
Genome Size, Chromosome Number and Morphological Data Reveal Unexpected Infraspecific Variability in Festuca (Poaceae)

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Geographical Origin
2.2. Flow Cytometric Analyses
2.3. Chromosome Counts
2.4. Morphometric Analyses
2.5. Statistical Analyses
3. Results
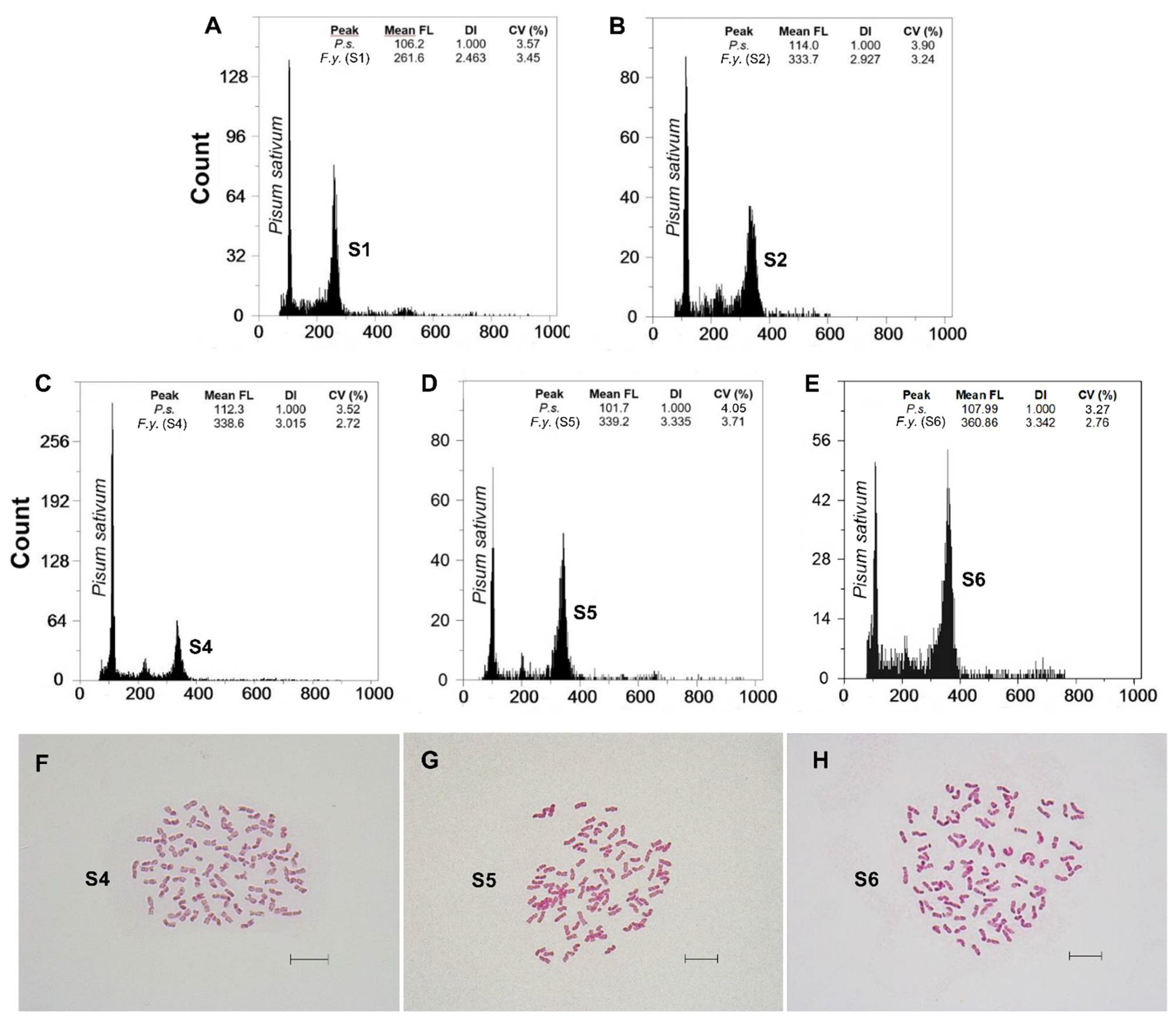
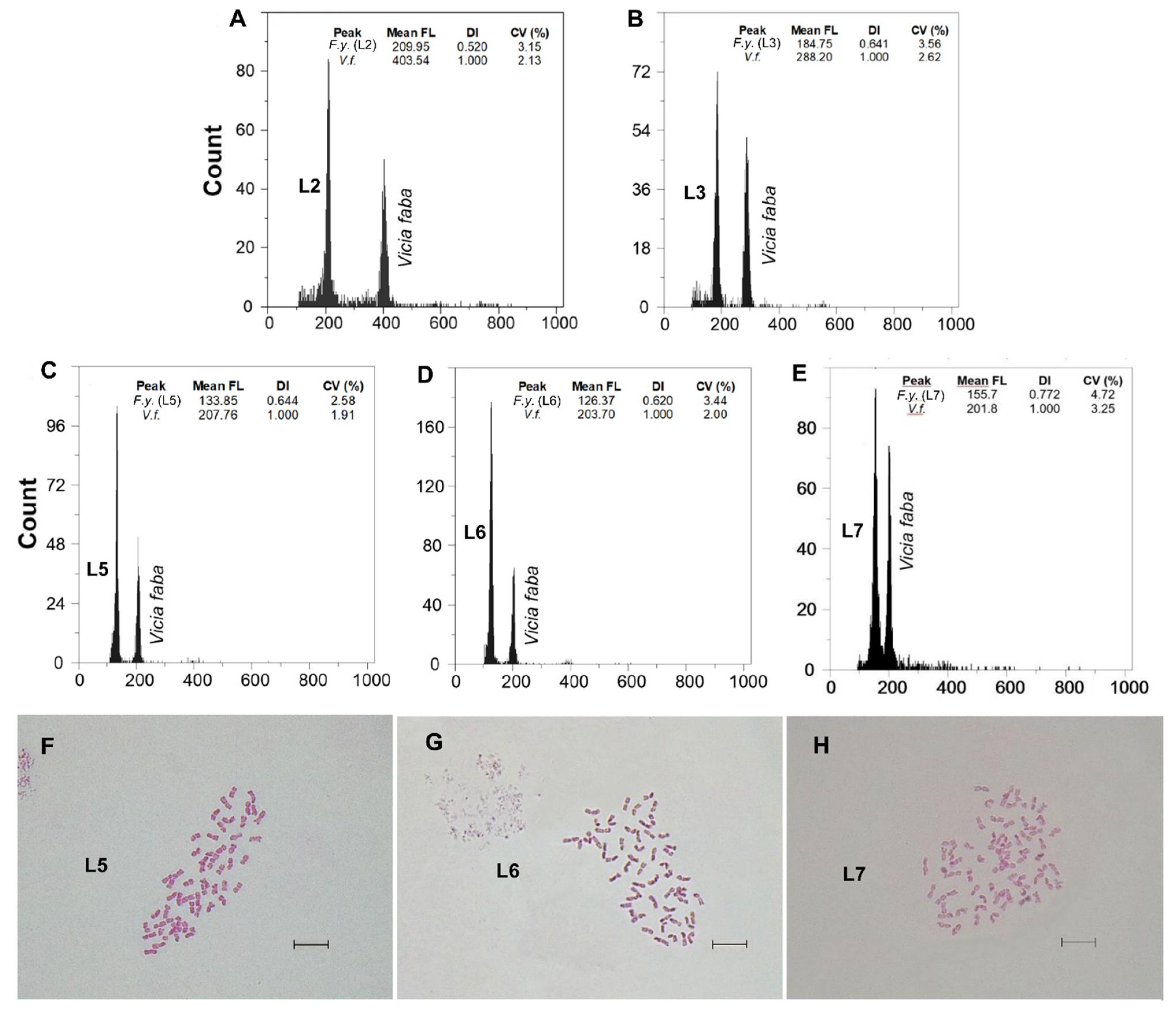
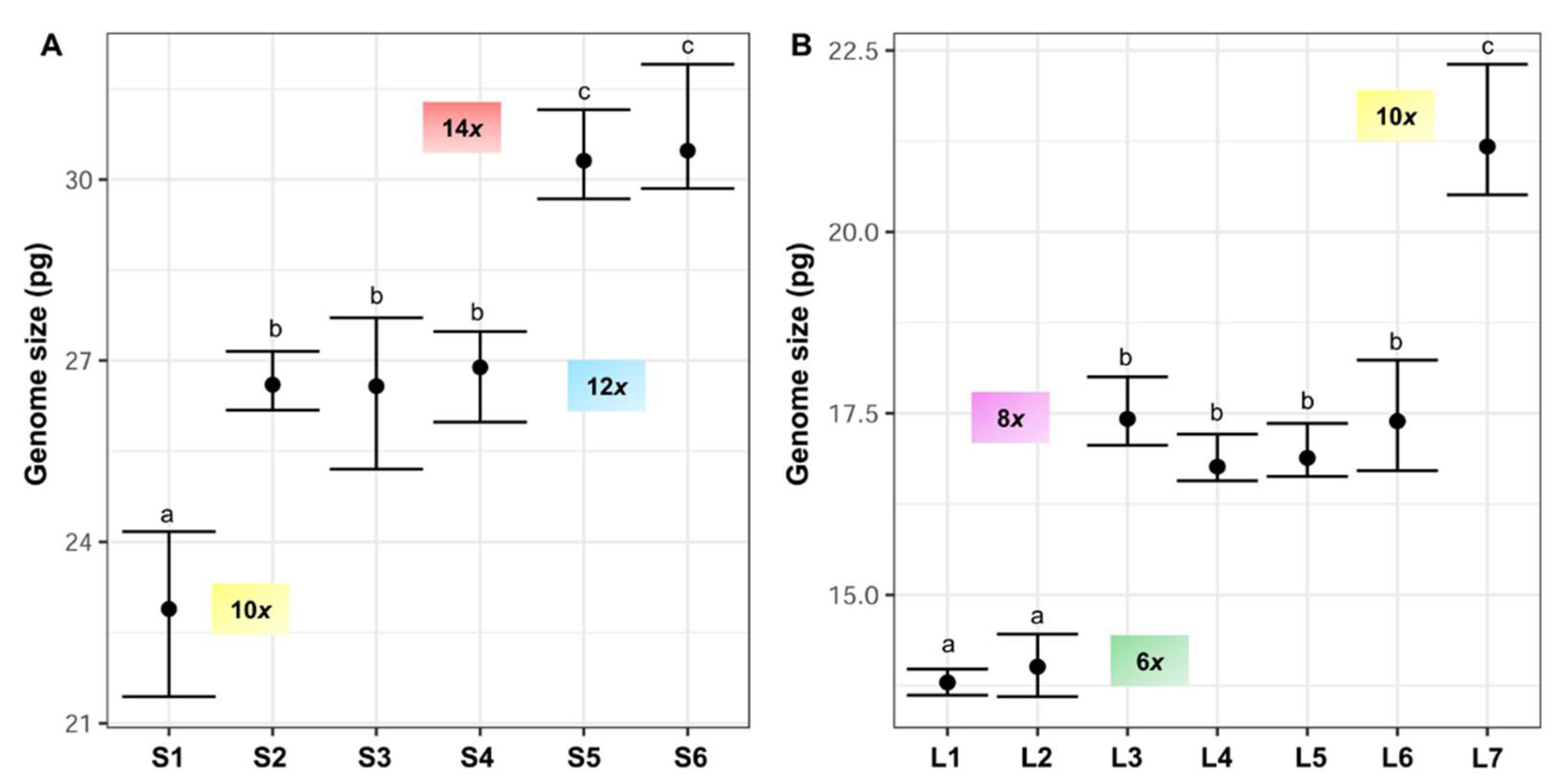
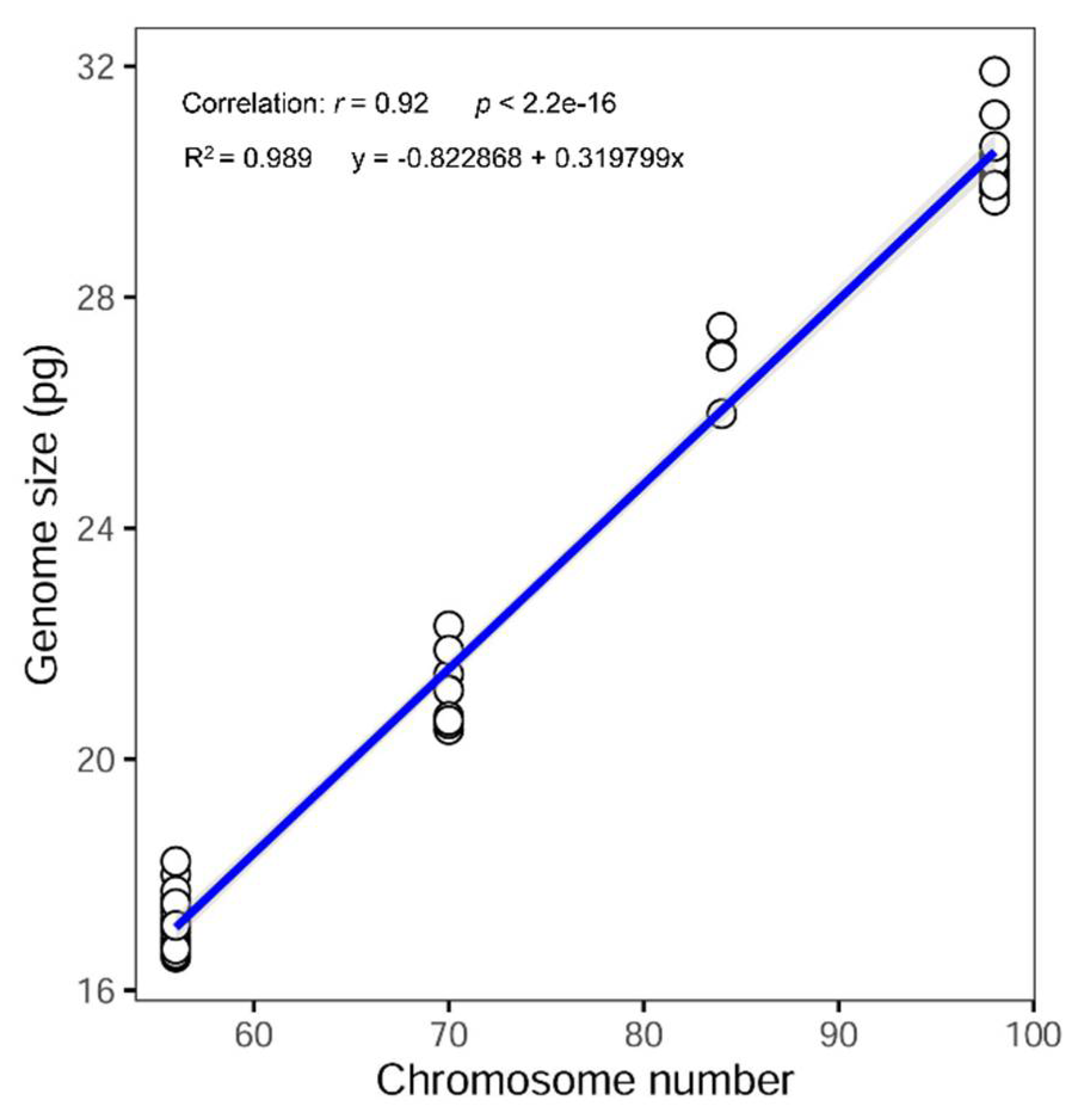
3.1. Genome Size and DNA-Ploidy Level Variation
3.2. Chromosome Numbers
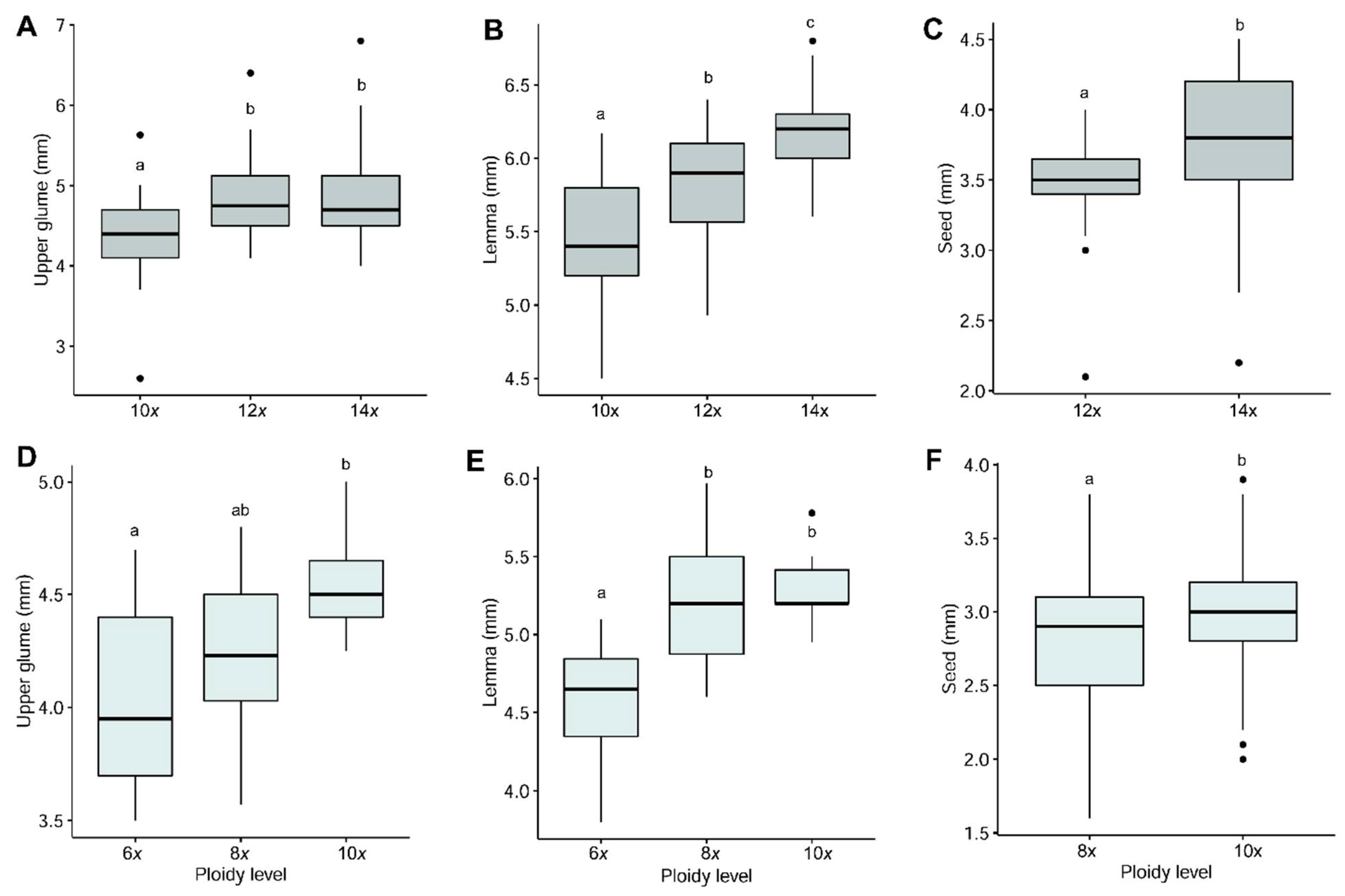
3.3. Morphological Variation
4. Discussion
4.1. Cytogenetic Variation
4.2. Morphological Variation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soltis, D.E.; Soltis, P.S. Polyploidy: Recurrent formation and genome evolution. Trends Ecol. Evol. 1999, 14, 348–352. [Google Scholar] [CrossRef]
- Otto, S.P.; Whitton, J. Polyploidy incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [Green Version]
- Soltis, D.E.; Visger, C.J.; Marchant, D.B.; Soltis, P.S. Polyploidy: Pitfalls and paths to a paradigm. Am. J. Bot. 2016, 103, 1146–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971; pp. 1–216. [Google Scholar]
- Beaulieu, J.M.; Leitch, I.J.; Patel, S.; Pendharkar, A.; Knight, C.A. Genome size is a strong predictor of cell size and stomatal density in angiosperms. New Phytol. 2008, 179, 975–986. [Google Scholar] [CrossRef] [Green Version]
- Balao, F.; Herrera, J.; Talavera, S. Phenotypic consequences of polyploidy and genome size at the microevolutionary scale: A multivariate morphological approach. New Phytol. 2011, 192, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Rios, E.F.; Kenworthy, K.E.; Munoz, P.R. Association of phenotypic traits with ploidy and genome size in annual ryegrass. Crop Sci. 2015, 55, 2078–2090. [Google Scholar] [CrossRef]
- Levy, A.A.; Feldman, M. The impact of polyploidy on grass genome evolution. Plant Physiol. 2002, 130, 1587–1593. [Google Scholar] [CrossRef] [Green Version]
- Watson, L.; Dallwitz, M.J. The Grass Genera of the World; CAB International: Wallingford, UK, 1992; pp. 1–1038. [Google Scholar]
- Clayton, W.D.; Vorontsova, M.S.; Harman, K.T.; Williamson, H. GrassBase—The Online World Grass Flora 2006 onwards. Available online: http://www.kew.org/data/grasses-db.html (accessed on 14 April 2021).
- Malik, C.P.; Thomas, P.T. Karyotypic studies in some Lolium and Festuca species. Caryologia 1966, 19, 167–196. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, J.; Kopecký, D.; Castro, S.; Santos, C.; Silveira, P. Flow cytometric and cytogenetic analices of Iberian Peninsula Festuca spp. Plant Syst. Evol. 2007, 269, 89–105. [Google Scholar] [CrossRef]
- Devesa, J.A.; Martínez-Sagarra, G.; López Nieto, E.; Muñoz Rodríguez, A.; Cebolla, C.; Ortúñez, E. Festuca L. In Flora Iberica; Devesa, J.A., Romero Zarco, C., Buira, A., Quintanar, A., Aedo, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 2020; Volume 19, pp. 200–373. [Google Scholar]
- Martínez Sagarra, G.; Devesa, J.A. Sobre el nombre Festuca yvesii subsp. graniticola (Kerguélen & Morla) Mart.-Sagarra & Devesa. Acta Bot. Malac. 2021, 46. [Google Scholar] [CrossRef]
- Kerguélen, M.; Morla, C. Festuca graniticola, nueva especie del noroeste de la Península Ibérica. Anales Jard. Bot. Madrid 1985, 42, 155–158. [Google Scholar]
- Fuente, V.; Ortúñez, E. Nueva especie de Festuca, L. sección Festuca (Poaceae) en la Península Ibérica. Lazaroa 2000, 21, 3–6. [Google Scholar]
- Fuente, V.; Ferrero, L.M.; Ortúñez, E. Chromosome counts in the genus Festuca L. section Festuca (Poaceae) in the Iberian Peninsula. Bot. J. Linn. Soc. 2001, 137, 385–398. [Google Scholar] [CrossRef]
- Ortúñez, E.; Fuente, V. Reports (394-400) in Mediterranean chromosome number reports. Fl. Medit. 1995, 5, 261–265. [Google Scholar]
- Kerguélen, M. Les Gramineae (Poaceae) de la flore française. Essai de mise au point taxonomique et nomenclaturale. Lejeuniaser. 2 1975, 75, 1–343. [Google Scholar]
- Fuente, V.; Ortúñez, E. A new species of Festuca L. (Poaceae) from Pyrenees (Spain). Fontqueria 1994, 40, 35–42. [Google Scholar]
- Suda, J.; Krahulcová, A.; Trávníek, P.; Krahulec, F. Ploidy level versus DNA ploidy level: An appeal for consistent terminology. Taxon 2006, 55, 447–450. [Google Scholar] [CrossRef]
- Pellicer, J.; Leitch, I.J. The application of flow cytometry for estimating genome size and ploidy level in plants. Methods Mol. Biol. 2014, 1115, 279–307. [Google Scholar]
- Suda, J.; Trávníček, P. Reliable DNA ploidy determination in dehydrated tissues of vascular plants by DAPI flow cytometry—New prospects for plant research. Cytom. Part A 2006, 69, 273–280. [Google Scholar] [CrossRef]
- Šmarda, P. DNA ploidy levels and intraspecific DNA content variability in Romanian fescues (Festuca, Poaceae) measured in fresh and herbarium material. Folia Geobot. 2006, 41, 417–432. [Google Scholar] [CrossRef]
- Šmarda, P.; Stančík, D. Ploidy Level Variability in South American Fescues (Festuca L., Poaceae): Use of flow cytometry in up to 5 1/2-year-old caryopses and herbarium specimens. Plant Biol. 2006, 8, 73–80. [Google Scholar] [CrossRef]
- Viruel, J.; Conejero, M.; Hidalgo, O.; Pokorny, L.; Powell, R.F.; Forest, F.; Kantar, M.B.; Soto Gomez, M.; Graham, S.W.; Gravendeel, B.; et al. A target capture-based method to estimate ploidy from herbarium specimens. Front. Plant Sci. 2019, 10, 937. [Google Scholar] [CrossRef]
- Lysak, M.A.; Doležel, J. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 1998, 51, 123–132. [Google Scholar] [CrossRef]
- Marhold, K.; Kudoh, H.; Pak, J.H.; Watanabe, K.; Španiel, S.; Lihová, J. Cytotype diversity and genome size variation in eastern Asian polyploid Cardamine (Brassicaceae) species. Ann. Bot. 2009, 105, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, A.; Loureiro, J.; Arroyo, J.; Olmedo-Vicente, E.; Castro, S. Cytogenetic diversity in the polyploid complex Linum suffruticosum sl. (Linaceae). Bot. J. Linn. Soc. 2021, 195, 216–232. [Google Scholar] [CrossRef]
- Bureš, P.; Wang, Y.F.; Horová, L.; Suda, J. Genome size variation in Central European species of Cirsium (Compositae) and their natural hybrids. Ann. Bot. 2004, 94, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Mahelka, V.; Suda, J.; Jarolímová, V.; Trávníček, P.; Krahulec, F. Genome size discriminates between closely related taxa Elytrigia repens and E. intermedia (Poaceae: Triticeae) and their hybrid. Folia Geobot. 2005, 40, 367–384. [Google Scholar] [CrossRef]
- Garnatje, T.; Garcia, S.; Vilatersana, R.; Vallès, J. Genome size variation in the genus Carthamnus (Asteraceae, Cardueae): Systematic implications and additive changes during alloploidization. Ann. Bot. 2006, 97, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Suda, J.; Krahulcová, A.; Trávníček, P.; Rosenbaumová, R.; Peckert, T.; Krahulec, F. Genome size variation and species relationships in Hieracium subgenus Pilosella (Asteraceae) as inferred by flow cytometry. Ann. Bot. 2007, 100, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Španiel, S.; Marhold, K.; Hodálová, I.; Lihová, J. Diploid and tetraploid cytotypes of Centaurea stoebe (Asteraceae) in Central Europe: Morphological differentiation and cytotype distribution patterns. Folia Geobot. 2008, 43, 131–158. [Google Scholar] [CrossRef]
- Cires, E.; Cuesta, C.; Peredo, E.L.; Revilla, M.Á.; Fernández Prieto, J.A. Genome size variation and morphological differentiation within Ranunculus parnassifolius group (Ranunculaceae) from calcareous screes in the Northwest of Spain. Plant Syst. Evol. 2009, 281, 193–208. [Google Scholar] [CrossRef]
- Chumová, Z.; Krejčíková, J.; Mandáková, T.; Suda, J.; Trávníček, P. Evolutionary and taxonomic implications of variation in nuclear genome size: Lesson from the grass genus Anthoxanthum (Poaceae). PLoS ONE 2015, 10, e0133748. [Google Scholar] [CrossRef]
- Prančl, J.; Koutecký, P.; Trávníček, P.; Jarolímová, V.; Lučanová, M.; Koutecká, E.; Kaplan, Z. Cytotype variation, cryptic diversity and hybridization in Ranunculus sect. Batrachium revealed by flow cytometry and chromosome numbers. Preslia 2018, 90, 195–223. [Google Scholar] [CrossRef]
- Rewers, M.; Jedrzejczyk, I.; Rewicz, A.; Jakubska-Busse, A. Genome Size Diversity in Rare, Endangered, and Protected Orchids in Poland. Genes 2021, 12, 563. [Google Scholar] [CrossRef] [PubMed]
- Leitch, I.J.; Johnston, E.; Pellicer, J.; Hidalgo, O.; Bennett, M.D. Angiosperm DNA C-Values Database (Release 9.0, April 2019). 2019. Available online: https://cvalues.science.kew.org/ (accessed on 14 April 2021).
- Huff, D.R.; Palazzo, A.J. Fine fescue species determination by laser flow cytometry. Crop Sci. 1998, 38, 445–450. [Google Scholar] [CrossRef]
- Arumuganathan, K.; Tallury, S.P.; Fraser, M.L.; Bruneau, A.H.; Qu, R. Nuclear DNA content of thirteen turfgrass species by flow cytometry. Crop Sci. 1999, 39, 1518–1521. [Google Scholar]
- Šmarda, P.; Bureš, P.; Horová, L.; Foggi, B.; Rossi, G. Genome size and GC content evolution of Festuca: Ancestral expansion and subsequent reduction. Ann. Bot. 2008, 101, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rewicz, A.; Tomczyk, P.P.; Kiedrzyński, M.; Zielińska, K.M.; Jędrzejczyk, I.; Rewers, M.; Kiedrzyńska, E.; Rewicz, T. Morphometric traits in the fine-leaved fescues depend on ploidy level: The case of Festuca amethystina L. PeerJ 2018, 6, e5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Hamernick, S.; Ortiz, J.B.; Watkins, E. DNA content and ploidy estimation of Festuca ovina accessions by flow cytometry. Crop Sci. 2020, 60, 2757–2767. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Doležel, J.; Sgorbati, S.; Lucretii, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plant. 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Snow, R. Alcoholic hydrochloric acid-carmine as stain for chromosomes in squash preparations. Stain Technol. 1963, 38, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Bainard, J.D.; Husband, B.C.; Baldwin, S.J.; Fazekas, A.J.; Gregory, T.R.; Newmaster, S.G.; Kron, P. The effects of rapid desiccation on estimates of plant genome size. Chromosome Res. 2011, 19, 825–842. [Google Scholar] [CrossRef]
- Leitch, I.J.; Bennett, M.D. Genome downsizing in polyploid plants. Biol. J. Linn. Soc. 2004, 82, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Ezquerro-López, D.; Kopecký, D.; Inda, L.Á. Cytogenetic relationships within the Maghrebian clade of Festuca subgen. Schedonorus (Poaceae), using flow cytometry and FISH. Anales Jard. Bot. Madrid 2017, 74, e052. [Google Scholar]
- Bennetzen, J.L.; Ma, J.; Devos, K.M. Mechanism of recent genome size variation in flowering plants. Ann. Bot. 2005, 95, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Šmarda, P.; Bureš, P. Intraspecific DNA content variability in Festuca pallens on different geographical scales and ploidy levels. Ann. Bot. 2006, 98, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Šmarda, P.; Bureš, P.; Horová, L.; Rotreklová, O. Intrapopulation genome size dynamics in Festuca pallens. Ann. Bot. 2008, 102, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Catalán, P. Phylogeny and evolution of Festuca L. and related genera of the subtribe Loliinae (Poeae, Poaceae). In Plant Genome: Biodiversity and Evolution; Sharma, A.K., Sharma, A., Eds.; Science Publishers: London, UK, 2006; Volume 1D, pp. 255–303. [Google Scholar]
- Inda, L.A.; Segarra-Moragues, J.G.; Müller, J.; Peterson, P.M.; Catalán, P. Dated historical biogeography of the temperate Loliinae (Poaceae, Pooideae) grasses in the northern and southern hemispheres. Mol. Phylogenetics Evol. 2008, 46, 932–957. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Sampoux, J.P.; Huyghe, C. Contribution of ploidy-level variation and adaptive trait diversity to the environmental distribution of taxa in the ‘fine-leaved fescue’ line age (genus Festuca subg. Festuca). J. Biogeogr. 2009, 36, 1978–1993. [Google Scholar] [CrossRef]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, J.; Castro, M.; Cerca de Oliveira, J.; Mota, L.; Torices, R. Genome size variation and polyploidy incidence in the alpine flora from Spain. Anales Jard. Bot. Madrid 2013, 70, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Sagarra, G. Estudio Taxonómico de Festuca L. Sect. Festuca (Poaceae) en la Península Ibérica. Ph.D. Thesis, University of Córdoba, Córdoba, Spain, 2018. [Google Scholar]
- Chen, Z.J. Genetic and epigenetic mechanisms for gene expression and phenotypic variation in plant polyploids. Annu. Rev. Plant Biol. 2007, 58, 377–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madlung, A.; Masuelli, R.W.; Watson, B.; Reynolds, S.H.; Davison, J.; Comai, L. Remodeling of DNA methylation and phenotypic and transcriptional changes in synthetic Arabidopsis allotetraploids. Plant Physiol. 2002, 129, 733–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stebbins, G.L. Cytogenetics and evolution of the grass family. Am. J. Bot. 1956, 43, 890–905. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies (According to Devesa et al., [13]) | 2n | Method | References |
|---|---|---|---|
| F. yvesii subsp. yvesii | 2n = 6x = 42 * | Counting 1 | [19] |
| 2n = 8x = 56 | Counting 1 | [17] | |
| F. yvesii subsp. altopyrenaica | 2n = 4x = 28 | Counting 2 | [20] |
| F. yvesii subsp. summilusitana | 2n = 6x = 42 | Counting 3,4 | [17,18] |
| 2n = 10x = 70 | Counting and FCM 4 | [12,17] | |
| 2n = 12x = 84 | Counting and FCM 4 | [12] | |
| F. yvesii subsp. lagascae | 2n = 6x = 42 | Counting 5 | [17] |
| 2n = 8x = 56 | Counting 1 | [17] |
| Taxon | Mountain Range | Locality | n | 2C Mean ± SD (pg) | 2C Range (pg) | Material | DNA-Ploidy Level | 2n |
|---|---|---|---|---|---|---|---|---|
| F. yvesii subsp. summilusitana | W range: Serra da Estrela (Portugal) | S1: Castelo Branco, Manteigas | 9 | 22.89 ± 0.96 | 21.44–24.17 | F | 10x | - |
| S2: Castelo Branco, Sabugueiro | 8 | 26.60 ± 0.35 | 26.18–27.15 | F | 12x | - | ||
| S3: Castelo Branco, Torre | 6 | 26.58 ± 0.87 | 25.20–27.71 | F | 12x | - | ||
| C-W range: Sierra de Gredos and Candelario (Spain) | S4: Ávila, Puerto del Pico | 5 | 26.89 ± 0.55 | 25.98–27.48 | F | 12x | 84 | |
| S5: Ávila, Plataforma de Gredos | 7 | 30.31 ± 0.49 | 29.68–31.16 | F | 14x | 98 | ||
| S6: Salamanca, La Covatilla | 6 | 30.48 ± 0.73 | 29.85–31.91 | F | 14x | 98 | ||
| NW mountains (Spain) | S7: León, Villalibre de Somoza | 3 | 24.67 ± 1.47 | 23.01–25.84 | D | 10x | - | |
| S8: Zamora, Sierra Segundera | 1 | 24.11 | D | 10x | - | |||
| S9: Zamora, Alto de San Juan | 1 | 24.82 | D | 10x | - | |||
| F. yvesii subsp. lagascae | n range: Sierra de la Demanda and Cebollera (Spain) | L1: Logroño, Valdezcaray | 6 | 13.79 ± 0.15 | 13.62–13.98 | F | 6x | - |
| L2: Burgos, Trigaza | 10 | 14.01 ± 0.32 | 13.60–14.46 | F | 6x | - | ||
| L3: Logroño/Soria, Puerto de Piqueras | 10 | 17.42 ± 0.29 | 17.06–18.00 | F | 8x | - | ||
| C range: Sierra de Guadarrama (Spain) | L4: Madrid, Puerto de Guadarrama | 7 | 16.76 ± 0.22 | 16.57–17.21 | F | 8x | 56 | |
| L5: Madrid, Puerto de Navacerrada | 7 | 16.88 ± 0.24 | 16.63–17.36 | F | 8x | 56 | ||
| L6: Madrid, Puerto de la Morcuera | 4 | 17.39 ± 0.65 | 16.71–18.23 | F | 8x | 56 | ||
| L7: Madrid, Puerto de Canencia | 9 | 21.18 ± 0.62 | 20.51–22.31 | F | 10x | 70 | ||
| North range: Cantabrian mountains (Spain) | L8: Puerto de San Glorio | 2 | 19.31 ± 0.41 | 19.02–19.59 | D | 8x | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Sagarra, G.; Castro, S.; Mota, L.; Loureiro, J.; Devesa, J.A. Genome Size, Chromosome Number and Morphological Data Reveal Unexpected Infraspecific Variability in Festuca (Poaceae). Genes 2021, 12, 906. https://doi.org/10.3390/genes12060906
Martínez-Sagarra G, Castro S, Mota L, Loureiro J, Devesa JA. Genome Size, Chromosome Number and Morphological Data Reveal Unexpected Infraspecific Variability in Festuca (Poaceae). Genes. 2021; 12(6):906. https://doi.org/10.3390/genes12060906
Chicago/Turabian StyleMartínez-Sagarra, Gloria, Sílvia Castro, Lucie Mota, João Loureiro, and Juan A. Devesa. 2021. "Genome Size, Chromosome Number and Morphological Data Reveal Unexpected Infraspecific Variability in Festuca (Poaceae)" Genes 12, no. 6: 906. https://doi.org/10.3390/genes12060906
APA StyleMartínez-Sagarra, G., Castro, S., Mota, L., Loureiro, J., & Devesa, J. A. (2021). Genome Size, Chromosome Number and Morphological Data Reveal Unexpected Infraspecific Variability in Festuca (Poaceae). Genes, 12(6), 906. https://doi.org/10.3390/genes12060906