
Towards Forensic DNA Phenotyping for Predicting Visible Traits in Dogs

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Documentation and DNA Extraction
2.2. Marker Selection and Study Design
2.3. PCR Amplification and Sanger Sequencing
3. Results
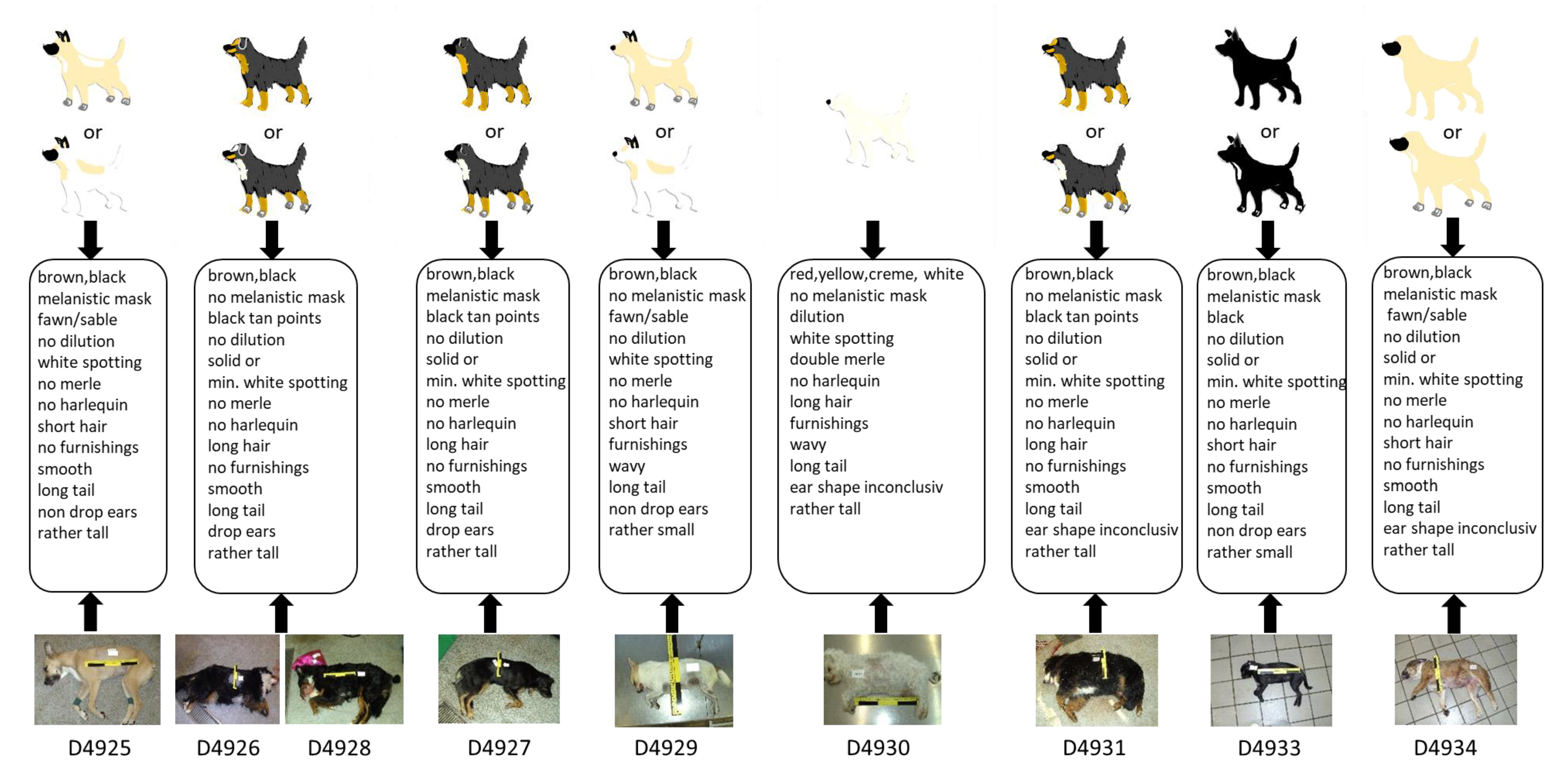
3.1. Pilot Test
3.2. Marker Test
3.2.1. Coat Colour and Coat Pattern
3.2.2. Coat Structure
3.2.3. Tail Length

{kind=link}
{kind=link}
| Trait Category | Marker ID | Locus | Allele | Marker Type | Nucleotide | Dominance (x) | Phenotype | Reference |
|---|---|---|---|---|---|---|---|---|
| Coat colour, coat pattern | MC1R_306ter | E (extension) | e | SNP | T | red, yellow, cream, white | [33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55] | |
| E | C | x | black, brown | |||||
| MC1R_M264V | E (extension) | E | SNP | A | no melanistic mask (black, brown) | [32,34] | ||
| Em | G | x | melanistic mask (black, brown) | |||||
| CBD103_S54 | K (from ‘dominant black’) | ky | INDEL | GGG | yellow—expression of agouti alleles | [28,80] | ||
| KB | DelGGG | x | black, brown, blue | |||||
| CBD103_S53 | K (from ‘dominant black’) | ky | SNP | G | yellow—expression of agouti alleles | [28,80] | ||
| KB | C | x | black, brown, blue | |||||
| ASIP S82 | A (agouti) | Ay | SNP | T | x | fawn/sable | [27] | |
| aw | G | wild type/agouti (black, brown) | ||||||
| ASIP H83 | A (agouti) | Ay | SNP | A | x | fawn/sable | [27] | |
| aw | G | wild type/agouti (black, brown) | ||||||
| ASIP_SINE | A (agouti) | at | SINE | SINE | tan points (black, brown), tricolour | [29] | ||
| aw | no SINE | wild type/agouti (black, brown) | ||||||
| ASIP_R96 | A (agouti) | at | SNP | C | tan points (black, brown), tricolour | [27] | ||
| a | T | recessive black | ||||||
| TYRP1_Q331ter | B (brown) | B | SNP | C | x | black | [33,55] | |
| bs | T | brown | ||||||
| TYRP1_345delP | B (brown) | B | INDEL | CCT | x | black | [33,55] | |
| bd | DelCCT | brown | ||||||
| MLPH_157471_c. -22G>A | D (dilutes eumelanin) | D | SNP | G | x | not diluted pigmentation | [30,56,84] | |
| d | A | diluted pigmentation | ||||||
| MITF_SNP | S (spotting) | S | SNP | A | not clarified | solid colorred, minimal white spotting | [47] | |
| S | G | white spotting | ||||||
| MITF_INS | S (spotting) | S | SINE | no SINE | solid coloured, minimal white spotting | [85] | ||
| Sp | SINE | white spotting (Sp—piebald) | ||||||
| S/Sp | pseudo irish spotting | |||||||
| PMEL | M (merle) | M | SINE | SINE | semi dominant | merle | [39,40,41,46] | |
| m | no SINE | no merle | ||||||
| M/m | SINE/no SINE | mild merle | ||||||
| PSMB7 | H (harlequin) | H | SNP | G | x | harlequin | [35,37] | |
| h | T | no harlequin | ||||||
| Coatcoat structure | FGF5 | n.a. | n.a. | SNP | G | x | short hair | [36] |
| T | long hair | |||||||
| RSPO2 | n.a. | n.a. | INDEL | no Ins | no furnishings | [36] | ||
| Ins | x | furnishings | ||||||
| KRT71 | n.a. | n.a. | SNP | A | semi dominant | smooth coat | [36,40] | |
| G | curly coat | |||||||
| A/G | wavy coat | |||||||
| Tail length | T-Box_C295G | n.a. | n.a. | SNP | G | x | bobtail | [48,54] |
| C | long tail | |||||||
| Ear shape | BICFPJ1062878 | n.a. | n.a. | SNP | G | not clarified | non drop ears | [89] |
| A | drop ears | |||||||
| Body size | IGF1R | n.a. | n.a. | SNP | G | rather tall | [49] | |
| A | x | rather small |
3.2.4. Ear Shape
3.2.5. Body Size
3.3. Blind Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Kayser, M. Forensic DNA Phenotyping: Predicting human appearance from crime scene material for investigative purposes. Forensic Sci. Int. Genet. 2015, 18, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.; de Knijff, P. Improving human forensics through advances in genetics, genomics and molecular biology. Nat. Rev. Genet. 2011, 12, 179–192. [Google Scholar] [CrossRef]
- Grimes, E.A.; Noake, P.J.; Dixon, L.; Urquhart, A. Sequence polymorphism in the human melanocortin 1 receptor gene as an indicator of the red hair phenotype. Forensic Sci. Int. 2001, 122, 124–129. [Google Scholar] [CrossRef]
- Chaitanya, L.; Breslin, K.; Zuniga, S.; Wirken, L.; Pospiech, E.; Kukla-Bartoszek, M.; Sijen, T.; Knijff, P.; Liu, F.; Branicki, W.; et al. The HIrisPlex-S system for eye, hair and skin colour prediction from DNA: Introduction and forensic developmental validation. Forensic Sci. Int. Genet. 2018, 35, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.; Chaitanya, L.; Clarisse, L.; Wirken, L.; Draus-Barini, J.; Kovatsi, L.; Maeda, H.; Ishikawa, T.; Sijen, T.; de Knijff, P.; et al. Developmental validation of the HIrisPlex system: DNA-based eye and hair colour prediction for forensic and anthropological usage. Forensic Sci. Int. Genet. 2014, 9, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Kayser, M. A Practical Guide to the HIrisPlex System: Simultaneous Prediction of Eye and Hair Color from DNA. In Forensic DNA Typing Protocols. Methods Mol Biol. 2016, 1420, 213–231. [Google Scholar] [CrossRef]
- Walsh, S.; Lindenbergh, A.; Zuniga, S.B.; Sijen, T.; de Knijff, P.; Kayser, M.; Ballantyne, K.N. Developmental validation of the IrisPlex system: Determination of blue and brown iris colour for forensic intelligence. Forensic Sci. Int. Genet. 2011, 5, 464–471. [Google Scholar] [CrossRef]
- Walsh, S.; Liu, F.; Ballantyne, K.N.; van Oven, M.; Lao, O.; Kayser, M. IrisPlex: A sensitive DNA tool for accurate prediction of blue and brown eye colour in the absence of ancestry information. Forensic Sci. Int. Genet. 2011, 5, 170–180. [Google Scholar] [CrossRef]
- Schneider, P.M.; Prainsack, B.; Kayser, M. The Use of Forensic DNA Phenotyping in Predicting Appearance and Biogeographic Ancestry. Dtsch. Ärzteblatt Int. 2019, 116, 873–880. [Google Scholar] [CrossRef]
- Ostrander, E.A.; Wayne, R.K.; Freedman, A.H.; Davis, B.W. Demographic history, selection and functional diversity of the canine genome. Nat. Rev. Genet. 2017, 18, 705–720. [Google Scholar] [CrossRef]
- Freedman, A.H.; Gronau, I.; Schweizer, R.M.; Ortega-Del Vecchyo, D.; Han, E.; Silva, P.M.; Galaverni, M.; Fan, Z.; Marx, P.; Lorente-Galdos, B.; et al. Genome sequencing highlights the dynamic early history of dogs. PLoS Genet. 2014, 10, e1004016. [Google Scholar] [CrossRef] [Green Version]
- Freedman, A.H.; Wayne, R.K. Deciphering the Origin of Dogs: From Fossils to Genomes. Annu. Rev. Anim. Biosci. 2017, 5, 281–307. [Google Scholar] [CrossRef]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpre, M.B.; Sablin, M.V.; Lopez-Giraldez, F.; Domingo-Roura, X.; et al. Complete mitochondrial genomes of ancient canids suggest a European origin of domestic dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frantz, L.A.; Mullin, V.E.; Pionnier-Capitan, M.; Lebrasseur, O.; Ollivier, M.; Perri, A.; Linderholm, A.; Mattiangeli, V.; Teasdale, M.D.; Dimopoulos, E.A.; et al. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science 2016, 352, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Perri, A.R.; Feuerborn, T.R.; Frantz, L.A.F.; Larson, G.; Malhi, R.S.; Meltzer, D.J.; Witt, K.E. Dog domestication and the dual dispersal of people and dogs into the Americas. Proc. Natl. Acad. Sci. USA 2021, 118, 6. [Google Scholar] [CrossRef] [PubMed]
- Vilà, C.; Leonard, J.A. Canid Phylogeny and Origin of the Domestic Dog. In The Genetics of the Dog; Ostrander, E.A., Ruvinsky, A., Eds.; CABI: Wallingford, UK, 2012. [Google Scholar]
- Perri, A. A wolf in dog’s clothing: Initial dog domestication and Pleistocene wolf variation. J. Archaeol. Sci. 2016, 68, 1–4. [Google Scholar] [CrossRef]
- Baumann, C.; Pfrengle, S.; Munzel, S.C.; Molak, M.; Feuerborn, T.R.; Breidenstein, A.; Reiter, E.; Albrecht, G.; Kind, C.J.; Verjux, C.; et al. A refined proposal for the origin of dogs: The case study of Gnirshohle, a Magdalenian cave site. Sci. Rep. 2021, 11, 5137. [Google Scholar] [CrossRef]
- Botigue, L.R.; Song, S.; Scheu, A.; Gopalan, S.; Pendleton, A.L.; Oetjens, M.; Taravella, A.M.; Seregely, T.; Zeeb-Lanz, A.; Arbogast, R.M.; et al. Ancient European dog genomes reveal continuity since the Early Neolithic. Nat. Commun. 2017, 8, 16082. [Google Scholar] [CrossRef]
- Bergstrom, A.; Frantz, L.; Schmidt, R.; Ersmark, E.; Lebrasseur, O.; Girdland-Flink, L.; Lin, A.T.; Stora, J.; Sjogren, K.G.; Anthony, D.; et al. Origins and genetic legacy of prehistoric dogs. Science 2020, 370, 557–564. [Google Scholar] [CrossRef]
- Skoglund, P.; Ersmark, E.; Palkopoulou, E.; Dalen, L. Ancient wolf genome reveals an early divergence of domestic dog ancestors and admixture into high-latitude breeds. Curr. Biol. 2015, 25, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.G.; Gilbert, S.F. From caveman companion to medical innovator: Genomic insights into the origin and evolution of domestic dogs. Adv. Genom. Genet. 2015, 5, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Moody, J.A.; Clark, L.A.; Murphy, K.E. Working Dogs: History and Applications. In The Dog and Its Genome; Ostrander, E., Giger, U., Lindblad-Toh, E., Eds.; Cold Springer Harbor Laboratory Press: New York, NY, USA, 2006; pp. 1–18. [Google Scholar]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., 3rd; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Parker, H.G. Genomic analyses of modern dog breeds. Mamm. Genome 2012, 23, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic Analyses Reveal the Influence of Geographic Origin, Migration, and Hybridization on Modern Dog Breed Development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef]
- Berryere, T.G.; Kerns, J.A.; Barsh, G.S.; Schmutz, S.M. Association of an Agouti allele with fawn or sable coat color in domestic dogs. Mamm. Genome 2005, 16, 262–272. [Google Scholar] [CrossRef]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A β-defensin mutation causes black coat color in domestic dogs. Science 2007, 318, 1418–1423. [Google Scholar] [CrossRef] [Green Version]
- Dreger, D.L.; Schmutz, S.M. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. J. Hered. 2011, 102, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Philipp, U.; Hamann, H.; Mecklenburg, L.; Nishino, S.; Mignot, E.; Gunzel-Apel, A.R.; Schmutz, S.M.; Leeb, T. Polymorphisms within the canine MLPH gene are associated with dilute coat color in dogs. BMC Genet. 2005, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmutz, S.M.; Berryere, T.G. The genetics of cream coat color in dogs. J. Hered. 2007, 98, 544–548. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, S.M.; Berryere, T.G.; Ellinwood, N.M.; Kerns, J.A.; Barsh, G.S. MC1R studies in dogs with melanistic mask or brindle patterns. J. Hered. 2003, 94, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmutz, S.M.; Berryere, T.G.; Goldfinch, A.D. TYRP1 and MC1R genotypes and their effects on coat color in dogs. Mamm. Genome 2002, 13, 380–387. [Google Scholar] [CrossRef]
- Newton, J.M.; Wilkie, A.L.; He, L.; Jordan, S.A.; Metallinos, D.L.; Holmes, N.G.; Jackson, I.J.; Barsh, G.S. Melanocortin 1 receptor variation in the domestic dog. Mamm. Genome 2000, 11, 24–30. [Google Scholar] [CrossRef]
- Clark, L.A.; Starr, A.N.; Tsai, K.L.; Murphy, K.E. Genome-wide linkage scan localizes the harlequin locus in the Great Dane to chromosome 9. Gene 2008, 418, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Cadieu, E.; Neff, M.W.; Quignon, P.; Walsh, K.; Chase, K.; Parker, H.G.; Vonholdt, B.M.; Rhue, A.; Boyko, A.; Byers, A.; et al. coat variation in the domestic dog is governed by variants in three genes. Science 2009, 326, 150–153. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.A.; Tsai, K.L.; Starr, A.N.; Nowend, K.L.; Murphy, K.E. A missense mutation in the 20S proteasome beta2 subunit of Great Danes having harlequin coat patterning. Genomics 2011, 97, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Kaelin, C.B.; Barsh, G.S. Genetics of pigmentation in dogs and cats. Annu. Rev. Anim. Biosci. 2013, 1, 125–156. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.C.; Evans, J.M.; Tsai, K.L.; Clarke, L.A. Length variations within the Merle retrotransposon of canine PMEL: Correlating genotype with phenotype. Mob. DNA 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Saif, R.; Iftekhar, A.; Asif, F.; Alghanem, M.S. Dog Coat Colour Genetics: A Review. Adv. Life Sci. 2020, 7, 215–224. [Google Scholar]
- Varga, L.; Lenart, X.; Zenke, P.; Orban, L.; Hudak, P.; Ninausz, N.; Pelles, Z.; Szoke, A. Being Merle: The Molecular Genetic Background of the Canine Merle Mutation. Genes 2020, 11, 660. [Google Scholar] [CrossRef] [PubMed]
- Vaysse, A.; Ratnakumar, A.; Derrien, T.; Axelsson, E.; Rosengren Pielberg, G.; Sigurdsson, S.; Fall, T.; Seppala, E.H.; Hansen, M.S.; Lawley, C.T.; et al. Identification of genomic regions associated with phenotypic variation between dog breeds using selection mapping. PLoS Genet. 2011, 7, e1002316. [Google Scholar] [CrossRef] [Green Version]
- Boyko, A.R. The domestic dog: Man’s best friend in the genomic era. Genome Biol. 2011, 12, 216. [Google Scholar] [CrossRef]
- Boyko, A.R.; Quignon, P.; Li, L.; Schoenebeck, J.J.; Degenhardt, J.D.; Lohmueller, K.E.; Zhao, K.; Brisbin, A.; Parker, H.G.; vonHoldt, B.M.; et al. A simple genetic architecture underlies morphological variation in dogs. PLoS Biol. 2010, 8, e1000451. [Google Scholar] [CrossRef] [Green Version]
- Shearin, A.L.; Ostrander, E.A. Canine morphology: Hunting for genes and tracking mutations. PLoS Biol. 2010, 8, e1000310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, L.A.; Wahl, J.M.; Rees, C.A.; Murphy, K.E. Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog. Proc. Natl. Acad. Sci. USA 2006, 103, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Leegwater, P.A.; van Hagen, M.A.; van Oost, B.A. Localization of white spotting locus in boxer dogs on CFA20 by genome-wide linkage analysis with 1500 SNPs. J. Hered. 2007, 98, 549–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hytonen, M.K.; Grall, A.; Hedan, B.; Dreano, S.; Seguin, S.J.; Delattre, D.; Thomas, A.; Galibert, F.; Paulin, L.; Lohi, H.; et al. Ancestral T-Box Mutation Is Present in Many, but Not All, Short-Tailed Dog Breeds. J. Hered. 2009, 100, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Hoopes, B.C.; Rimbault, M.; Liebers, D.; Ostrander, E.A.; Sutter, N.B. The insulin-like growth factor 1 receptor (IGF1R) contributes to reduced size in dogs. Mamm. Genome 2012, 23, 780–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, E.K.; Baranowska, I.; Wade, C.M.; Salmon Hillbertz, N.H.C.; Zody, M.C.; Anderson, N.; Biagi, T.M.; Patterson, N.; Pielberg, G.R.; Kulbokas, E.J.; et al. Efficient mapping of mendelian traits in dogs through genome-wide association. Nat. Genet. 2007, 39, 1321–1328. [Google Scholar] [CrossRef]
- Thomas, R.; Duke, S.E.; Karlsson, E.K.; Evans, A.; Ellis, P.; Lindblad-Toh, K.; Langford, C.F.; Breen, M. A genome assembly-integrated dog 1 Mb BAC microarray: A cytogenetic resource for canine cancer studies and comparative genomic analysis. Cytogenet. Genome Res. 2008, 122, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Sutter, N.B.; Bustamante, C.D.; Chase, K.; Gray, M.M.; Zhao, K.Y.; Zhu, L.; Padhukasahasram, B.; Karlins, E.; Davis, S.; Jones, P.G.; et al. A single IGF1 allele is a major determinant of small size in dogs. Science 2007, 316, 112–115. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An Expressed Fgf4 Retrogene Is Associated with Breed-Defining Chondrodysplasia in Domestic Dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef] [Green Version]
- Haworth, K.; Putt, W.; Cattanach, B.; Breen, M.; Binns, M.; Lingaas, F.; Edwards, Y.H. Canine homolog of the T-box transcription factor T; failure of the protein to bind to its DNA target leads to a short-tail phenotype. Mamm. Genome 2001, 12, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G. Genes affecting coat colour and pattern in domestic dogs: A review. Anim. Genet. 2007, 38, 539–549. [Google Scholar] [CrossRef]
- Welle, M.; Philipp, U.; Rufenacht, S.; Roosje, P.; Scharfenstein, M.; Schutz, E.; Brenig, B.; Linek, M.; Mecklenburg, L.; Grest, P.; et al. MLPH Genotype-Melanin Phenotype Correlation in Dilute Dogs. J. Hered. 2009, 100, S75–S79. [Google Scholar] [CrossRef] [Green Version]
- Sykes, N.; Beirne, P.; Horowitz, A.; Jones, I.; Kalof, L.; Karlsson, E.; King, T.; Litwak, H.; McDonald, R.A.; Murphy, L.J.; et al. Humanity’s Best Friend: A Dog-Centric Approach to Addressing Global Challenges. Animals 2020, 10, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Sarenbo, S.; Svensson, P.A. Bitten or struck by dog: A rising number of fatalities in Europe, 1995–2016. Forensic Sci. Int. 2021, 318, 110592. [Google Scholar] [CrossRef]
- Berger, B.; Berger, C.; Hecht, W.; Hellmann, A.; Rohleder, U.; Schleenbecker, U.; Parson, W. Validation of two canine STR multiplex-assays following the ISFG recommendations for non-human DNA analysis. Forensic Sci. Int. Genet. 2014, 8, 90–100. [Google Scholar] [CrossRef]
- Berger, B.; Berger, C.; Heinrich, J.; Niederstätter, H.; Hecht, W.; Hellmann, A.; Rohleder, U.; Schleenbecker, U.; Morf, N.; Freire-Aradas, A.; et al. Dog breed affiliation with a forensically validated canine STR set. Forensic Sci. Int. Genet. 2018, 37, 126–134. [Google Scholar] [CrossRef]
- Eichmann, C.; Berger, B.; Parson, W. A proposed nomenclature for 15 canine-specific polymorphic STR loci for forensic purposes. Int. J. Leg. Med. 2004, 118, 249–266. [Google Scholar] [CrossRef]
- Wictum, E.; Kun, T.; Lindquist, C.; Malvick, J.; Vankan, D.; Sacks, B. Developmental validation of DogFiler, a novel multiplex for canine DNA profiling in forensic casework. Forensic Sci. Int. Genet. 2013, 7, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Van Asch, B.; Pereira, F. State-of-the-Art and Future Prospects of Canine STR-Based Genotyping. Open Forensic Sci. J. 2010, 3, 45–52. [Google Scholar] [CrossRef]
- Kanthaswamy, S.; Oldt, R.F.; Montes, M.; Falak, A. Comparing two commercial domestic dog (Canis familiaris) STR genotyping kits for forensic identity calculations in a mixed-breed dog population sample. Anim. Genet. 2019, 50, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Kanthaswamy, S.; Tom, B.K.; Mattila, A.-M.; Johnston, E.; Dayton, M.; Kinaga, J.; Erickson, B.J.-A.; Halverson, J.; Fantin, D.; DeNise, S.; et al. Canine population data generated from a multiplex STR kit for use in forensic casework. J. Forensic Sci. 2009, 54, 829–840. [Google Scholar] [CrossRef]
- Hellmann, A.P.; Rohleder, U.; Eichmann, C.; Pfeiffer, I.; Parson, W.; Schleenbecker, U. A Proposal for Standardization in Forensic Canine DNA Typing: Allele Nomenclature of Six Canine-Specific STR Loci. J. Forensic Sci. 2006, 51, 274–281. [Google Scholar] [CrossRef]
- Van Asch, B.; Alves, C.; Gusmao, L.; Pereira, V.; Pereira, F.; Amorim, A. A new autosomal STR nineplex for canine identification and parentage testing. Electrophoresis 2009, 30, 417–423. [Google Scholar] [CrossRef]
- Budowle, B.; Garofano, P.; Hellman, A.; Ketchum, M.; Kanthaswamy, S.; Parson, W.; van Haeringen, W.; Fain, S.; Broad, T. Recommendations for animal DNA forensic and identity testing. Int. J. Leg. Med. 2005, 119, 295–302. [Google Scholar] [CrossRef]
- Linacre, A.; Gusmao, L.; Hecht, W.; Hellmann, A.P.; Mayr, W.R.; Parson, W.; Prinz, M.; Schneider, P.M.; Morling, N. ISFG: Recommendations regarding the use of non-human (animal) DNA in forensic genetic investigations. Forensic Sci. Int. Genet. 2011, 5, 501–505. [Google Scholar] [CrossRef]
- Safer, M.A.; Murphy, R.P.; Wise, R.A.; Bussey, L.; Millett, C.; Holfeld, B. Educating jurors about eyewitness testimony in criminal cases with circumstantial and forensic evidence. Int. J. Law Psychiatry 2016, 47, 86–92. [Google Scholar] [CrossRef]
- Albright, T.D. Why eyewitnesses fail. Proc. Natl. Acad. Sci. USA 2017, 114, 7758–7764. [Google Scholar] [CrossRef] [Green Version]
- Wise, R.A.; Kehn, A. Can the effectiveness of eyewitness expert testimony be improved? Psychiatry Psychol. Law 2020, 27, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Wixted, J.T.; Wells, G.L. The Relationship between Eyewitness Confidence and Identification Accuracy: A New Synthesis. Psychol. Sci. Public Interest 2017, 18, 10–65. [Google Scholar] [CrossRef]
- Berger, B.; Heinrich, J.; Niederstatter, H.; Hecht, W.; Morf, N.; Hellmann, A.; Rohleder, U.; Schleenbecker, U.; Berger, C.; Parson, W.; et al. Forensic characterization and statistical considerations of the CaDNAP 13-STR panel in 1184 domestic dogs from Germany, Austria, and Switzerland. Forensic Sci. Int. Genet. 2019, 42, 90–98. [Google Scholar] [CrossRef]
- Evans, J.J.; Wictum, E.J.; Penedo, M.C.; Kanthaswamy, S. Real-time polymerase chain reaction quantification of canine DNA. J. Forensic Sci. 2007, 52, 93–96. [Google Scholar] [CrossRef]
- Little, C.C. The Inheritance of Coat Color in Dogs; Comstock: Ithaca, NY, USA, 1957; Volume 118, p. 572. [Google Scholar]
- Schmutz, S.M.; Melekhovets, Y. Coat color DNA testing in dogs: Theory meets practice. Mol. Cell. Probes 2012, 26, 238–242. [Google Scholar] [CrossRef]
- Kaelin, C.B.; Barsh, G.S. Molecular genetics of coat colour, texture and length in the dog. In The Genetics of the Dog; Ostrander, E.A., Ruvinsky, A., Eds.; CABI: Wallingford, UK, 2012; pp. 57–82. [Google Scholar]
- Kerns, J.A.; Cargill, E.J.; Clark, L.A.; Candille, S.I.; Berryere, T.G.; Olivier, M.; Lust, G.; Todhunter, R.J.; Schmutz, S.M.; Murphy, K.E.; et al. Linkage and segregation analysis of black and brindle coat color in domestic dogs. Genetics 2007, 176, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Kerns, J.A.; Newton, J.; Berryere, T.G.; Rubin, E.M.; Cheng, J.F.; Schmutz, S.M.; Barsh, G.S. Characterization of the dog Agouti gene and a nonagoutimutation in German Shepherd Dogs. Mamm. Genome 2004, 15, 798–808. [Google Scholar] [CrossRef]
- Dreger, D.L.; Hooser, B.N.; Hughes, A.M.; Ganesan, B.; Donner, J.; Anderson, H.; Holtvoigt, L.; Ekenstedt, K.J. True Colors: Commercially-acquired morphological genotypes reveal hidden allele variation among dog breeds, informing both trait ancestry and breed potential. PLoS ONE 2019, 14, e0223995. [Google Scholar]
- Schmutz, S.M.; Berryere, T.G.; Barta, J.L.; Reddick, K.D.; Schmutz, J.K. Agouti sequence polymorphisms in coyotes, wolves and dogs suggest hybridization. J. Hered. 2007, 98, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Philipp, U.; Quignon, P.; Scott, A.; Andre, C.; Breen, M.; Leeb, T. Chromosomal assignment of the canine melanophilin gene (MLPH): A candidate gene for coat color dilution in Pinschers. J. Hered. 2005, 96, 774–776. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, S.M.; Berryere, T.G.; Dreger, D.L. MITF and White Spotting in Dogs: A Population Study. J. Hered. 2009, 100, S66–S74. [Google Scholar] [CrossRef] [Green Version]
- Langevin, M.; Synkova, H.; Jancuskova, T.; Pekova, S. Merle phenotypes in dogs-SILV SINE insertions from Mc to Mh. PLoS ONE 2018, 13, e0198536. [Google Scholar]
- Parker, H.G.; Shearin, A.L.; Ostrander, E.A. Man’s best friend becomes biology’s best in show: Genome analyses in the domestic dog. Annu. Rev. Genet. 2010, 44, 309–336. [Google Scholar] [CrossRef] [Green Version]
- Indrebo, A.; Langeland, M.; Juul, H.M.; Skogmo, H.K.; Rengmark, A.H.; Lingaas, F. A study of inherited short tail and taillessness in Pembroke Welsh corgi. J. Small Anim. Pract. 2008, 49, 220–224. [Google Scholar] [CrossRef]
- Jones, P.; Chase, K.; Martin, A.; Davern, P.; Ostrander, E.A.; Lark, K.G. Single-nucleotide-polymorphism-based association mapping of dog stereotypes. Genetics 2008, 179, 1033–1044. [Google Scholar] [CrossRef] [Green Version]
- Bannasch, D.; Young, A.; Myers, J.; Truve, K.; Dickinson, P.; Gregg, J.; Davis, R.; Bongcam-Rudloff, E.; Webster, M.T.; Lindblad-Toh, K.; et al. Localization of canine brachycephaly using an across breed mapping approach. PLoS ONE 2010, 5, e9632. [Google Scholar] [CrossRef] [Green Version]
- Schoenebeck, J.J.; Hutchinson, S.A.; Byers, A.; Beale, H.C.; Carrington, B.; Faden, D.L.; Rimbault, M.; Decker, B.; Kidd, J.M.; Sood, R.; et al. Variation of BMP3 Contributes to Dog Breed Skull Diversity. PLoS Genet. 2012, 8, e1002849. [Google Scholar] [CrossRef] [Green Version]
- Schoenebeck, J.J.; Ostrander, E.A. The genetics of canine skull shape variation. Genetics 2013, 193, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Rimbault, M.; Ostrander, E.A. So many doggone traits: Mapping genetics of multiple phenotypes in the domestic dog. Hum. Mol. Genet. 2012, 21, R52–R57. [Google Scholar] [CrossRef] [Green Version]
- Dreger, D.L.; Parker, H.G.; Ostrander, E.A.; Schmutz, S.M. Identification of a Mutation that Is Associated with the Saddle Tan and Black-and-Tan Phenotypes in Basset Hounds and Pembroke Welsh Corgis. J. Hered. 2013, 104, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wallerman, O.; Arendt, M.L.; Sundstrom, E.; Karlsson, A.; Nordin, J.; Makelainen, S.; Pielberg, G.R.; Hanson, J.; Ohlsson, A.; et al. A novel canine reference genome resolves genomic architecture and uncovers transcript complexity. Commun. Biol. 2021, 4, 185. [Google Scholar] [CrossRef]
- Hoeppner, M.P.; Lundquist, A.; Pirun, M.; Meadows, J.R.; Zamani, N.; Johnson, J.; Sundstrom, G.; Cook, A.; FitzGerald, M.G.; Swofford, R.; et al. An improved canine genome and a comprehensive catalogue of coding genes and non-coding transcripts. PLoS ONE 2014, 9, e91172. [Google Scholar] [CrossRef] [PubMed]
- Schoenebeck, J.J.; Ostrander, E.A. Insights into morphology and disease from the dog genome project. Annu. Rev. Cell. Dev. Biol. 2014, 30, 535–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donner, J.; Anderson, H.; Davison, S.; Hughes, A.M.; Bouirmane, J.; Lindqvist, J.; Lytle, K.M.; Ganesan, B.; Ottka, C.; Ruotanen, P.; et al. Frequency and distribution of 152 genetic disease variants in over 100,000 mixed breed and purebred dogs. PLoS Genet. 2018, 14, e1007361. [Google Scholar] [CrossRef]
- Anderson, H.; Honkanen, L.; Ruotanen, P.; Mathlin, J.; Donner, J. Comprehensive genetic testing combined with citizen science reveals a recently characterized ancient MC1R mutation associated with partial recessive red phenotypes in dog. Canine Med. Genet. 2020, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Grubwieser, P.; Muhlmann, R.; Berger, B.; Niederstatter, H.; Pavlic, M.; Parson, W. A new “miniSTR-multiplex” displaying reduced amplicon lengths for the analysis of degraded DNA. Int. J. Leg. Med. 2006, 120, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Grubwieser, P.; Muhlmann, R.; Parson, W. New sensitive amplification primers for the STR locus D2S1338 for degraded casework DNA. Int. J. Leg. Med. 2003, 117, 185–188. [Google Scholar] [CrossRef]
- Hughes-Stamm, S.R.; Ashton, K.J.; van Daal, A. Assessment of DNA degradation and the genotyping success of highly degraded samples. Int. J. Leg. Med. 2011, 125, 341–348. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, C.; Heinrich, J.; Berger, B.; Hecht, W.; Parson, W.; on behalf of CaDNAP. Towards Forensic DNA Phenotyping for Predicting Visible Traits in Dogs. Genes 2021, 12, 908. https://doi.org/10.3390/genes12060908
Berger C, Heinrich J, Berger B, Hecht W, Parson W, on behalf of CaDNAP. Towards Forensic DNA Phenotyping for Predicting Visible Traits in Dogs. Genes. 2021; 12(6):908. https://doi.org/10.3390/genes12060908
Chicago/Turabian StyleBerger, Cordula, Josephin Heinrich, Burkhard Berger, Werner Hecht, Walther Parson, and on behalf of CaDNAP. 2021. "Towards Forensic DNA Phenotyping for Predicting Visible Traits in Dogs" Genes 12, no. 6: 908. https://doi.org/10.3390/genes12060908
APA StyleBerger, C., Heinrich, J., Berger, B., Hecht, W., Parson, W., & on behalf of CaDNAP. (2021). Towards Forensic DNA Phenotyping for Predicting Visible Traits in Dogs. Genes, 12(6), 908. https://doi.org/10.3390/genes12060908