
Transcriptional Pausing and Activation at Exons-1 and -2, Respectively, Mediate the MGMT Gene Expression in Human Glioblastoma Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genomic Databases
2.2. Transcription Factor Binding Sites (TFBSs) and CpG Islands
2.3. Glioblastoma Cell Culture
2.4. Western Blotting (WB)
2.5. RNA Extraction, Conventional RT-PCR, and RT-qPCR
2.6. Nuclear Run-On (NRO) Transcription Assays
2.7. Statistical Analysis
3. Results
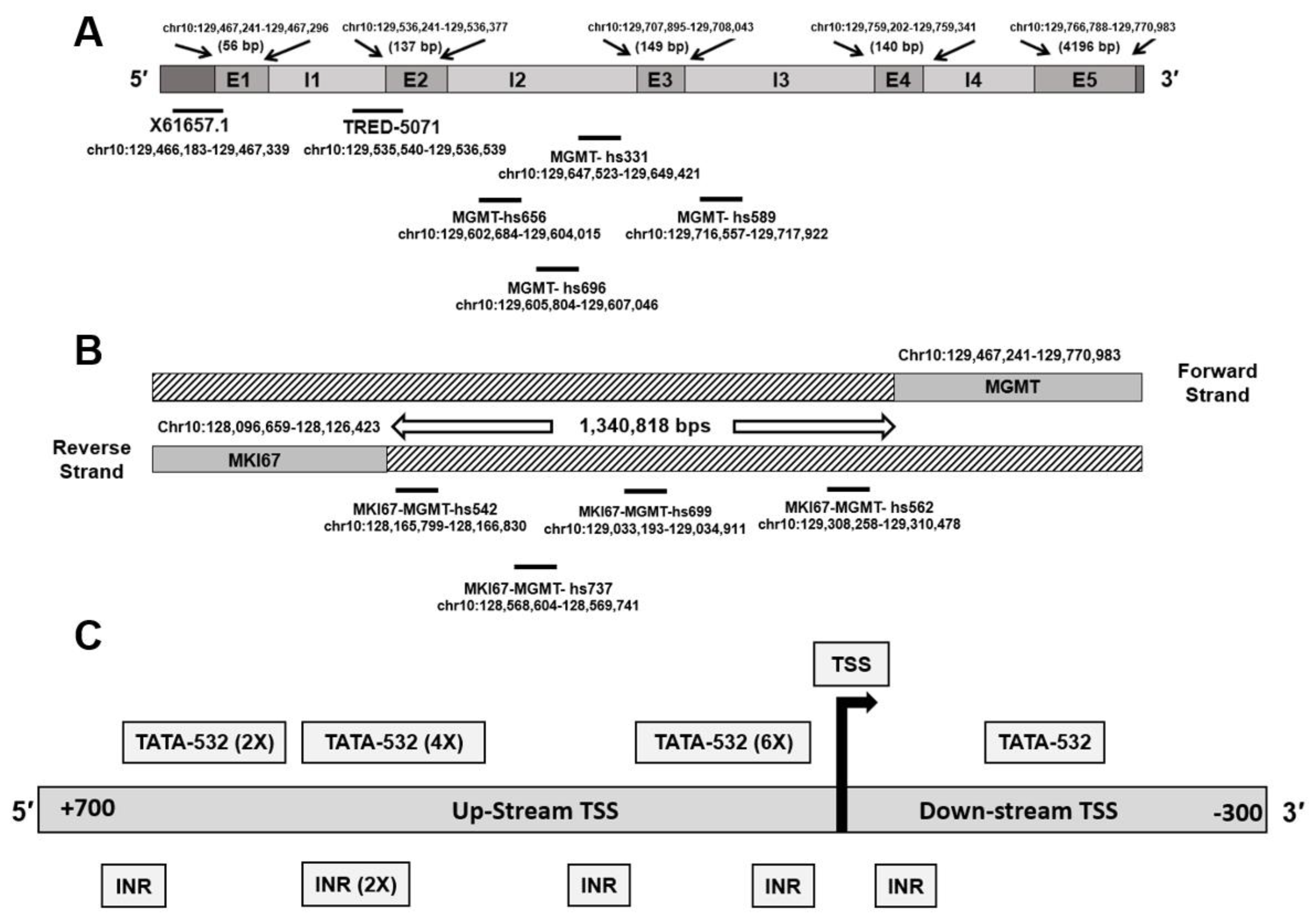
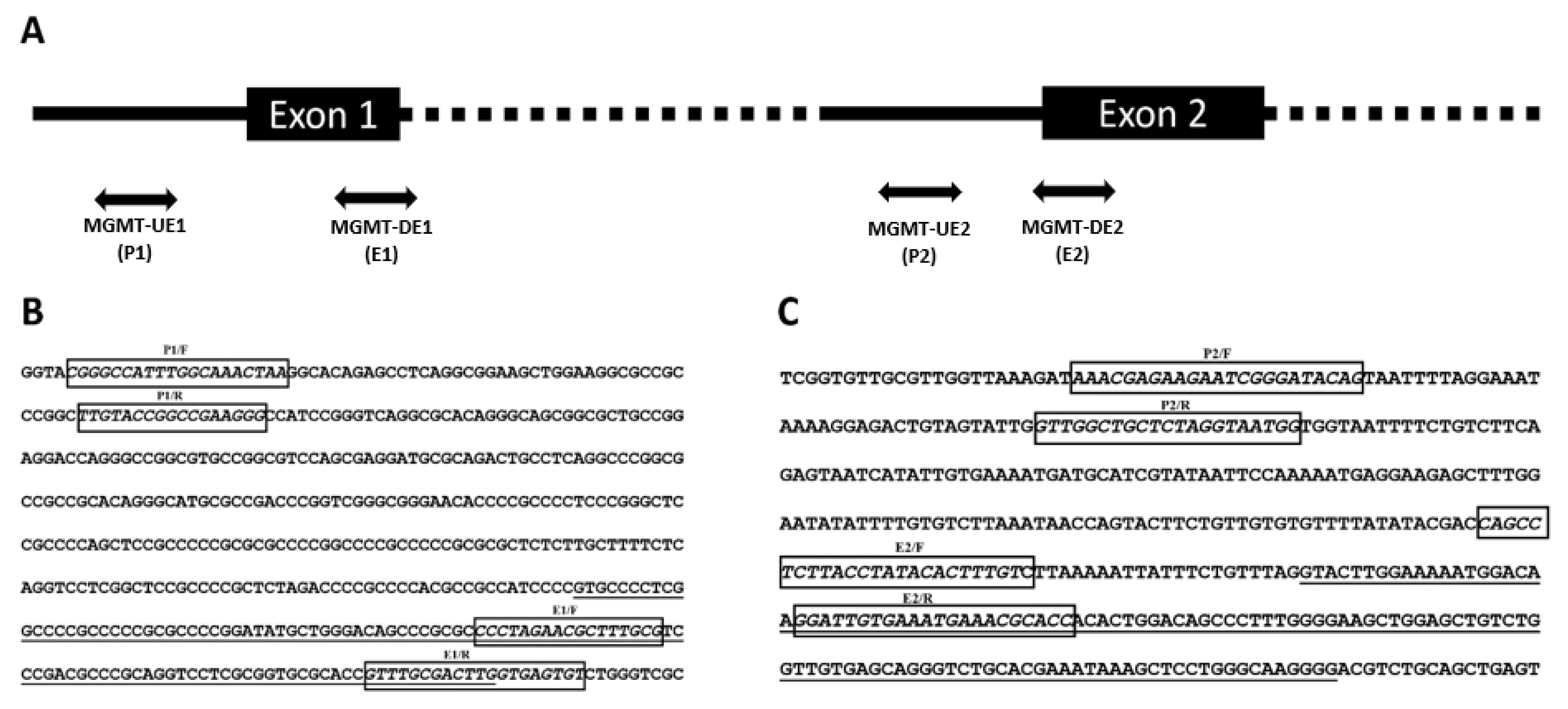
3.1. Genomic Context of MGMT Locus, MGMT Promoters, and Enhancer Sequences in the Vicinity
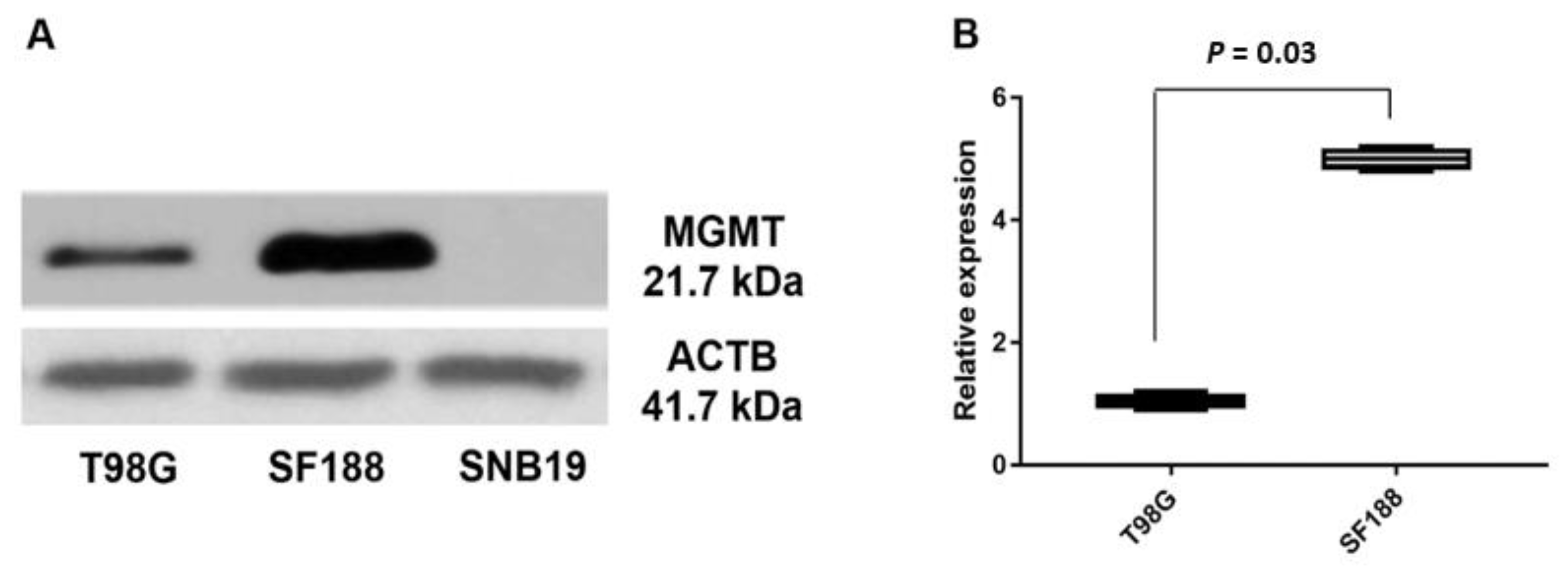
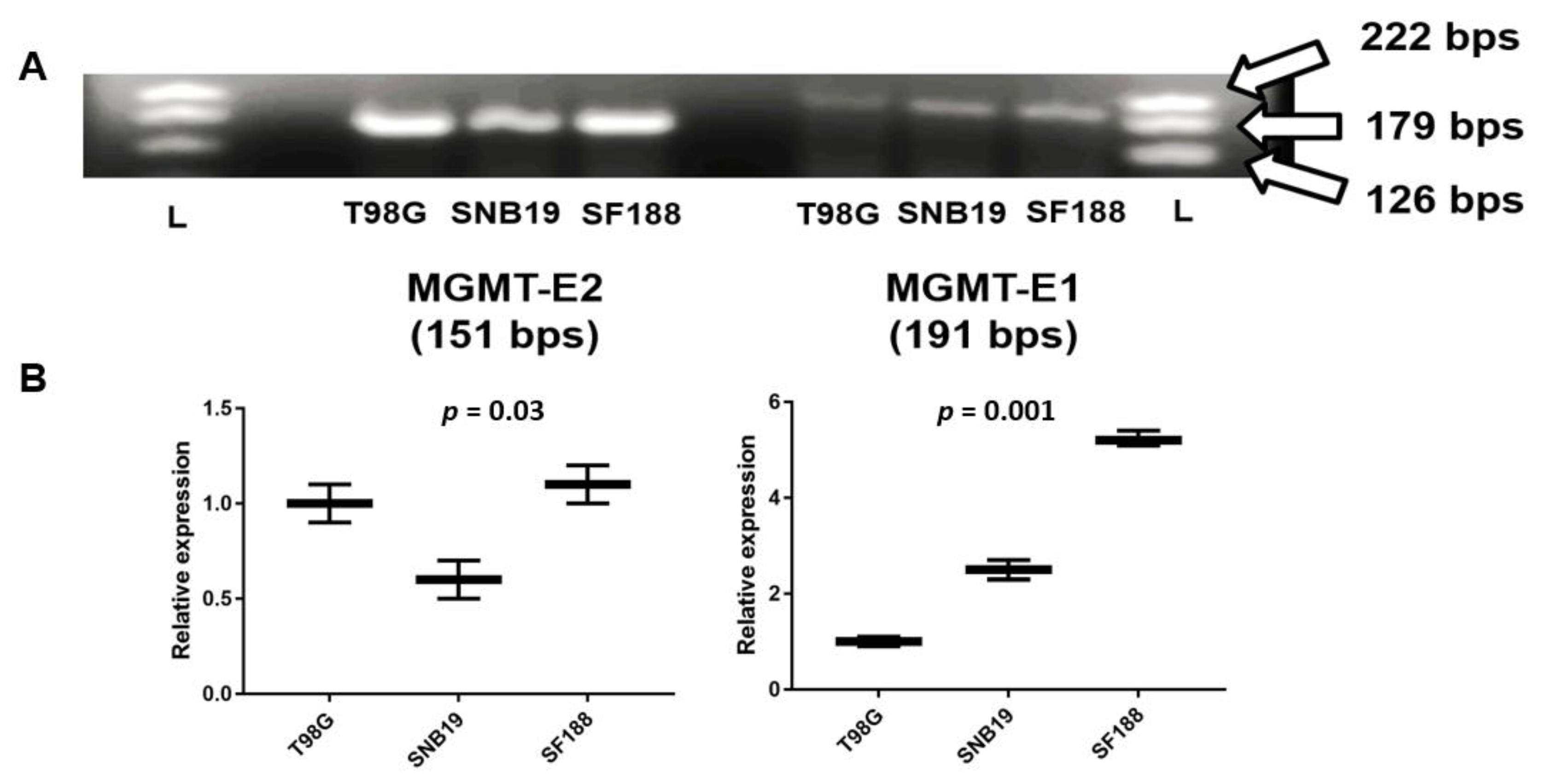
3.2. Expression of MGMT Promoters Proximal to Exon 1 and Exon 2 in MGMT-Proficient and -Deficient GBM Cell Lines
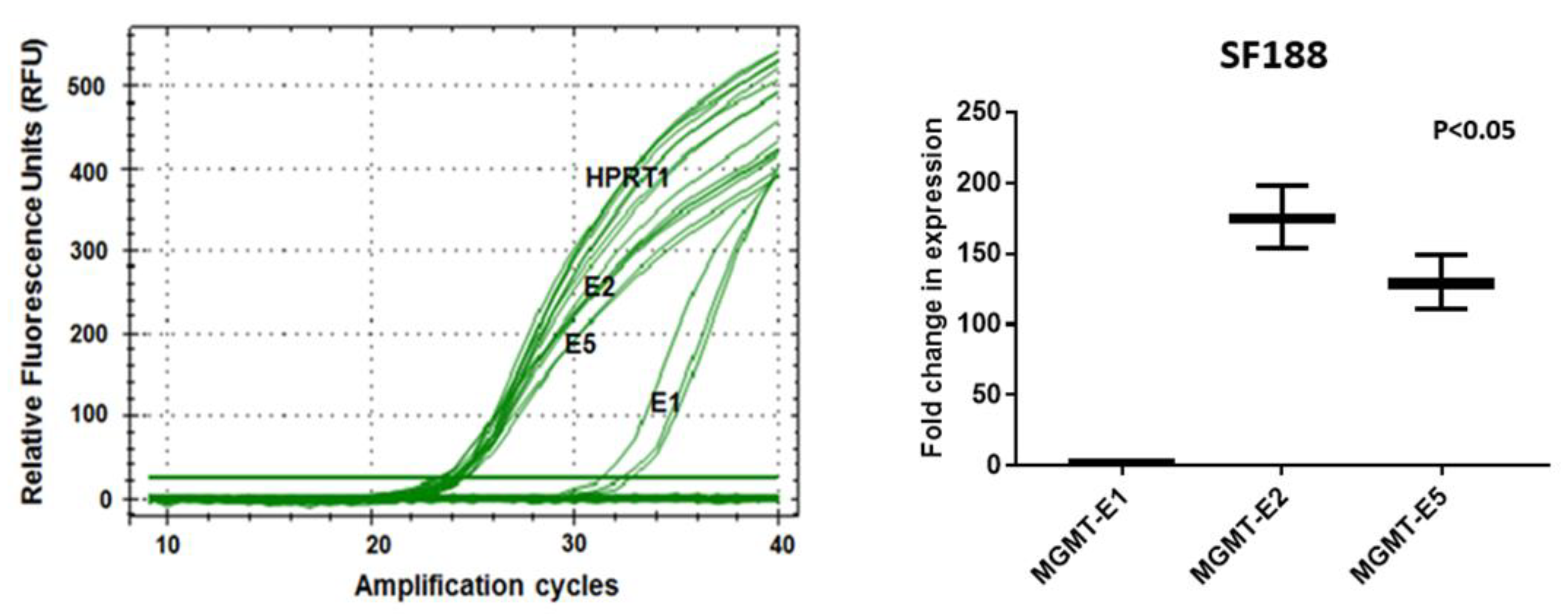
3.3. RT q-PCR Analysis of MGMT-E1, MGMT-E2, and MGMT-E5 Expression
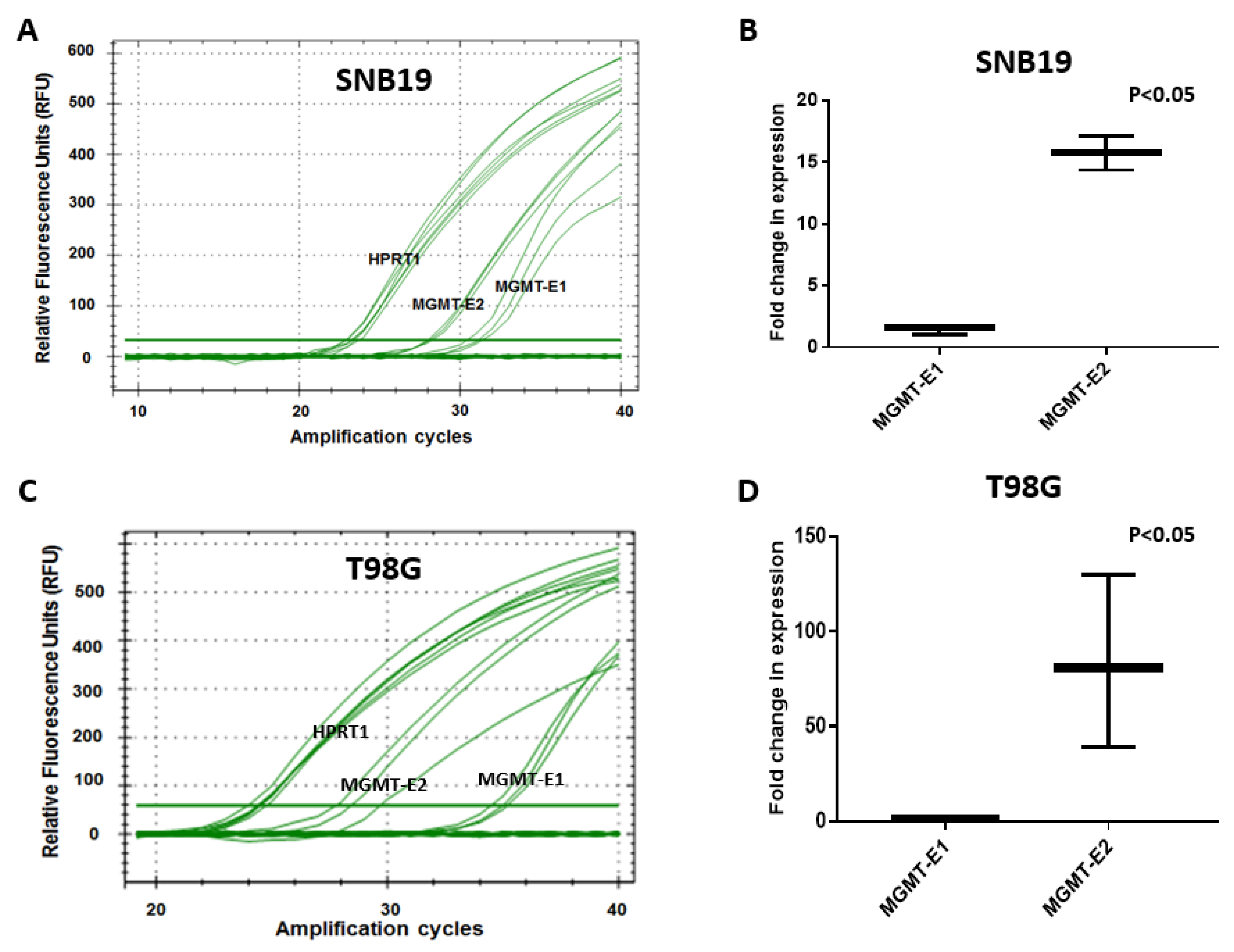
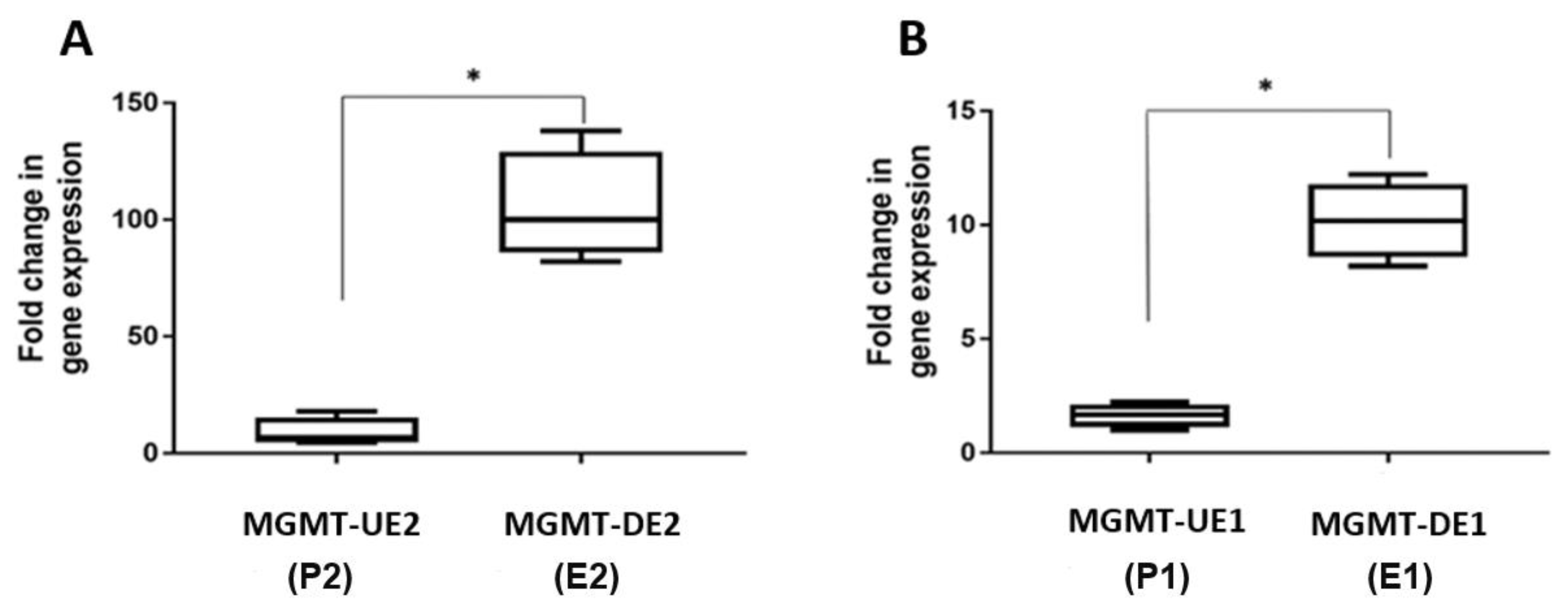
3.4. Quantitation of MGMT-E1 and MGMT-E2 Transcription Rates by Nuclear Run-On (NRO) Transcription Assays
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pegg, A.E. Repair of O6-alkylguanine by alkyltransferases. Mutat. Res. Mutat. Res. 2000, 462, 83–100. [Google Scholar] [CrossRef]
- Kaina, B.; Christmann, M. DNA repair in personalized brain cancer therapy with temozolomide and nitrosoureas. DNA Repair 2019, 78, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Mishina, Y.; Duguid, E.M.; He, C. Direct Reversal of DNA Alkylation Damage. Chem. Rev. 2006, 106, 215–232. [Google Scholar] [CrossRef]
- Gerson, S.L. Clinical Relevance of MGMT in the Treatment of Cancer. J. Clin. Oncol. 2002, 20, 2388–2399. [Google Scholar] [CrossRef]
- Kaina, B.; Margison, G.P.; Christmann, M. Targeting O6-methylguanine-DNA methyltransferase with specific inhibitors as a strategy in cancer therapy. Cell Mol. Life Sci. 2010, 67, 3663–3681. [Google Scholar] [CrossRef]
- Sarkaria, J.N.; Kitange, G.J.; James, C.D.; Plummer, R.; Calvert, H.; Weller, M.; Wick, W. Mechanisms of chemoresistance to alkylating agents in malignant glioma. Clin. Cancer Res. 2008, 14, 2900–2908. [Google Scholar] [CrossRef] [Green Version]
- Tubbs, J.L.; Pegg, A.E.; Tainer, J.A. DNA binding, nucleotide flipping, and the helix-turn-helix motif in base repair by O6-alkylguanine-DNA alkyltransferase and its implications for cancer chemotherapy. DNA Repair 2007, 6, 1100–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim Al-Obaide, M.A.; Arutla, V.; Bacolod, M.D.; Wang, W.; Zhang, R.; Srivenugopal, K.S. Genomic Space of MGMT in Human Glioma Revisited: Novel Motifs, Regulatory RNAs, NRF1, 2, and CTCF Involvement in Gene Expression. Int. J. Mol. Sci. 2021, 22, 2492. [Google Scholar] [CrossRef]
- Harris, L.C.; Potter, P.M.; Tano, K.; Shiota, S.; Mitra, S.; Brent, T.P. Characterization of the promoter region of the human O6-methylguanine-DNA methyltransferase gene. Nucleic Acids Res. 1991, 19, 6163–6167. [Google Scholar] [CrossRef] [Green Version]
- Cabrini, G.; Fabbri, E.; Lo Nigro, C.; Dechecchi, M.C.; Gambari, R. Regulation of expression of O6-methylguanine-DNA methyltransferase and the treatment of glioblastoma. Int. J. Oncol. 2015, 47, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Binabaj, M.M.; Bahrami, A.; ShahidSales, S.; Joodi, M.; Joudi Mashhad, M.; Hassanian, S.M.; Anvari, K.; Avan, A. The prognostic value of MGMT promoter methylation in glioblastoma: A meta-analysis of clinical trials. J. Cell Physiol. 2018, 233, 378–386. [Google Scholar] [CrossRef]
- Bacolod, M.D.; Barany, F. MGMT Epigenetics: The Influence of Gene Body Methylation and other Insights Derived from Integrated Methylomic, Transcriptomic, and Chromatin Analyses for Various Cancer Types. Curr. Cancer Drug Targets 2021, in press. [Google Scholar] [CrossRef]
- Weller, M.; Stupp, R.; Reifenberger, G.; Brandes, A.A.; van den Bent, M.J.; Wick, W.; Hegi, M.E. MGMT promoter methylation in malignant gliomas: Ready for personalized medicine? Nat. Rev. Neurol. 2010, 6, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Costello, J.F.; Futscher, B.W.; Tano, K.; Graunke, D.M.; Pieper, R.O. Graded methylation in the promoter and body of the O6-methylguanine DNA methyltransferase (MGMT) gene correlates with MGMT expression in human glioma cells. J. Biol. Chem. 1994, 269, 17228–17237. [Google Scholar] [CrossRef]
- Pandith, A.A.; Qasim, I.; Zahoor, W.; Shah, P.; Bhat, A.R.; Sanadhya, D.; Shah, Z.A.; Naikoo, N.A. Concordant association validates MGMT methylation and protein expression as favorable prognostic factors in glioma patients on alkylating chemotherapy (Temozolomide). Sci. Rep. 2018, 8, 6704. [Google Scholar] [CrossRef] [Green Version]
- Everhard, S.; Tost, J.; El Abdalaoui, H.; Crinière, E.; Busato, F.; Marie, Y.; Gut, I.G.; Sanson, M.; Mokhtari, K.; Laigle-Donadey, F.; et al. Identification of regions correlating MGMT promoter methylation and gene expression in glioblastomas. Neuro Oncol. 2009, 11, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uno, M.; Oba-Shinjo, S.M.; Camargo, A.A.; Moura, R.P.; Aguiar, P.H.; Cabrera, H.N.; Begnami, M.; Rosemberg, S.; Teixeira, M.J.; Marie, S.K. Correlation of MGMT promoter methylation status with gene and protein expression levels in glioblastoma. Clinics 2011, 66, 1747–1755. [Google Scholar] [CrossRef] [Green Version]
- Bobola, M.S.; Alnoor, M.; Chen, J.Y.; Kolstoe, D.D.; Silbergeld, D.L.; Rostomily, R.C.; Blank, A.; Chamberlain, M.C.; Silber, J.R. O6-methylguanine-DNA methyltransferase activity is associated with response to alkylating agent therapy and with MGMT promoter methylation in glioblastoma and anaplastic glioma. BBA Clin. 2015, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Moen, E.L.; Stark, A.L.; Zhang, W.; Dolan, M.E.; Godley, L.A. The role of gene body cytosine modifications in MGMT expression and sensitivity to temozolomide. Mol. Cancer Ther. 2014, 13, 1334–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malta, T.M.; De Souza, C.F.; Sabedot, T.S.; Silva, T.C.; Mosella, M.S.; Kalkanis, S.N.; Snyder, J.; Castro, A.V.B.; Noushmehr, H. Glioma CpG island methylator phenotype (G-CIMP): Biological and clinical implications. Neuro Oncol. 2018, 20, 608–620. [Google Scholar] [CrossRef]
- Esteller, M.; Garcia-Foncillas, J.; Andion, E.; Goodman, S.N.; Hidalgo, O.F.; Vanaclocha, V.; Baylin, S.B.; Herman, J.G. Inactivation of the DNA-Repair Gene MGMT and the Clinical Response of Gliomas to Alkylating Agents. N. Engl. J. Med. 2000, 343, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Krex, D.; Klink, B.; Hartmann, C.; von Deimling, A.; Pietsch, T.; Simon, M.; Sabel, M.; Steinbach, J.P.; Heese, O.; Reifenberger, G.; et al. German Glioma Network. Long-term survival with glioblastoma multiforme. Brain 2007, 130, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT Gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Xuan, Z.; Zhao, F.; Zhang, M.Q. TRED: A transcriptional regulatory element database, new entries, and other development. Nucleic Acids Res. 2007, 35, D137–D140. [Google Scholar] [CrossRef] [PubMed]
- Srivenugopal, K.S.; Mullapudi, S.R.; Shou, J.; Hazra, T.K.; Ali-Osman, F. Protein phosphorylation is a regulatory mechanism for O6-alkylguanine-DNA alkyltransferase in human brain tumor cells. Cancer Res. 2000, 60, 282–287. [Google Scholar]
- Valente, V.; Teixeira, S.A.; Neder, L.; Okamoto, O.K.; Oba-Shinjo, S.M.; Marie, S.K.; Scrideli, C.A.; Paçó-Larson, M.L.; Carlotti, C.G., Jr. Selection of suitable housekeeping genes for expression analysis in glioblastoma using quantitative RT-PCR. BMC Mol. Biol. 2009, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Smale, S.T. Nuclear Run-on Assay. Cold Spring Harb. Protoc. 2009. [Google Scholar] [CrossRef]
- Roberts, T.C.; Hart, J.R.; Kaikkonen, M.U.; Weinberg, M.S.; Vogt, P.K.; Morris, K.V. Quantification of nascent transcription by bromouridine immunocapture nuclear run-on RT-qPCR. Nat. Protoc. 2015, 10, 1198–1211. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-Y.; Shiekhattar, R. Architectural and functional commonalities between enhancers and promoters. Cell 2015, 162, 948–959. [Google Scholar] [CrossRef] [Green Version]
- Shlyueva, D.; Stampfel, G.; Stark, A. Transcriptional enhancers: From properties to genome-wide predictions. Nat. Rev. Genet. 2014, 15, 272–286. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, M.; Gan, H.; Wang, H.; Lee, J.-H.; Fang, D.; Kitange, G.J.; He, L.; Hu, Z.; Parney, I.F.; et al. A novel enhancer regulates MGMT expression and promotes temozolomide resistance in glioblastoma. Nat. Commun. 2018, 9, 2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, N.; Thillet, J.L.; Wanherdrick, K.; Idbaih, A.; Legrier, M.-E.; Dutrillaux, B.; Sanson, M.; Poupon, M.F. Genetic alterations associated with acquired temozolomide resistance in SNB-19, a human glioma cell line. Mol. Cancer Ther. 2006, 5, 2182–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Risueno, A.; Roson-Burgo, B.; Dolnik, A.; Hernandez-Rivas, J.M.; Bullinger, L.; Rivas, J.D.L. A robust estimation of exon expression to identify alternatively spliced genes applied to human tissues and cancer samples. BMC Genom. 2014, 15, 879. [Google Scholar] [CrossRef] [Green Version]
- Marzluff, W.F.; Huang, R.C.C. Transcription of RNA in isolated nuclei. In Transcription and Translation, a Practical Approach; Hames, B.D., Higgins, S.J., Eds.; IRL Press: Oxford, UK, 1986; pp. 89–128. [Google Scholar]
- Harris, L.C.; Remack, J.S.; Brent, T.P. In vitro methylation of the O6-methylguanine-DNA methyltransferase promoter reduces transcription. Biochim. Biophys. Acta 1994, 1217, 141–146. [Google Scholar]
- Paranjpe, A.; Bailey, N.I.; Konduri, S.; Bobustuc, G.C.; Ali-Osman, F.; Yusuf, M.A.; Punganuru, S.R.; Madala, H.R.; Basak, D.; Mostofa, A.; et al. New insights into estrogenic regulation of O6-methylguanine DNA-methyltransferase (MGMT) in human breast cancer cells: Co-degradation of ER- and MGMT proteins by fulvestrant or O6-benzylguanine indicates fresh avenues for therapy. J. Biomed. Res. 2016, 30, 393–410. [Google Scholar]
- Fornace, A.J., Jr.; Papathanasiou, M.A.; Hollander, M.C.; Yarosh, D.B. Expression of the O6-methylguanine-DNA methyltransferase gene MGMT in MER+ and MER- human tumor cells. Cancer Res. 1990, 50, 7908–7911. [Google Scholar]
- Butler, J.E.F.; Kadonaga, J.T. The RNA polymerase II core promoter: A key component in the regulation of gene expression. Genes Dev. 2002, 16, 2583–2592. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.; Landry, H.M.; Churchman, L.S. Pause & Go: From the discovery of RNA polymerase pausing to its functional implications. Curr. Opin. Cell Biol. 2017, 46, 72–80. [Google Scholar]
- Strobl, L.J.; Eick, D. Hold back of RNA polymerase II at the transcription start site mediates down-regulation of c-myc in vivo. EMBO J. 1992, 11, 3307–3314. [Google Scholar] [CrossRef]
- Krumm, A.; Meulia, T.; Brunvand, M.; Groudine, M. The block to transcriptional elongation within the human c-myc gene is determined in the promoter-proximal region. Genes Dev. 1992, 6, 2201–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, D.L.; Groudine, M. Novel promoter upstream of the human c-myc gene and regulation of c-myc expression in B-cell lymphomas. Mol. Cell Biol. 1986, 6, 3481–3489. [Google Scholar] [CrossRef] [Green Version]
- Ko, L.J.; Engel, J.D. DNA-binding specificities of the GATA transcription factor family. Mol. Cell. Biol. 1993, 13, 4011–4022. [Google Scholar] [CrossRef] [Green Version]
- Granok, H.; Leibovitch, B.A.; Shaffer, C.D.; Elgin, S.C. Chromatin. Gaga over GAGA factor. Curr. Biol. 1995, 5, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.-Y.; Chang, Y.-L.; Swamy, K.B.S.; Chiang, R.-L.; Huang, D.-H. GAGA factor, a positive regulator of global gene expression, modulates transcriptional pausing and organization of upstream nucleosomes. Epigenetics Chromatin 2016, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Giardino-Torchia, M.L.; Ashwell, J.D. Getting MAD at MYC. Proc. Natl. Acad. Sci. USA 2018, 115, 9821–9823. [Google Scholar] [CrossRef] [Green Version]
- Georgakilas, G.K.; Perdikopanis, N.; Hatzigeorgiou, A. Solving the transcription start site identification problem with ADAPT-CAGE: A machine learning algorithm for the analysis of CAGE data. Sci. Rep. 2020, 10, 877. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | ID | Bracketing Genes | Coordinates (hg38) | References |
|---|---|---|---|---|
| Promoter | MGMT-E1 promoter (X61657.1) | MGMT (5′ UTR) | chr10:129,466,183–129,467,339 | Harris et al. 1991 [9] |
| promoter | MGMT-E2 promoter (TRED-5071) | MGMT | chr10:129,535,540–129,536,539 | TRED-5071, [8] and this study |
| Enhancer | MGMT-hs331 | MGMT (intragenic) | chr10:129,647,523–129,649,421 | VISTA Enhancer Browser https://enhancer.lbl.gov, accessed on 14 May 2021. |
| Enhancer | MKI67-MGMT-hs542 | MKI67-MGMT | chr10:128,165,799–128,166,830 | |
| Enhancer | MKI67-MGMT- hs562 | MKI67-MGMT | chr10:129,308,258–129,310,478 | |
| Enhancer | MGMT- hs589 | MGMT (intragenic) | chr10:129,716,557–129,717,922 | |
| Enhancer | MGMT-hs656 | MGMT (intragenic) | chr10:129,602,684–129,604,015 | |
| Enhancer | MGMT-hs696 | MGMT (intragenic) | chr10:129,605,804–129,607,046 | |
| Enhancer | MKI67-MGMT-hs699 | MKI67-MGMT | chr10:129,033,193–129,034,911 | |
| Enhancer | MKI67-MGMT- hs737 | MKI67-MGMT | chr10:128,568,604–128,569,741 | |
| Enhancer | MKI67-MGMT | MKI167- MGMT | Chr10:130,704,894– 130,708,206 | Chen et al. 2018 [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Obaide, M.A.I.; Srivenugopal, K.S. Transcriptional Pausing and Activation at Exons-1 and -2, Respectively, Mediate the MGMT Gene Expression in Human Glioblastoma Cells. Genes 2021, 12, 888. https://doi.org/10.3390/genes12060888
Al-Obaide MAI, Srivenugopal KS. Transcriptional Pausing and Activation at Exons-1 and -2, Respectively, Mediate the MGMT Gene Expression in Human Glioblastoma Cells. Genes. 2021; 12(6):888. https://doi.org/10.3390/genes12060888
Chicago/Turabian StyleAl-Obaide, Mohammed A. Ibrahim, and Kalkunte S. Srivenugopal. 2021. "Transcriptional Pausing and Activation at Exons-1 and -2, Respectively, Mediate the MGMT Gene Expression in Human Glioblastoma Cells" Genes 12, no. 6: 888. https://doi.org/10.3390/genes12060888
APA StyleAl-Obaide, M. A. I., & Srivenugopal, K. S. (2021). Transcriptional Pausing and Activation at Exons-1 and -2, Respectively, Mediate the MGMT Gene Expression in Human Glioblastoma Cells. Genes, 12(6), 888. https://doi.org/10.3390/genes12060888