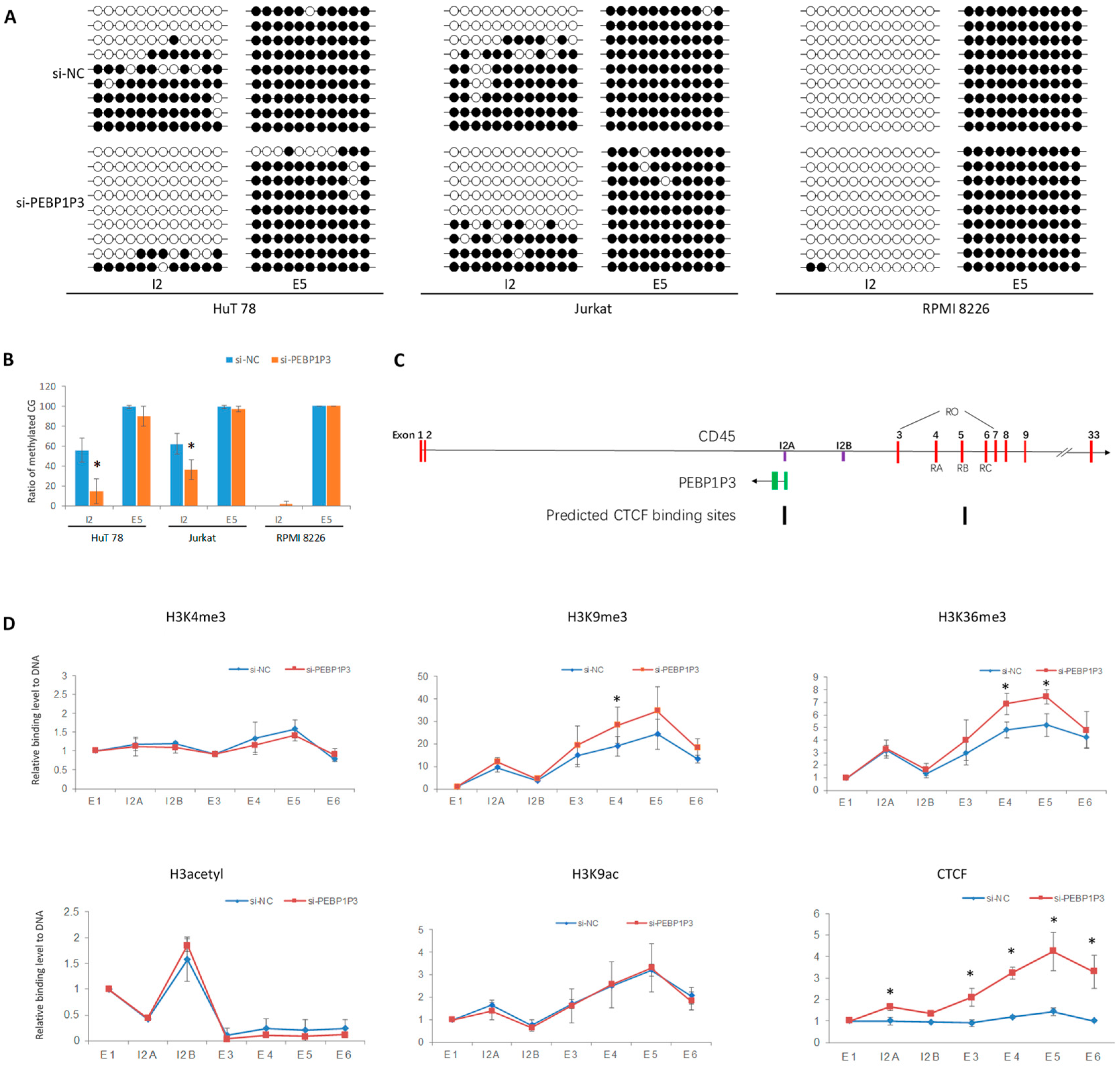
3.4. PEBP1P3 RNA Regulated DNA and Histone Modification of CD45
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A critical regulator of signaling thresholds in immune cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef]
- Chang, V.T.; Fernandes, R.A.; Ganzinger, K.A.; Lee, S.F.; Siebold, C.; McColl, J.; Jonsson, P.; Palayret, M.; Harlos, K.; Coles, C.H.; et al. Initiation of T cell signaling by CD45 segregation at ‘close contacts’. Nat. Immunol. 2016, 17, 574–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zikherman, J.; Weiss, A. Alternative splicing of CD45: The tip of the iceberg. Immunity 2008, 29, 839–841. [Google Scholar] [CrossRef] [Green Version]
- Courtney, A.H.; Shvets, A.A.; Lu, W.; Griffante, G.; Mollenauer, M.; Horkova, V.; Lo, W.L.; Yu, S.; Stepanek, O.; Chakraborty, A.K.; et al. CD45 functions as a signaling gatekeeper in T cells. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]
- Rheinlander, A.; Schraven, B.; Bommhardt, U. CD45 in human physiology and clinical medicine. Immunol. Lett. 2018, 196, 22–32. [Google Scholar] [CrossRef]
- Boxall, S.; Stanton, T.; Hirai, K.; Ward, V.; Yasui, T.; Tahara, H.; Tamori, A.; Nishiguchi, S.; Shiomi, S.; Ishiko, O.; et al. Disease associations and altered immune function in CD45 138G variant carriers. Hum. Mol. Genet. 2004, 13, 2377–2384. [Google Scholar] [CrossRef]
- Windhagen, A.; Sonmez, D.; Hornig-Do, H.T.; Kalinowsky, A.; Schwinzer, R. Altered CD45 isoform expression in C77G carriers influences cytokine responsiveness and adhesion properties of T cells. Clin. Exp. Immunol. 2007, 150, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Samaan, S.; Guerin-El Khourouj, V.; Auboeuf, D.; Peltier, L.; Pedron, B.; Ouachee-Chardin, M.; Gourgouillon, N.; Baruchel, A.; Dalle, J.H.; Sterkers, G. Outcome of children treated with haematopoietic-stem cell transplantations from donors expressing the rare C77G variant of the PTPRC (CD45) gene. Br. J. Haematol. 2011, 153, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Rothrock, C.R.; House, A.E.; Lynch, K.W. HnRNP L represses exon splicing via a regulated exonic splicing silencer. EMBO J. 2005, 24, 2792–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberdoerffer, S.; Moita, L.F.; Neems, D.; Freitas, R.P.; Hacohen, N.; Rao, A. Regulation of CD45 alternative splicing by heterogeneous ribonucleoprotein, hnRNPLL. Science 2008, 321, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Preussner, M.; Schreiner, S.; Hung, L.H.; Porstner, M.; Jack, H.M.; Benes, V.; Ratsch, G.; Bindereif, A. HnRNP L and L-like cooperate in multiple-exon regulation of CD45 alternative splicing. Nucleic Acids Res. 2012, 40, 5666–5678. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Oberdoerffer, S. Co-transcriptional regulation of alternative pre-mRNA splicing. Biochim. Biophys. Acta 2012, 1819, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marina, R.J.; Sturgill, D.; Bailly, M.A.; Thenoz, M.; Varma, G.; Prigge, M.F.; Nanan, K.K.; Shukla, S.; Haque, N.; Oberdoerffer, S. TET-catalyzed oxidation of intragenic 5-methylcytosine regulates CTCF-dependent alternative splicing. EMBO J. 2016, 35, 335–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldi, T.; Cortazar, M.A.; Sheridan, R.M.; Bentley, D.L. Coupling of RNA Polymerase II Transcription Elongation with Pre-mRNA Splicing. J. Mol. Biol. 2016, 428, 2623–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, J.; Yin, J.; Su, Z. Natural antisense RNAs are involved in the regulation of CD45 expression in autoimmune diseases. Lupus 2015, 24, 235–239. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.T. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar]
- Bao, L.; Zhou, M.; Cui, Y. CTCFBSDB: A CTCF-binding site database for characterization of vertebrate genomic insulators. Nucleic Acids Res. 2008, 36, 83–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebarth, J.D.; Bhattacharya, A.; Cui, Y. CTCFBSDB 2.0: A database for CTCF-binding sites and genome organization. Nucleic Acids Res. 2013, 41, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Faghihi, M.A.; Wahlestedt, C. Regulatory roles of natural antisense transcripts. Nat. Rev. Mol. Cell Biol. 2009, 10, 637–643. [Google Scholar] [CrossRef]
- Khorkova, O.; Myers, A.J.; Hsiao, J.; Wahlestedt, C. Natural antisense transcripts. Hum. Mol. Genet 2014, 23, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Farnebo, M.; Bykov, V.J.; Wiman, K.G. The p53 tumor suppressor: A master regulator of diverse cellular processes and therapeutic target in cancer. Biochem. Biophys. Res. Commun. 2010, 396, 85–89. [Google Scholar] [CrossRef]
- Qu, X.; Alsager, S.; Zhuo, Y.; Shan, B. HOX transcript antisense RNA (HOTAIR) in cancer. Cancer Lett. 2019, 454, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Acuna, L.G.; Barros, M.J.; Nunez, P.; Penaloza, D.; Montt, F.; Pedraza, D.; Crossley, K.; Gil, F.; Fuentes, J.A.; Calderon, I.L. The cis-encoded antisense RNA IsrA from Salmonella Typhimurium represses the expression of STM0294.1n (iasE), an SOS-induced gene coding for an endoribonuclease activity. Biochem. Biophys. Res. Commun. 2020, 526, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Luco, R.F.; Allo, M.; Schor, I.E.; Kornblihtt, A.R.; Misteli, T. Epigenetics in alternative pre-mRNA splicing. Cell 2011, 144, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothrock, C.; Cannon, B.; Hahm, B.; Lynch, K.W. A conserved signal-responsive sequence mediates activation-induced alternative splicing of CD45. Mol. Cell 2003, 12, 1317–1324. [Google Scholar] [CrossRef]
- Tong, A.; Nguyen, J.; Lynch, K.W. Differential expression of CD45 isoforms is controlled by the combined activity of basal and inducible splicing-regulatory elements in each of the variable exons. J. Biol. Chem. 2005, 280, 38297–38304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zhao, W.; Olson, S.D.; Prabhakara, K.S.; Zhou, X. Alternative splicing links histone modifications to stem cell fate decision. Genome Biol. 2018, 19, 133. [Google Scholar] [CrossRef] [Green Version]
- Rahhal, R.; Seto, E. Emerging roles of histone modifications and HDACs in RNA splicing. Nucleic Acids Res. 2019, 47, 4911–4926. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Luo, H.; Lee, S.; Jin, F.; Yang, J.S.; Montellier, E.; Buchou, T.; Cheng, Z.; Rousseaux, S.; Rajagopal, N.; et al. Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell 2011, 146, 1016–1028. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.L.; Pan, Q.; Tominaga, K.; Blencowe, B.J.; Pereira-Smith, O.M.; Misteli, T. Regulation of alternative splicing by local histone modifications: Potential roles for RNA-guided mechanisms. Nucleic Acids Res. 2014, 42, 701–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorenson, M.R.; Jha, D.K.; Ucles, S.A.; Flood, D.M.; Strahl, B.D.; Stevens, S.W.; Kress, T.L. Histone H3K36 methylation regulates pre-mRNA splicing in Saccharomyces cerevisiae. RNA Biol. 2016, 13, 412–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davie, J.R.; Xu, W.; Delcuve, G.P. Histone H3K4 trimethylation: Dynamic interplay with pre-mRNA splicing. Biochem. Cell Biol. 2016, 94, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Li, N.; Fu, D.; Ren, J.; Hui, J.; Peng, J.; Liu, Y.; Qiu, T.; Jiang, M.; Pan, Q.; et al. Histone methyltransferase SETD2 modulates alternative splicing to inhibit intestinal tumorigenesis. J. Clin. Investig. 2017, 127, 3375–3391. [Google Scholar] [CrossRef]
- Pradeepa, M.M.; Sutherland, H.G.; Ule, J.; Grimes, G.R.; Bickmore, W.A. Psip1/Ledgf p52 binds methylated histone H3K36 and splicing factors and contributes to the regulation of alternative splicing. PLoS Genet. 2012, 8, e1002717. [Google Scholar] [CrossRef] [Green Version]
- Vakoc, C.R.; Mandat, S.A.; Olenchock, B.A.; Blobel, G.A. Histone H3 lysine 9 methylation and HP1gamma are associated with transcription elongation through mammalian chromatin. Mol. Cell 2005, 19, 381–391. [Google Scholar] [CrossRef]
- Saint-Andre, V.; Batsche, E.; Rachez, C.; Muchardt, C. Histone H3 lysine 9 trimethylation and HP1gamma favor inclusion of alternative exons. Nat. Struct. Mol. Biol. 2011, 18, 337–344. [Google Scholar] [CrossRef]
- Bieberstein, N.I.; Kozakova, E.; Huranova, M.; Thakur, P.K.; Krchnakova, Z.; Krausova, M.; Carrillo Oesterreich, F.; Stanek, D. TALE-directed local modulation of H3K9 methylation shapes exon recognition. Sci. Rep. 2016, 6, 29961. [Google Scholar] [CrossRef] [Green Version]
- Neems, D.S.; Garza-Gongora, A.G.; Smith, E.D.; Kosak, S.T. Topologically associated domains enriched for lineage-specific genes reveal expression-dependent nuclear topologies during myogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 1691–1700. [Google Scholar] [CrossRef] [Green Version]
- Gorkin, D.U.; Qiu, Y.; Hu, M.; Fletez-Brant, K.; Liu, T.; Schmitt, A.D.; Noor, A.; Chiou, J.; Gaulton, K.J.; Sebat, J.; et al. Common DNA sequence variation influences 3-dimensional conformation of the human genome. Genome Biol. 2019, 20, 255. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.; Kern, F.; Schulz, M.H. Integrative prediction of gene expression with chromatin accessibility and conformation data. Epigenetics Chromatin 2020, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Velasco, M.; Kumar, M.; Lai, M.; Bhat, P.; Solis-Pinson, A.B.; Reyes, A.; Kleinsorg, S.; Noh, K.M.; Gibson, T.J.; Zaugg, J.B. CTCF-Mediated Chromatin Loops between Promoter and Gene Body Regulate Alternative Splicing across Individuals. Cell Syst. 2017, 5, 628–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Haarhuis, J.H.I.; Sedeno Cacciatore, A.; Oldenkamp, R.; van Ruiten, M.S.; Willems, L.; Teunissen, H.; Muir, K.W.; de Wit, E.; Rowland, B.D.; et al. The structural basis for cohesin-CTCF-anchored loops. Nature 2020, 578, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Wang, D.; Horton, J.R.; Zhang, X.; Corces, V.G.; Cheng, X. Structural Basis for the Versatile and Methylation-Dependent Binding of CTCF to DNA. Mol. Cell 2017, 66, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiehle, L.; Thorn, G.J.; Raddatz, G.; Clarkson, C.T.; Rippe, K.; Lyko, F.; Breiling, A.; Teif, V.B. DNA (de)methylation in embryonic stem cells controls CTCF-dependent chromatin boundaries. Genome Res. 2019, 29, 750–761. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Usage |
|---|---|---|
| CD45 E1-F | AGTAAAACCGAATCTGACATCATCACC | CD45 E1 DNA |
| CD45 E1-R | TGTTGTCTTATCAGACGAGGAACAATT | |
| CD45-I2A-F | TGCTTTGCTCTATTACTGTGGGC | CD45 I2A DNA |
| CD45-I2A-R | ATATTCTGGTACTGTGATGGGGTCA | |
| CD45-I2B-F | GCTATGGTGCCACCTACTGAAA | CD45 I2B DNA |
| CD45-I2B-R | ACAGTTACTACATTCTACACTTTGACT | |
| CD45 E3-F | TTGCCACTTGGTGAATGTTCTATC | CD45 E3 DNA |
| CD45 E3-R | GGAAGGTGTTGGGCTTTGC | |
| CD45 E4-F | GCAAAGATGCCCAGTGTTCCACTT | CD45 RA/E4 DNA |
| CD45 E4-R | TTCTCTTTCAAAGGTGCTTGCGG | |
| CD45 E5-F | TCATCAGTACAGACGCCTCACCTT | CD4 RB/E5 DNA |
| CD45 E5-R | CTGAATGTCTGCGTGTCAGTTCCA | |
| CD45 E6-F | AGCACCTTTCCTACAGACCCAGTT | CD45 RC/E6 DNA |
| CD45 E6-R | TGTTCGCTGTGATGGTGGTGTT | |
| CD45 E3/7-F | ACCTTCCCCCACTGATGCCTA | CD45 RO |
| CD45 E7-R | GTGGTTGAAATGACAGCGCTTC | |
| CD45 E9-F | GCACAAACAATGAGGTGCATAACC | CD45 RNA |
| CD45 E9-R | ATGTCTTATCAGGAGCAGTACATGA | |
| PEBP1P3-F | ATTCTGGTACTGTGATGGGGTCA | PEBP1P3 RNA |
| PEBP1P3-R | GATGCTTTGCTCTATTACTGTGGG | |
| GAPDH-F | GCACCGTCAAGGCTGAGAAC | GAPDH RNA |
| GAPDH-R | TGGTGAAGACGCCAGTGGA | |
| U6-F | GCTTCGGCAGCACATATACTAAAAT | U6 snRNA |
| U6-R | CGCTTCACGAATTTGCGTGTCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Z.; Liu, G.; Zhang, B.; Lin, Z.; Huang, D. Natural Antisense Transcript PEBP1P3 Regulates the RNA Expression, DNA Methylation and Histone Modification of CD45 Gene. Genes 2021, 12, 759. https://doi.org/10.3390/genes12050759
Su Z, Liu G, Zhang B, Lin Z, Huang D. Natural Antisense Transcript PEBP1P3 Regulates the RNA Expression, DNA Methylation and Histone Modification of CD45 Gene. Genes. 2021; 12(5):759. https://doi.org/10.3390/genes12050759
Chicago/Turabian StyleSu, Zhongjing, Guangyu Liu, Bin Zhang, Ze Lin, and Dongyang Huang. 2021. "Natural Antisense Transcript PEBP1P3 Regulates the RNA Expression, DNA Methylation and Histone Modification of CD45 Gene" Genes 12, no. 5: 759. https://doi.org/10.3390/genes12050759
APA StyleSu, Z., Liu, G., Zhang, B., Lin, Z., & Huang, D. (2021). Natural Antisense Transcript PEBP1P3 Regulates the RNA Expression, DNA Methylation and Histone Modification of CD45 Gene. Genes, 12(5), 759. https://doi.org/10.3390/genes12050759