
Genetic History of the Remnant Population of the Rare Orchid Cypripedium calceolus Based on Plastid and Nuclear rDNA



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Sampling
2.3. Molecular Analyses
2.4. Isolation of DNA
2.5. Amplification and Sequencing
2.6. Haplotype Analyses
2.7. Molecular Clock Analyses
3. Results
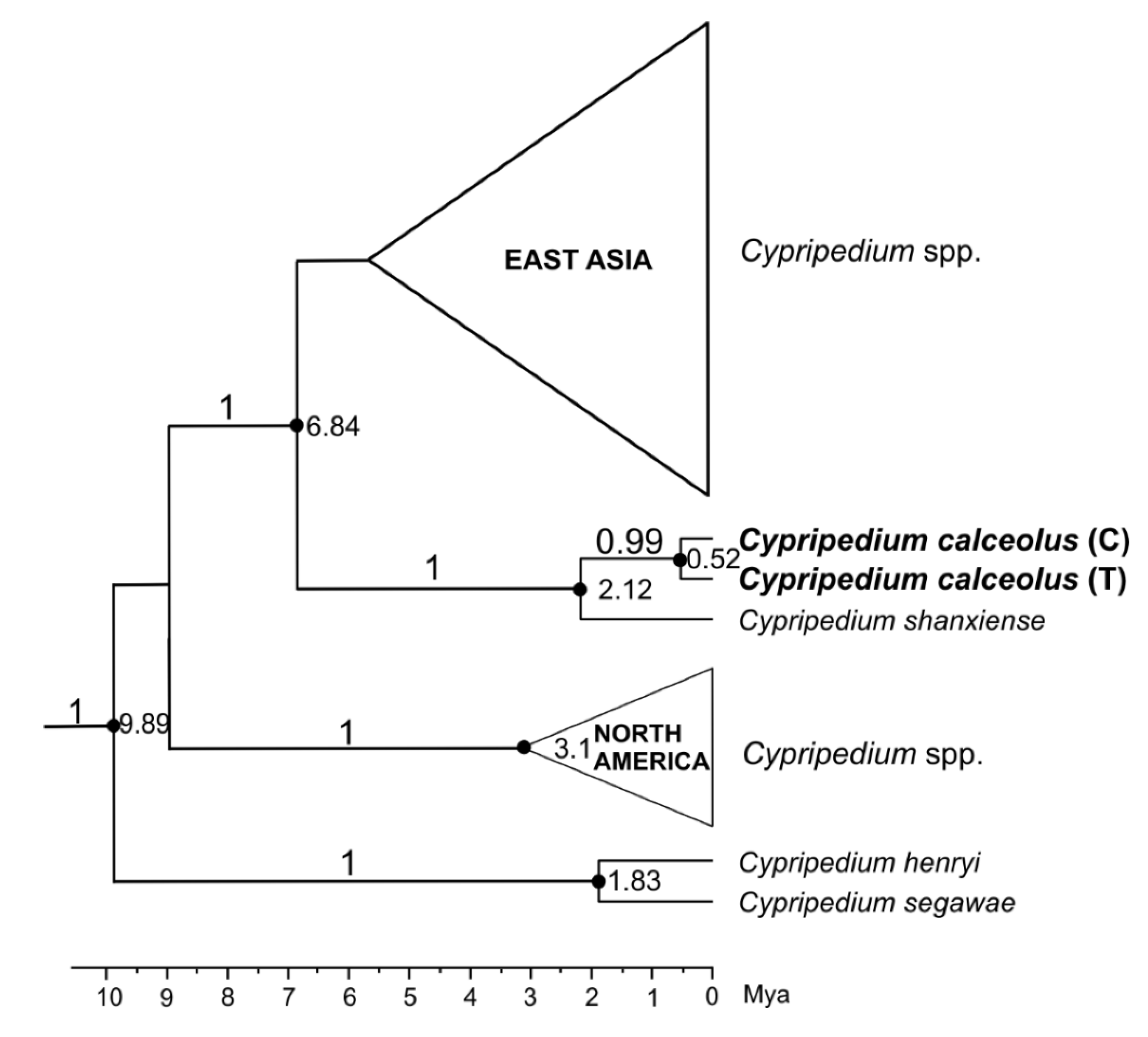
3.1. ITS Ribotypes/Molecular Clock Analysis
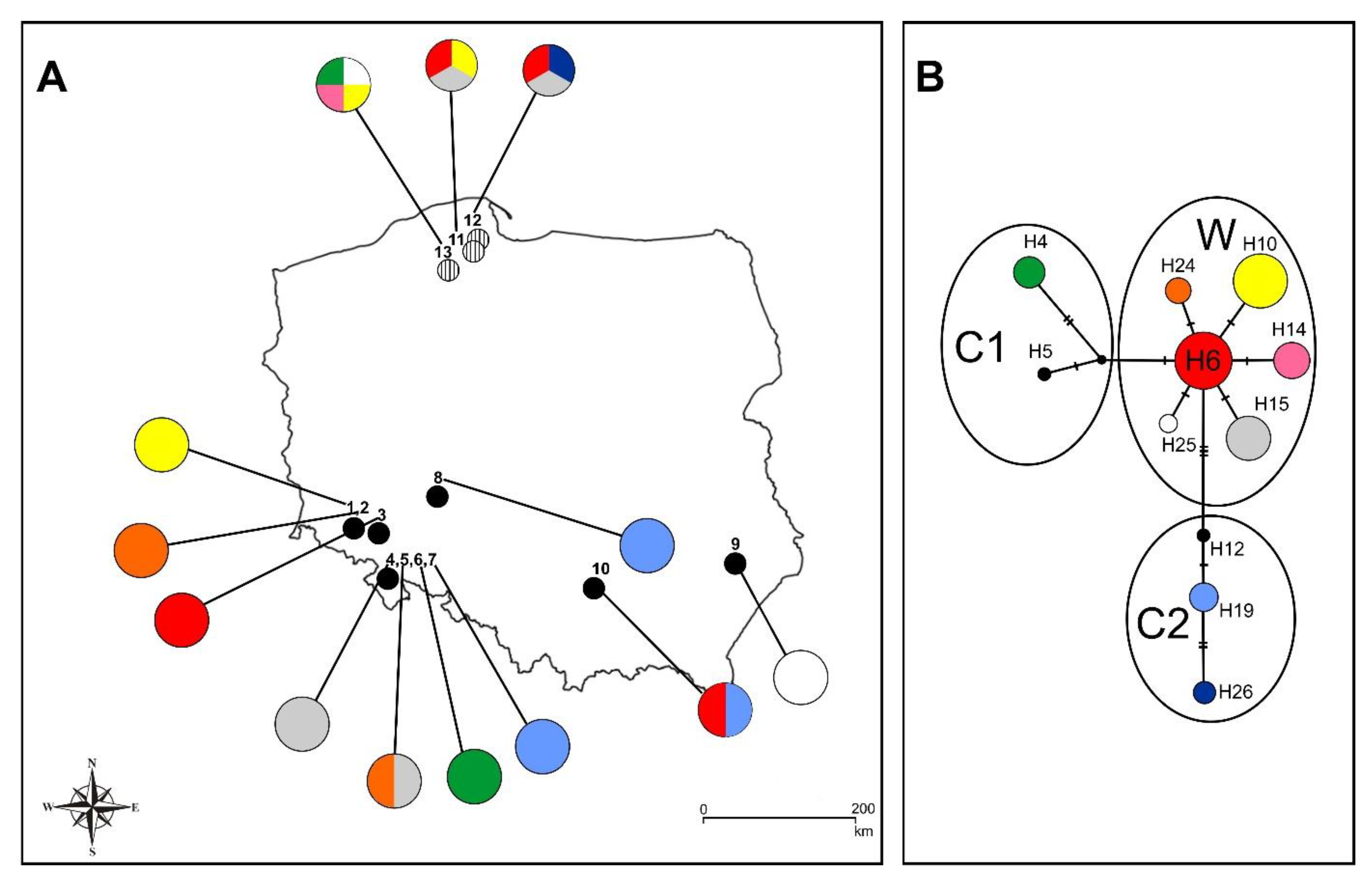
3.2. CpDNA Haplotype Variability
4. Discussion
4.1. History of Cypripedium calceolus
4.2. Plastid Variability of Polish Populations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kull, T. Fruit-set and recruitment in populations of Cypripedium calceolus L. in Estonia. Bot. J. Linn. Soc. 1998, 126, 27–38. [Google Scholar] [CrossRef]
- Kull, T. Biological flora of the British Isles-Cypripedium calceolus. J. Ecol. 1999, 87, 913–924. [Google Scholar] [CrossRef]
- Delforge, P. Orchids of Europe, North. Africa and the Middle East; A&C: London, UK, 2006. [Google Scholar]
- Szczęśniak, E.; Jakubska-Busse, A.; Śliwiński, M. Zróżnicowanie i rozmieszczenie zbiorowisk z udziałem Cypripedium calceolus L. (Orchidaceae) na Dolnym Śląsku. Acta Bot. Siles. 2012, 8, 97–128. [Google Scholar]
- Rankou, H.; Bilz, M. The IUCN Red List of Threatened Species. Available online: http://dx.doi.org/10.2305/IUCN.UK.2014-1.RLTS.T162021A43316125.en. (accessed on 15 June 2021).
- Minasiewicz, J.; Znaniecka, J.M.; Górniak, M.; Kawiński, A. Spatial genetic structure of an endangered orchid Cypripedium calceolus (Orchidaceae) at a regional scale: Limited gene flow in a fragmented landscape. Conserv. Genet. 2018, 19, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Terschuren, J. Action Plan for Cypripedium calceolus in Europe; Nature and environment, No 100; Council of Europe Publications: Strasbourg, France, 1999; p. 27. [Google Scholar]
- Jakubska-Busse, A.; Szczęśniak, E.; Śliwiński, M.; Narkiewicz, C. Zanikanie stanowisk obuwika pospolitego Cypripedium calceolus L., 1753 (Orchidaceae) w Sudetach. Przyr. Sudet. 2010, 13, 43–52. [Google Scholar]
- Jakubska-Busse, A.; Tsiftsis, S.; Śliwiński, M.; Křenová, Z.; Djordjević, V.; Steiu, C.; Kolanowska, M.; Efimov, P.; Hennigs, S.; Lustyk, P.; et al. How to protect natural habitats of rare terrestrial orchids effectively: A comparative case study of Cypripedium calceolus in different geographical regions of Europe. Plants 2021, 10, 404. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Dahlberg, C.J.; Carlgren, K.H.I.; Appelqvist, T. Pollination of the Lady’s slipper (Cypripedium calceolus) in Scandinavia-taxonomic and conservational aspects. Nord. J. Bot. 2009, 27, 266–273. [Google Scholar] [CrossRef]
- García, M.; Goñi, D.; Guzmán, D. Living at the edge: Local versus positional factors in the long-term population dynamics of an endangered orchid. Conserv. Biol. 2010, 24, 1219–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzosko, E.; Wróblewska, A.; Tałałaj, I.; Wasilewska, E. Genetic diversity of Cypripedium calceolus in Poland. Plant. Syst. Evol. 2011, 295, 83–96. [Google Scholar] [CrossRef]
- Kolanowska, M.; Jakubska-Busse, A. Is the lady’s-slipper orchid (Cypripedium calceolus) likely to shortly become extinct in Europe?-Insights based on ecological niche modelling. PLoS ONE 2020, 15, e0228420. [Google Scholar] [CrossRef]
- Kucharczyk, M. Krajowy Plan Ochrony Gatunku-Obuwik Pospolity (Cypripedium calceolus). 2018. Available online: https://doi.org/10.6084/m9.figshare.7223468.v3 (accessed on 29 April 2021).
- Brzosko, E.; Wróblewska, A.; Ratkiewicz, M. Spatial genetic structure and clonal diversity of island populations of lady’s slipper (Cypripedium calceolus) from the Biebrza National Park (northeast Poland). Mol. Ecol. 2002, 11, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Limpricht, W. Kalkpflanzen der westlichen Grafschaft Glantz. Fedders Rep. Beih. 1942, 131, 126–141. [Google Scholar]
- Limpricht, W. Kalkpflanzen der westlichen Grafschaft Glantz. Englers Bot. Jahrb. 1943, 73, 151–174. [Google Scholar]
- Limpricht, W. Kalkpflanzen Bober und Katchachgebirges. Bot. Jahrbücher Fur Syst. Pflanzengesch. Und Pflanzengeogr. 1944, 73, 375–417. [Google Scholar]
- Fay, M.F.; Bone, R.; Cook, P.; Kahandawala, I.; Greensmith, J.; Harris, S.; Pedersen, H.Æ.; Ingrouille, M.J.; Lexer, C. Genetic diversity in Cypripedium calceolus (Orchidaceae) with a focus on north-western Europe, as revealed by plastid DNA length polymorphisms. Ann. Bot. 2009, 104, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Gargiulo, R.; Pironon, S.; Zheleznaya, E.; Sanchez, M.D.; Balázs, Z.R.; Podar, D.; Wilkinson, T.; Jäkäläniemi, A.; Kull, T.; Väre, H.; et al. Phylogeography and post-glacial dynamics in the clonal-sexual orchid Cypripedium calceolus L. J. Biogeogr. 2019, 46, 526–538. [Google Scholar] [CrossRef]
- Taniguchi, H.; Nakamura, T.; Mizukami, H.; Kawano, S.; Sano, H.; Katsumi, M. Identity of Cypripedium calceolus (Orchidaceae) in Rebun Island: Comparative DNA analysis of related species. Genes Genet. Syst. 2001, 76, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, H.Æ.; Rasmussen, H.N.; Kahandawala, I.M.; Fay, M.F. Genetic diversity, compatibility patterns and seed quality in isolated populations of Cypripedium calceolus (Orchidaceae). Conserv. Genet. 2012, 13, 89–98. [Google Scholar] [CrossRef]
- Andronova, E.V.; Filippov, E.G. Мoрфoлoгические oсoбеннoсти цветкoв у самooпыляющихся растений Cypripedium calceolus и Cypripedium shanxiense (Orchidaceae). Бoтанический журнал 2007, 92, 360–364. (In Russian) [Google Scholar]
- Sundermann, H. Europäische und Mediterrane Orchideen: Eine Bestimmungsflora Mit Berücksichtigung der Ökologie; Brücke: Hildesheim, Germany, 1975. [Google Scholar]
- Cribb, P. The Genus Cypripedium; Timber Press: Portland, OR, USA, 1997; p. 301. [Google Scholar]
- Chase, M.W.; Hills, H.H. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Small, R.L.; Ryburn, J.A.; Cronn, R.C.; Seelanan, T.; Wendel, J.F. The Tor-Toise and the Hare: Choosing between noncoding plastome and nuclear Adh sequences for phylogeny reconstruction in a recently diverged plant group. Am. J. Bot. 1998, 85, 1301–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant. Mol Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Skinner, D.Z.; Liang, G.H.; Hulbert, S.H. Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor. Appl. Genet. 1994, 89, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Szlachetko, D.L.; Górniak, M.; Kowalkowska, A.K.; Kolanowska, M.; Jurczak-Kurek, A.; Archila-Morales, F. The natural history of the genus Cypripedium (Orchidaceae). Plant Biosyst. An Int. J. Deal. Asp. Plant Biol. 2021, 155, 772–796. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J. Tracer v1.5. 2009. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 17 February 2021).
- Guo, Y.Y.; Luo, Y.B.; Liu, Z.J.; Wang, X.Q. Evolution and Biogeography of the Slipper Orchids: Eocene Vicariance of the Conduplicate Genera in the Old and New World Tropics. PLoS ONE 2012, 7, e38788. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, S.R.; Gravendeel, B.; Singer, R.B.; Marshall, C.R.; Pierce, N.E. Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 2007, 448, 1042–1045. [Google Scholar] [CrossRef]
- Filippov, E.G.; Andronova, E.V. Genetic differentiation in plants of the genus Cypripedium from Russia inferred from allozyme data. Genetika 2011, 47, 615–623. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Szlachetko, D.L.; Kolanowska, M.; Muller, F.; Vannini, J.; Rojek, J.; Górniak, M. First Guatemalan record of natural hybridisation between Neotropical species of the lady’s slipper orchid (Orchidaceae, Cypripedioideae). PeerJ 2017, 5, e4162. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Mona, S.; Ray, N.; Arenas, M.; Excoffier, L. Genetic consequences of habitat fragmentation during a range expansion. Heredity 2014, 112, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Arenas, M.; Ray, N.; Currat, M.; Excoffier, L. Consequences of range contractions and range shifts on molecular diversity. Mol. Biol. Evol. 2012, 29, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Alignment Base Position | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 23–27 | 28–36 | 54–59 | 64–73 | 121–127 | 168–175 | 176–187 | 395–407 | 441–446 | 459–471 | 489 | 598–614 | 541–561 | 711–717 | 871–876 | |
| Haplotypes | |||||||||||||||
| H4 | - | + | + | - | + | + | - | - | + | - | T | - | - | - | + |
| H5 | - | + | + | - | + | + | - | + | + | - | T | - | + | + | + |
| H6 | - | + | + | - | + | + | - | - | + | - | T | - | + | + | - |
| H10 | - | + | + | - | + | - | - | - | + | - | T | - | + | + | - |
| H12 | - | + | + | + | - | + | - | - | + | - | C | - | + | + | - |
| H14 | + | + | + | - | + | + | - | - | + | - | T | - | + | + | - |
| H15 | - | + | + | - | + | + | + | - | + | - | T | - | + | + | - |
| H19 | - | + | + | + | - | + | - | - | + | + | C | - | + | + | - |
| H24 | - | + | - | - | + | + | - | - | + | - | T | - | + | + | - |
| H25 | - | - | + | - | + | + | - | - | + | - | T | - | + | + | - |
| H26 | - | + | + | + | - | + | - | - | - | + | C | + | + | + | - |
| Locality | Haplotype (accd-psa1) | GenBank Accession No | ITS Ribotype/GenBank Accession No | Haplotype (trnL–F)/GenBank Accession No | No. of Genets | Accd–psa1 Haplotype Localities (Country) * | |
|---|---|---|---|---|---|---|---|
| 1 | Mt. Młyniec | H10 | KF726125 | R3/MZ044292 | H1/MZ042956 | 4 | FR, ES, CH, UK |
| 2 | Mt. Połom | H24 | KF726126 | R2/MZ044290 | H1/MZ042957 | 3 | new haplotype |
| 3 | Grudno | H6 | KF726127 | R3/MZ044293 | H1/MZ042958 | 5 | PL, ES, SE, CH |
| 4 | Mt. Babilon (population extinct in 2016) | H15 | KF726129 | R1/MW785560 | H1/MZ042959 | 1 | PL |
| 5 | A nameless hill next to Mt. Słupiec (south slope) | H24 | KF726130 | R2/MW785559 | H1 MZ042960 | 4 | new haplotype |
| H24 | KF726130 | R3/MZ044294 | H1 MZ042960 | 2 | |||
| A nameless hill next to Mt. Słupiec (north slope) | H15 | KF726132 | R3/MZ044295 | H1/MZ042961 | 2 | PL | |
| 6 | Mt. Wapniarka | H4 | KF726134 | R3/MZ044296 | H1/MZ042962 | 5 | PL, SE |
| 7 | Mielnik | H19 | MW883889 | R1/MZ044291 | H1/MZ042963 | 6 | AT, IT |
| 8 | Nowa Ligota | H19 | KF726135 | R1/MZ044297 | H1/MZ042964 | 1 | AT, IT |
| H19 | KF726135 | R3/MW785561 | H1/MZ042964 | 4 | |||
| 9 | Kąty | H25 | KF726138 | R1/MZ044298 | H1/MZ042965 | 1 | new haplotype |
| 10 | Kalina Mała | H6 | MW883887 | R1/MZ044299 | H1/MZ042966 | 2 | PL, ES, SE, CH (H6); |
| H6 | MW883887 | R3/MZ044301 | H1/MZ042966 | 1 | |||
| H19 | MW883888 | R1/MZ044300 | H1/MZ042966 | 1 | AT, IT (H19) | ||
| 11 | Las Ostrzycki 1# | H10 | KP902524# | - | - | 8 | FR, ES, CH, UK |
| Las Ostrzycki 2_3# | H6 | KP902527# | - | - | 16 | PL, ES, SE, CH (H6) | |
| H15 | KP902525# | PL (H15) | |||||
| 12 | Prokowo# | H6 | KP902527# | - | - | 10 | PL, ES, SE, CH (H6); |
| H15 | KP902525# | PL (H15, H26) | |||||
| H26 | KP902528# | ||||||
| 13 | Jezioro Głuche/Bukówki# | H4 | KP902531# | - | - | 22 | PL, SE (H4) |
| H10 | KP902530# | FR, UK, ES, CH (H10) | |||||
| H14 | KP902529# | SE, FR, UK (H14) | |||||
| H25 | KP902524# | PL (H25) | |||||
| Poland West, North East, Estonia * | H4 | Personal information | - | - | 7 | PL, SE (H4) | |
| H5 | PL (H5) | ||||||
| H6 | PL, SE, ES, CH (H6) | ||||||
| H12 | PL (H12) | ||||||
| H15 | PL (H15) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Górniak, M.; Jakubska-Busse, A.; Ziętara, M.S. Genetic History of the Remnant Population of the Rare Orchid Cypripedium calceolus Based on Plastid and Nuclear rDNA. Genes 2021, 12, 940. https://doi.org/10.3390/genes12060940
Górniak M, Jakubska-Busse A, Ziętara MS. Genetic History of the Remnant Population of the Rare Orchid Cypripedium calceolus Based on Plastid and Nuclear rDNA. Genes. 2021; 12(6):940. https://doi.org/10.3390/genes12060940
Chicago/Turabian StyleGórniak, Marcin, Anna Jakubska-Busse, and Marek S. Ziętara. 2021. "Genetic History of the Remnant Population of the Rare Orchid Cypripedium calceolus Based on Plastid and Nuclear rDNA" Genes 12, no. 6: 940. https://doi.org/10.3390/genes12060940
APA StyleGórniak, M., Jakubska-Busse, A., & Ziętara, M. S. (2021). Genetic History of the Remnant Population of the Rare Orchid Cypripedium calceolus Based on Plastid and Nuclear rDNA. Genes, 12(6), 940. https://doi.org/10.3390/genes12060940