
Gene Conversion amongst Alu SINE Elements


Abstract
:1. Introduction
2. Materials and Methods
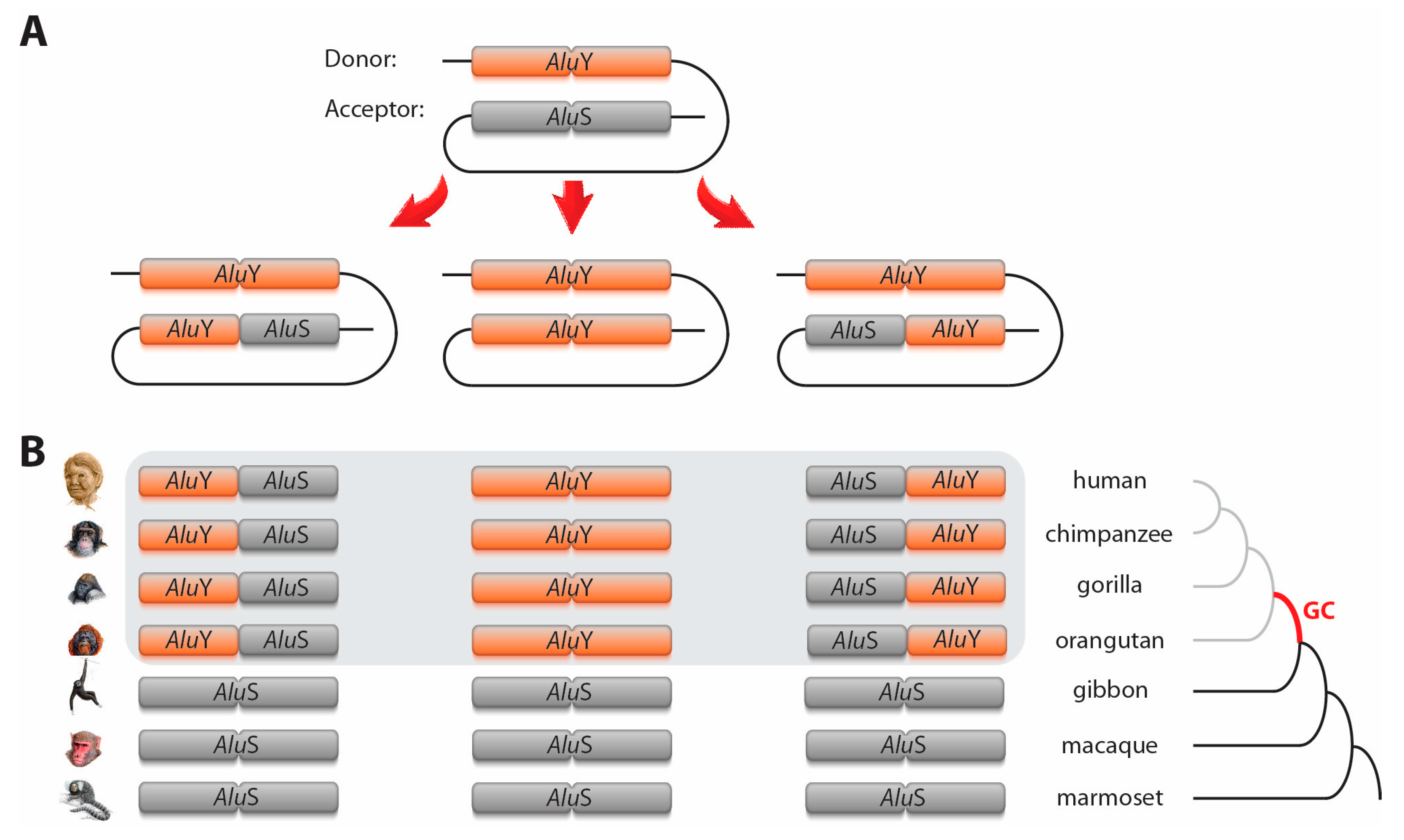
2.1. Hg38 Targeting AluY/Yc and AluS Sequences
2.2. Screening for Orthologous AluS/Y and AluY/Yc in Hominoidea
2.3. Detecting and Processing Cases of Alu Gene Conversion
2.4. Screening for Hybrid Elements in Human
2.5. Screening for Potential Polymorphic Gene Conversion in the Human Population
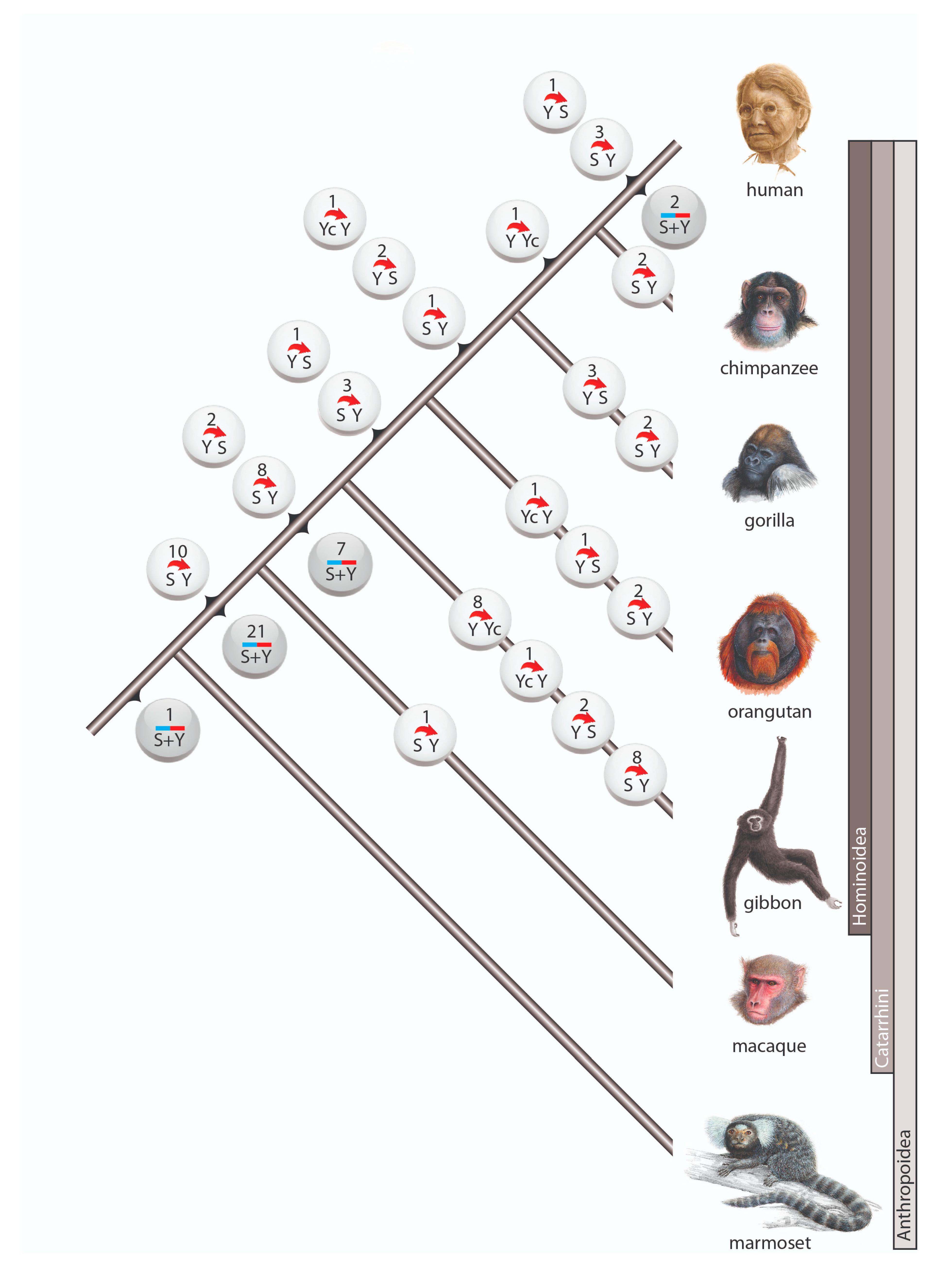
2.6. Counting of Gene Converted Alu TEs
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Koning, A.P.; Gu, W.; Castoe, T.A.; Batzer, M.A.; Pollock, D.D. Repetitive elements may comprise over two-thirds of the human genome. PLoS Genet. 2011, 7, e1002384. [Google Scholar] [CrossRef] [Green Version]
- Brookfield, J.F.; Johnson, L.J. The evolution of mobile DNAs: When will transposons create phylogenies that look as if there is a master gene? Genetics 2006, 173, 1115–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, A.; Rozhdestvensky, T.S.; Kuryshev, V.Y.; Schmitz, J.; Brosius, J. An unusual primate locus that attracted two independent Alu insertions and facilitates their transcription. J. Mol. Biol. 2005, 350, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.; Jurka, J. The age of Alu subfamilies. J. Mol. Evol. 1996, 42, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.R.; Turner, B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Kass, D.H.; Batzer, M.A.; Deininger, P.L. Gene conversion as a secondary mechanism of short interspersed element (SINE) evolution. Mol. Cell. Biol. 1995, 15, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.M.; Carroll, M.L.; Nguyen, S.V.; Salem, A.H.; Oldridge, M.; Wilkie, A.O.M.; Batzer, M.A.; Deininger, P.L. Potential gene conversion and source genes for recently integrated Alu elements. Genome Res. 2000, 10, 1485–1495. [Google Scholar] [CrossRef] [Green Version]
- Aleshin, A.; Zhi, D. Recombination-associated sequence homogenization of neighboring Alu elements: Signature of nonallelic gene conversion. Mol. Biol. Evol. 2010, 27, 2300–2311. [Google Scholar] [CrossRef] [Green Version]
- Kijima, T.E.; Innan, H. On the estimation of the insertion time of LTR retrotransposable elements. Mol. Biol. Evol. 2010, 27, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Trombetta, B.; Fantini, G.; D’Atanasio, E.; Sellitto, D.; Cruciani, F. Evidence of extensive non-allelic gene conversion among LTR elements in the human genome. Sci. Rep. 2016, 6, 28710. [Google Scholar] [CrossRef]
- Cossu, R.M.; Casola, C.; Giacomello, S.; Vidalis, A.; Scofield, D.G.; Zuccolo, A. LTR retrotransposons show low levels of unequal recombination and high rates of intraelement gene conversion in large plant genomes. Genome Biol. Evol. 2017, 9, 3449–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawcett, J.A.; Innan, H. The role of gene conversion between transposable elements in rewiring regulatory networks. Genome Biol. Evol. 2019, 11, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, C.E.; Bachtrog, D. Non-allelic gene conversion enables rapid evolutionary change at multiple regulatory sites encoded by transposable elements. eLife 2015, 4, e05899. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Styles, P.; Brookfield, J.F. Source gene composition and gene conversion of the AluYh and AluYi lineages of retrotransposons. BMC Evol. Biol. 2009, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Cordaux, R. Gene conversion maintains nonfunctional transposable elements in an obligate mutualistic endosymbiont. Mol. Biol. Evol. 2009, 26, 1679–1682. [Google Scholar] [CrossRef] [Green Version]
- Benovoy, D.; Drouin, G. Ectopic gene conversions in the human genome. Genomics 2009, 93, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Cooper, D.N.; Chuzhanova, N.; Férec, C.; Patrinos, G.P. Gene conversion: Mechanisms, evolution and human disease. Nat. Rev. Genet. 2007, 8, 762–775. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Matzke, A.; Churakov, G.; Kuritzin, A.; Mayr, G.; Brosius, J.; Schmitz, J. Waves of genomic hitchhikers shed light on the evolution of gamebirds (Aves: Galliformes). BMC Evol. Biol. 2007, 7, 190. [Google Scholar] [CrossRef] [Green Version]
- Deininger, P. Alu elements: Know the SINEs. Genome Biol. 2011, 12, 236. [Google Scholar] [CrossRef] [Green Version]
- Batzer, M.A.; Rubin, C.M.; Hellmann-Blumberg, U.; Alegria-Hartman, M.; Leeflang, E.P.; Stern, J.D.; Bazan, H.A.; Shaikh, T.H.; Deininger, P.L.; Schmid, C.W. Dispersion and insertion polymorphism in two small subfamilies of recently amplified human Alu Repeats. J. Mol. Biol. 1995, 247, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Salem, A.-H.; Ray, D.A.; Hedges, D.J.; Jurka, J.; Batzer, M.A. Analysis of the human Alu Ye lineage. BMC Evol. Biol. 2005, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doronina, L.; Reising, O.; Clawson, H.; Ray, D.A.; Schmitz, J. True homoplasy of retrotransposon insertions in primates. Syst. Biol. 2019, 68, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Doronina, L.; Matzke, A.; Churakov, G.; Stoll, M.; Huge, A.; Schmitz, J. The beaver’s phylogenetic lineage illuminated by retroposon reads. Sci. Rep. 2017, 7, 43562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churakov, G.; Zhang, F.; Grundmann, N.; Makalowski, W.; Noll, A.; Doronina, L.; Schmitz, J. The multicomparative 2-n-way genome suite. Genome Res. 2020, 30, 1508–1516. [Google Scholar] [CrossRef]
- Churakov, G.; Grundmann, N.; Kuritzin, A.; Brosius, J.; Makalowski, W.; Schmitz, J. A novel web-based TinT application and the chronology of the Primate Alu retroposon activity. BMC Evol. Biol. 2010, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- Schildkraut, E.; Miller, C.A.; Nickoloff, J.A. Transcription of a donor enhances its use during double-strand break-induced gene conversion in human cells. Mol. Cell. Biol. 2006, 26, 3098–3105. [Google Scholar] [CrossRef] [Green Version]
- Meyer, T.J.; McLain, A.T.; Oldenburg, J.M.; Faulk, C.; Bourgeois, M.G.; Conlin, E.M.; Mootnick, A.R.; De Jong, P.J.; Roos, C.; Carbone, L.; et al. An Alu-based phylogeny of gibbons (Hylobatidae). Mol. Biol. Evol. 2012, 29, 3441–3450. [Google Scholar] [CrossRef] [Green Version]
- Wacholder, A.; Pollock, D.D. PRDM9 and an epidemic of gene conversion and non-homologous recombination among Alu elements in ancestral gorillas. bioRxiv 2017, 241356. [Google Scholar]
- Jedlicka, P.; Lexa, M.; Kejnovsky, E. What can long terminal repeats tell us about the age of LTR retrotransposons, gene conversion and ectopic recombination? Front. Plant Sci. 2020, 11, 644. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Donor | Acceptor | Replaced Part of Alu | Number of Loci |
|---|---|---|---|
| Gene conversion with identified direction | |||
| AluY | AluS | Complete Alu | 11 |
| AluY | AluS | 3′-Alu unit (S-Y hybrid) | 23 |
| AluY | AluS | 5′-Alu unit (Y-S hybrid) | 6 |
| AluS | AluY | Complete Alu | 3 |
| AluS | AluY | 3′-Alu unit (Y-S hybrid) | 6 |
| AluS | AluY | 5′-Alu unit (S-Y hybrid) | 3 |
| AluYc | AluY | Diagnostic indel | 9 |
| AluY | AluYc | Diagnostic indel | 3 |
| Gene conversion with unidentified direction | |||
| Unidentified | Unidentified | AluS–AluY hybrid | 31 |
| Complex scenario | Complex scenario | Multiple gene conversion | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doronina, L.; Reising, O.; Schmitz, J. Gene Conversion amongst Alu SINE Elements. Genes 2021, 12, 905. https://doi.org/10.3390/genes12060905
Doronina L, Reising O, Schmitz J. Gene Conversion amongst Alu SINE Elements. Genes. 2021; 12(6):905. https://doi.org/10.3390/genes12060905
Chicago/Turabian StyleDoronina, Liliya, Olga Reising, and Jürgen Schmitz. 2021. "Gene Conversion amongst Alu SINE Elements" Genes 12, no. 6: 905. https://doi.org/10.3390/genes12060905
APA StyleDoronina, L., Reising, O., & Schmitz, J. (2021). Gene Conversion amongst Alu SINE Elements. Genes, 12(6), 905. https://doi.org/10.3390/genes12060905