
The Association of ATG16L1 Variations with Clinical Phenotypes of Adult-Onset Still’s Disease


 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Single-Nucleotide Polymorphism Selection and Genotyping Using Direct Sequencing
2.3. Determination of the mRNA Levels of ATGs Using Quantitative PCR
2.4. Determination of Serum Levels of Proinflammatory Cytokines Using Enzyme-Linked Immunoabsorbent Assay
2.5. Statistical Analysis
3. Results
3.1. Clinical Characteristics of Patients with AOSD
3.2. Genotypes, Allele Frequencies, and Haplotypes: Comparing Patients with AOSD and Healthy Controls
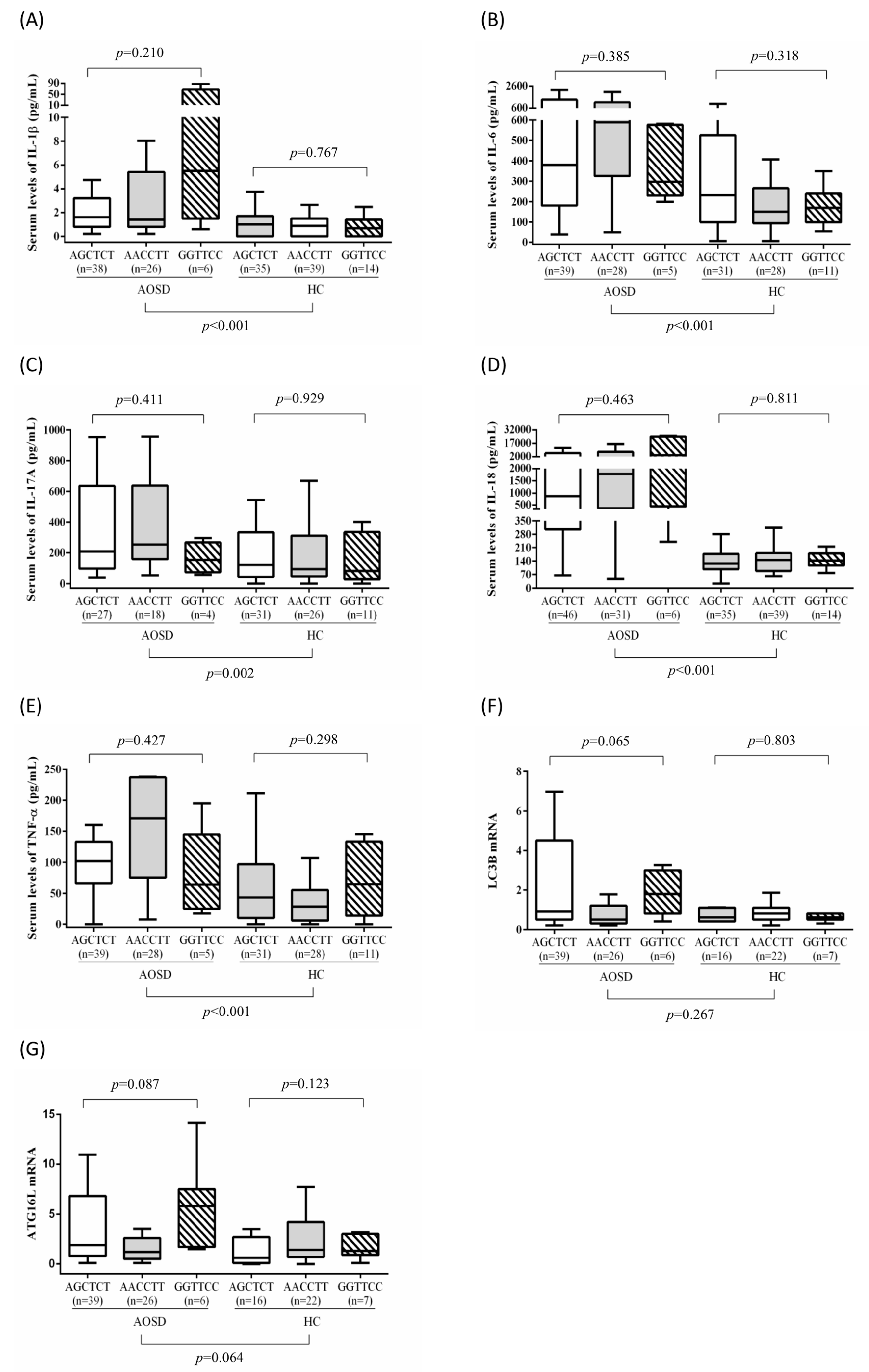
3.3. Association of ATG16L1 Polymorphisms with ATG mRNA Expression Levels and Serum Cytokine Levels in Patients with AOSD
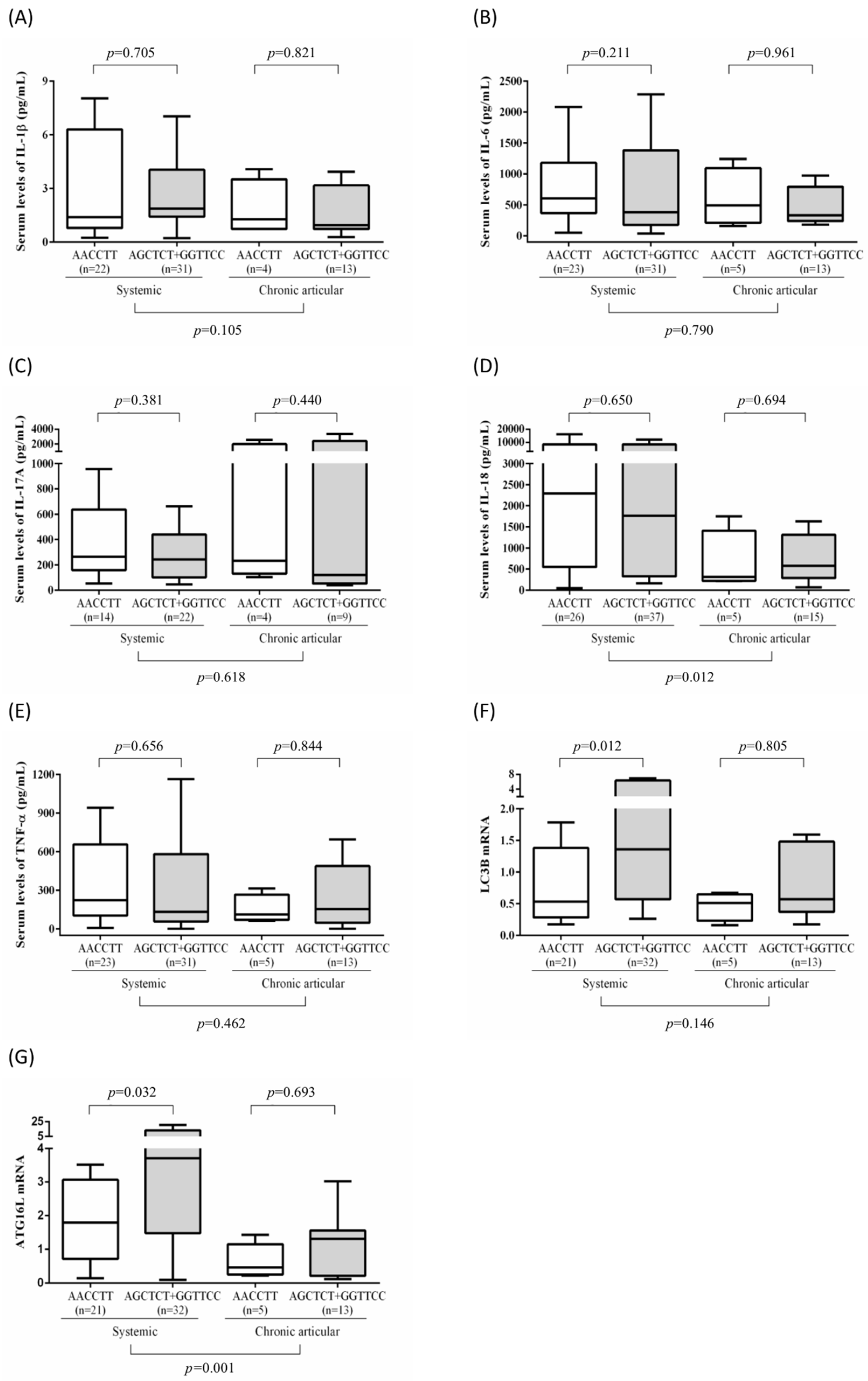
3.4. Association of Disease Outcomes with ATG16L1 Polymorphisms and Clinical Manifestations in Patients with AOSD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bywaters, E.G.L. Still’s disease of the adults. Ann. Rheum. Dis. 1971, 30, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Kadavath, S.; Efthimiou, P. Adult-onset Still’s disease-pathogenesis, clinical manifestations, and new treatment options. Ann. Med. 2015, 22, 1–9. [Google Scholar] [CrossRef]
- Kastner, D.L.; Aksentijevich, I.; Goldbach-Mansky, R. Autoinflammatory disease reloaded: A clinical perspective. Cell 2010, 140, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Balow, J.E.; Ryan, J.G.; Chae, J.J.; Booty, M.G.; Bulua, A.; Stone, D. Microarray-based gene expression profiling in patients with cryopyrine-associated periodic syndromes defines a disease-related signature and IL-1-responsive transcripts. Ann. Rheum. Dis. 2013, 72, 1064–1070. [Google Scholar] [CrossRef]
- Kawashima, M.; Yamamura, M.; Taniai, M.; Yamauchi, H.; Tanimoto, T.; Kurimoto, M.; Miyawaki, S.; Amano, T.; Takeuchi, T.; Makino, H. Levels of interleukin-18 and its binding inhibitors in the blood circulation of patients with adult-onset Still’s disease. Arthritis Rheum 2001, 44, 550–560. [Google Scholar] [CrossRef]
- Choi, J.H.; Suh, C.H.; Lee, Y.M.; Suh, Y.J.; Lee, S.K.; Kim, S.S.; Nahm, D.H.; Park, H.S. Serum cytokine profiles in patients with adult onset Still’s disease. J. Rheumatol. 2003, 30, 2422–2427. [Google Scholar] [PubMed]
- Chen, D.Y.; Lan, J.L.; Lin, F.J.; Hsieh, T.Y. Proinflammatory cytokine profiles in the sera and in the pathological tissues of patients with active untreated adult onset Still’s disease. J. Rheumatol. 2004, 31, 2189–2198. [Google Scholar] [PubMed]
- Hsieh, C.W.; Chang, C.Y.; Chen, Y.M.; Chen, H.H.; Hung, W.T.; Gung, N.R.; Wey, S.J.; Chen, D.Y. Impaired autophagic flux and its related inflammation in adult-onset Still’s disease patients. Oncotarget 2017, 9, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Fan, W.; Chen, K.; Ding, X.; Chen, S.; Zhong, Q. Identification of Barkor as a mammalian autophagy-specific factor for Beclin 1 and class III phosphatidylinositol 3-kinase. Proc. Natl. Acad. Sci. USA 2008, 105, 19211–19216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, C.W.; Fokken, C.; Turville, S.; Lünemann, A.; Schmidt, J.; Münz, C.; Lunemann, J. TNF-α induces macroautophagy and regulates MHC class II expression in human skeletal muscle cells. J. Biol. Chem. 2011, 286, 3970–3980. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.J.; Zhang, H. Autophagy in immunity: Implications in etiology of autoimmune/autoinflammatory diseases. Autophagy 2012, 8, 1286–1299. [Google Scholar] [CrossRef] [Green Version]
- Koutouzov, S.; Mathian, A.; Dalloul, A. Type-I interferons and systemic lupus erythematosus. Autoimmun Rev. 2006, 5, 554–562. [Google Scholar] [CrossRef]
- Gros, F.; Muller, S. Pharmacological regulators of autophagy and their link with modulators of lupus disease. Brit. J. Pharmacol. 2014, 171, 4337–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [Green Version]
- Romanov, J.; Walczak, M.; Ibiricu, I.; Schuchner, S.; Ogris, E.; Kraft, C.; Martens, S. Mechanism and functions of membrane binding by the Atg5-Atg12/Atg16 complex during autophagosome formation. EMBO J. 2012, 31, 4304–4317. [Google Scholar] [CrossRef] [PubMed]
- Hampe, J.; Franke, A.; Rosenstiel, P.; Till, A.; Teuber, M.; Huse, K.; Albrecht, M.; Mayr, G.; De La Vega, F.M.; Briggs, J.; et al. A genome-wide association scan of nonsynonymous SNPs identifies a susceptibility variant for Crohn disease in ATG16L1. Nat. Genet. 2007, 39, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Rioux, J.D.; Xavier, R.J.; Taylor, K.D.; Silverberg, M.S.; Goyette, P.; Huett, A.; Green, T.; Kuballa, P.; Barmada, M.M.; Datta, L.W.; et al. Genome-wide association study identifies new susceptibility loci for Crohn disease and implicates autophagy in disease pathogenesis. Nat. Genet. 2007, 39, 596–604. [Google Scholar] [CrossRef]
- McDermott, M.F.; Aksentijevich, I.; Galon, J.; McDermott, E.M.; Ogunkolade, B.W.; Centola, M. Germline mutations in the extracellular domains of the 55 kDa TNF receptor, TNFR1, define a family of dominantly inherited autoinflammatory syndromes. Cell 1999, 97, 133–144. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ohta, A.; Tsunematsu, T.; Kasukawa, R.; Mizushima, Y.; Kashiwagi, H.; Kashiwazaki, S.; Tanimoto, K.; Matsumoto, Y.; Ota, T. Preliminary criteria for classification of adult Still’s disease. J. Rheumatol. 1992, 19, 424–430. [Google Scholar] [PubMed]
- Rau, M.; Schiller, M.; Krienke, S.; Heyder, P.; Lorenz, H.; Blank, N. Clinical manifestations but not cytokine profiles differentiate adult-onset Still’s disease and sepsis. J. Rheumatol. 2010, 37, 2369–2376. [Google Scholar] [CrossRef]
- Mavragani, C.P.; Spyridakis, E.G.; Koutsilieris, M. Adult-onset Still’s disease: From pathophysiology to targeted therapies. Int. J. Inflamm. 2012, 2012, 879020. [Google Scholar] [CrossRef] [Green Version]
- Gerfaud-Valentin, M.; Jamilloux, Y.; Iwaz, J.; Sève, P. Adult-onset Still’s disease. Autoimmun Rev. 2014, 13, 708–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Pham, D.; Kim, S.H.; Losol, P.; Yang, E.M.; Shin, Y.S.; Ye, Y.M.; Park, H.S. Association of autophagy related gene polymorphisms with neutrophilic airway inflammation in adult asthma. Korean J. Intern. Med. 2016, 31, 375–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.A.; Taylor, K.E.; Graham, R.R.; Nititham, J.; Lee, A.T.; Ortmann, W.A. Differential genetic associations for systemic lupus erythematosus based on anti-dsDNA autoantibody production. PLoS Genet. 2011, 7, e1001323. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, J.M.; Daley, D.; Graham, J.; Berry, B.R.; Gallagher, R.P.; Connors, J.M.; Gascoyne, R.D.; Spinelli, J.J.; Brooks-Wilson, A.R. Genetic variation in cell death genes and risk of non-Hodgkin lymphoma. PLoS ONE 2012, 7, e31560. [Google Scholar] [CrossRef] [PubMed]
- Poon, A.H.; Chouiali, F.; Tse, S.M.; Litonjua, A.A.; Hussain, S.N.; Baglole, C.J. Genetic and histologic evidence for autophagy in asthma pathogenesis. J. Allergy Clin. Immunol. 2012, 129, 569–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, P.Y.J.; Beyens, G.; Guañabens, N.; Boonen, S.; Papapoulos, S.; Karperien, M.; Eekhoff, M.; Van Wesenbeeck, L.; Jennes, K.; Geusens, P.; et al. Founder effect in different European countries for the recurrent P392L SQSTM1 mutation in Paget’s Disease of Bone. Calcif. Tissue Int. 2008, 83, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Ellwanger, D.C.; Hartsperger, M.L.; Pfeufer, A.; Stümpflen, V. Cis-acting polymorphisms affect complex traits through modifications of microRNA regulation pathways. PLoS ONE 2012, 7, e36694. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Essers, J.B.; Kugathasan, S.; Escher, J.C.; Lettre, G.; Butler, J.L.; Stephens, M.C.; Ramoni, M.F.; Grand, R.J.; Hirschhorn, J. Association of linear growth impairment in pediatric Crohn’s disease and a known height locus: A pilot study. Ann. Hum. Genet. 2010, 74, 497–498. [Google Scholar] [CrossRef] [Green Version]
- Usategui-Martín, R.; García-Aparicio, J.; Corral-Gudino, L.; Calero-Paniagua, I.; Del Pino-Montes, J.; Sarmiento, R.G. Polymorphisms in Autophagy Genes Are Associated with Paget Disease of Bone. PLoS ONE 2015, 10, e0128984. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Hung, S.I.; Chung, W.H.; Liu, Z.S.; Chen, C.H.; Hsih, M.S.; Hui, R.C.; Chu, C.Y.; Chen, Y.T. Common risk allele in aromatic antiepilep-tic-drug induced Stevens-Johnson syndrome and toxic epidermal necrolysis in Han Chinese. Pharmacogenomics 2010, 11, 349–356. [Google Scholar] [CrossRef]
- Glas, J.; Konrad, A.; Schmechel, S.; Dambacher, J.; Seiderer, J.; Schroff, F.; Wetzke, M.; Roeske, D.; Török, H.-P.; Tonenchi, L.; et al. The ATG16L1 gene variants rs2241879 and rs2241880 (T300A) are strongly associated with susceptibility to Crohn’s disease in the German population. Am. J. Gastroenterol. 2008, 103, 682–691. [Google Scholar] [CrossRef]
- Lacher, M.; Schroepf, S.; Ballauff, A.; Lohse, P.; von Schweinitz, D.; Kappler, R.; Koletzko, S. Autophagy 16-like 1 rs2241880 G allele is associated with Crohn’s disease in German children. Acta Paediatr. 2009, 98, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Wakai, K.; Ohta, A.; Tamakoshi, A.; Ohno, Y.; Kawamura, T.; Aoki, R.; Kojima, M.; Lin, Y.; Hashimoto, S.; Inaba, Y.; et al. Estimated prevalence and incidence of adult Still’s disease: Findings by a nationwide epidemiological survey in Japan. J. Epidemiol. 1997, 7, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Evensen, K.J.; Nossent, H.C. Epidemiology and outcome of adult-onset Still’s disease in Northern Norway. Scand J. Rheumatol. 2006, 35, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Fujita, N.; Jang, M.H.; Uematsu, S.; Yang, B.-G.; Satoh, T.; Omori, H.; Noda, T.; Yamamoto, N.; Komatsu, M.; et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1β production. Nature 2008, 456, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.Y.; Jung, H.Y.; Park, K.Y.; Kim, S.K. Enhanced p62 expression through impaired proteasomal degradation is involved in caspase-1 activation in monosodium urate crystal-induced interleukin-1b expression. Rheumatology 2014, 53, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.B.; Ahn, J.Y.; Lee, H.S.; Kim, T.H.; Jun, J.B.; Jung, S. S Association between Fcγ IIa and IIIa polymorphism and clinical manifestations in Korean patients with adult-onset Still’s disease. J. Korean Med. Sci. 2002, 17, 75–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.Y.; Chen, Y.M.; Chen, H.H.; Hsieh, C.W.; Lin, C.C.; Lan, J.L. Functional Association of Interleukin 18 Gene –607(C/A) Promoter Polymorphisms with Disease Course in Chinese Patients with Adult-onset Still’s Disease. J. Rheumatol. 2009, 36, 2284–2289. [Google Scholar] [CrossRef]




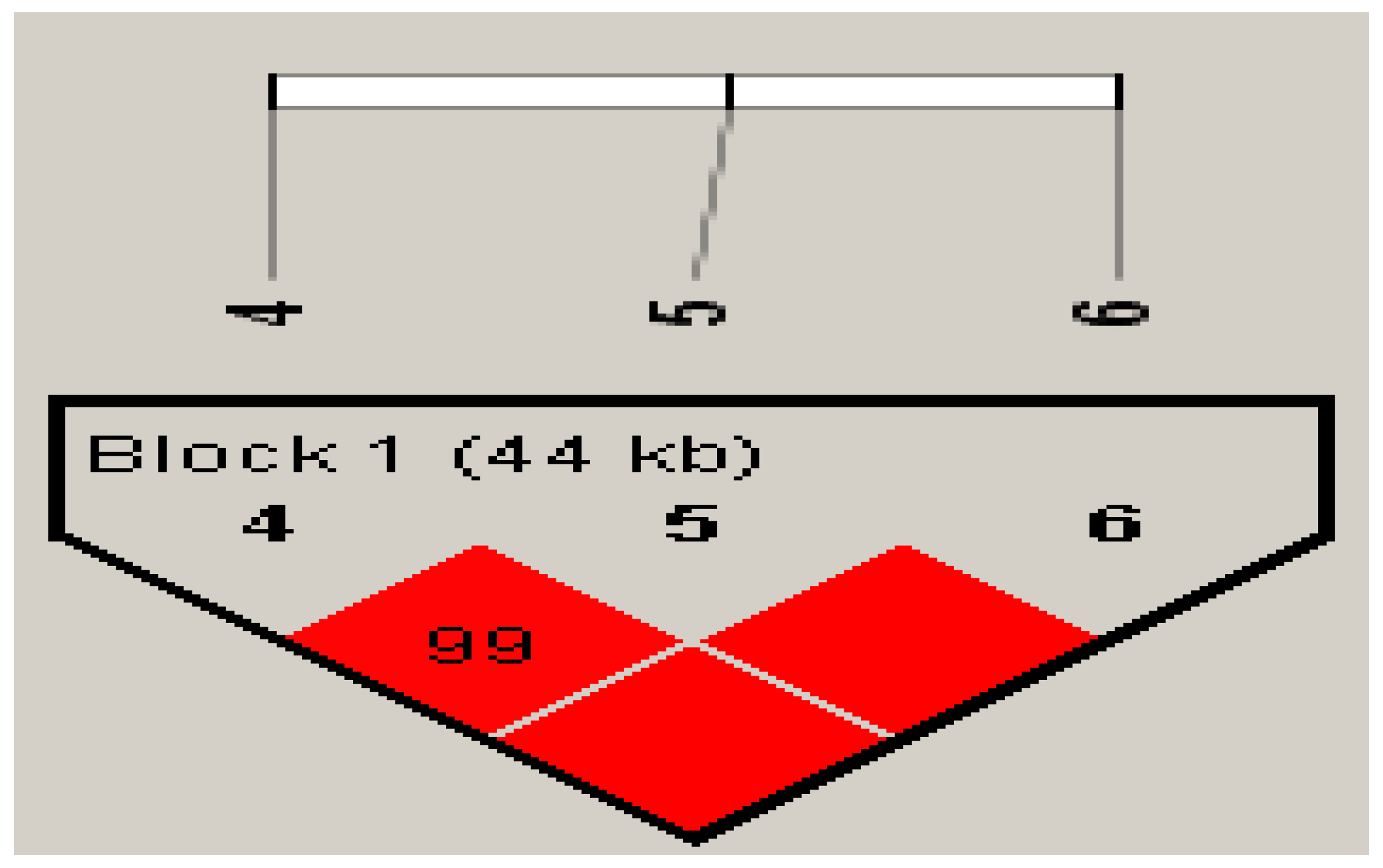{kind=link}
{kind=link}
{kind=link}
| Gene | SNP ID | TaqMan no. Chromosome: Position | Location | MAF | HWE (p-Value) |
|---|---|---|---|---|---|
| Beclin-1 | rs10512488 | C_27102741_10 17:42811886 | intron variant | A (G > A), 0.01 | 0.751 |
| ATG5 | rs573775 | C_910347_20 6:106316991 | intron variant | A (G > A), 0.33 | 0.586 |
| SQSTM1 | rs565280 | C_645001_20 5:179826926 | intron variant | A (G > A), 0.02 | 0.917 |
| ATG16L1 | rs2241880 | C_9095577_20 2:233274722 | transcript variant | G (A > G), 0.34 | 0.064 |
| rs10210302 | C_30179764_10 2:233250193 | transcript variant | T (C > T), 0.34 | 0.141 | |
| rs1045100 | C_8741775_20 2:233294951 | transcript variant | C (T > C), 0.33 | 0.201 |
| AOSD (n = 129) | HC (n = 129) | p value | OR (95% CI) | OR (95% CI) a | |||
|---|---|---|---|---|---|---|---|
| ATG16Lrs2241880 | 0.001 ** | ||||||
| GG | 7 (5.4%) | 22 (17.1%) | Reference | Reference | |||
| AG | 74 (57.4%) | 50 (38.8%) | 4.65 | (1.85–11.71) | 4.72 | (1.87–11.90) | |
| AA | 48 (37.2%) | 57 (44.2%) | 2.65 | (1.04–6.73) | 2.61 | (1.02–6.64) | |
| ATG16Lrs10210302 | 0.001 ** | ||||||
| TT | 6 (4.7%) | 21 (16.3%) | Reference | Reference | |||
| CT | 75 (58.1%) | 52 (40.3%) | 5.05 | (1.91–13.37) | 5.14 | (1.94–13.65) | |
| CC | 48 (37.2%) | 56 (43.4%) | 3.00 | (1.12–8.04) | 2.98 | (1.11–7.99) | |
| ATG16Lrs1045100 | 0.003 ** | ||||||
| TT | 49 (38.0%) | 58 (45.0%) | Reference | Reference | |||
| CT | 74 (57.4%) | 52 (40.3%) | 1.68 | (1.00–2.83) | 1.74 | (1.03–2.93) | |
| CC | 6 (4.7%) | 19 (14.7%) | 0.37 | (0.14–1.01) | 0.37 | (0.14–1.01) | |
| AOSD (n = 127) | HC (n = 124) | p-value | Logistic regression | ||||
| n (%) | n (%) | OR (95% CI) | p value | ||||
| ATG16L1 haplotypes | 0.002 | ||||||
| GG/TT/CC haplotype | 6 (4.7%) | 19 (15.3%) | 0.37 [0.14–0.997] | 0.049 * | |||
| AG/CT/CT haplotype | 73 (57.5%) | 49 (39.5%) | 1.74 [1.02–2.95] | 0.040 * | |||
| AA/CC/TT haplotype | 48 (37.8%) | 56 (45.2%) | Reference | ||||
| AOSD | |||||||
|---|---|---|---|---|---|---|---|
| AGCTCT + GGTTCC | AACCTT | p-Value | |||||
| n | Median | IQR | n | Median | IQR | ||
| LC3B mRNA | 45 | 1.1 | (1.1–4.3) | 26 | 0.5 | (0.5–1.2) | 0.026 * |
| ATG16L mRNA | 45 | 2.0 | (2.0–8.1) | 26 | 1.2 | (1.2–2.6) | 0.088 |
| Serum levels of IL-1 (pg/mL) | 44 | 1.7 | (1.7–4) | 26 | 1.4 | (1.4–5.4) | 0.947 |
| Serum levels of IL-6 (pg/mL) | 44 | 354.4 | (354.4–1155.1) | 28 | 589.2 | (589.2–1138) | 0.208 |
| Serum levels of IL-17A (pg/mL) | 31 | 180.1 | (180.1–559.5) | 18 | 252.5 | (252.5–637) | 0.272 |
| Serum levels of IL-18 (pg/mL) | 52 | 945.0 | (945–6632.9) | 31 | 1778.9 | (1778.9–7418) | 0.519 |
| Serum levels of TNF-α (pg/mL) | 44 | 143.3 | (143.3–573.8) | 28 | 219.0 | (219–581.3) | 0.529 |
| Univariate Analysis | Multivariate Analysis | |||||
|---|---|---|---|---|---|---|
| OR | (95% CI) | p Value | OR | (95% CI) | p Value | |
| Age | 0.99 | (0.97–1.02) | 0.603 | |||
| Gender | ||||||
| Male | ||||||
| Female | 1.00 | (0.40–2.53) | 0.997 | |||
| ATG16L1 | ||||||
| AGCTCT + GGTTCC | ref. | ref. | ||||
| AACCTT | 2.87 | (1.08–7.69) | 0.035 * | 3.25 | (1.15–9.14) | 0.026 * |
| Rash | 1.07 | (0.35–3.22) | 0.907 | |||
| Arthritis | 0.00 | (0.00– | 0.997 | |||
| Sore throat | 1.87 | (0.74–4.75) | 0.188 | |||
| Lymphadenopathy | 2.32 | (0.94–5.74) | 0.068 | |||
| hepatospenomegaly | 2.25 | (0.48–10.53) | 0.303 | |||
| Leukocytosis | 1.24 | (0.46–3.31) | 0.667 | |||
| Liver dysfunction | 2.76 | (1.03–7.38) | 0.043 * | |||
| Disease activity score | 1.08 | (0.76–1.54) | 0.662 | |||
| ALT | 1.03 | (1.01–1.05) | 0.013 * | 1.03 | (1.00–1.05) | 0.018 * |
| WBC | 1.00 | (1.00–1.00) | 0.718 | |||
| Hemoglobin | 0.90 | (0.70–1.16) | 0.427 | |||
| Platelet | 1.00 | (1.00–1.00) | 0.342 | |||
| ESR | 1.00 | (0.99–1.01) | 0.747 | |||
| CRP | 1.07 | (1.00–1.15) | 0.046 * | 1.06 | (0.98–1.15) | 0.119 |
| Ferritin | 1.00 | (1.00–1.00) | 0.070 | |||
| Serum levels of IL-1 (pg/mL) | 1.00 | (0.97–1.04) | 0.957 | |||
| Serum levels of IL-6 (pg/mL) | 1.00 | (1.00–1.00) | 0.876 | |||
| Serum levels of IL-17A (pg/mL) | 1.00 | (1.00–1.00) | 0.050 | |||
| Serum levels of IL-18 (pg/mL) | 1.0003 | (1.00–1.001) | 0.040 * | |||
| Serum levels of TNF-α (pg/mL) | 1.00 | (1.00–1.00) | 0.307 | |||
| LC3-II mRNA | 1.01 | (0.98–1.05) | 0.402 | |||
| ATG16L1 mRNA | 1.00 | (1.00–1.00) | 0.760 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, W.-T.; Hung, S.-I.; Chen, Y.-M.; Hsieh, C.-W.; Chen, H.-H.; Tang, K.-T.; Chen, D.-Y.; Lan, T.-H. The Association of ATG16L1 Variations with Clinical Phenotypes of Adult-Onset Still’s Disease. Genes 2021, 12, 904. https://doi.org/10.3390/genes12060904
Hung W-T, Hung S-I, Chen Y-M, Hsieh C-W, Chen H-H, Tang K-T, Chen D-Y, Lan T-H. The Association of ATG16L1 Variations with Clinical Phenotypes of Adult-Onset Still’s Disease. Genes. 2021; 12(6):904. https://doi.org/10.3390/genes12060904
Chicago/Turabian StyleHung, Wei-Ting, Shuen-Iu Hung, Yi-Ming Chen, Chia-Wei Hsieh, Hsin-Hua Chen, Kuo-Tung Tang, Der-Yuan Chen, and Tsuo-Hung Lan. 2021. "The Association of ATG16L1 Variations with Clinical Phenotypes of Adult-Onset Still’s Disease" Genes 12, no. 6: 904. https://doi.org/10.3390/genes12060904
APA StyleHung, W. -T., Hung, S. -I., Chen, Y. -M., Hsieh, C. -W., Chen, H. -H., Tang, K. -T., Chen, D. -Y., & Lan, T. -H. (2021). The Association of ATG16L1 Variations with Clinical Phenotypes of Adult-Onset Still’s Disease. Genes, 12(6), 904. https://doi.org/10.3390/genes12060904