
Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration


Abstract
:1. Introduction
- closure and wound healing;
- immune and/or nonspecific defense reaction;
- recruitment of cells that are to form a regenerate;
- growth of the regenerate;
- patterning and differentiation.
2. The Transcriptomic Era
3. Back to the Future
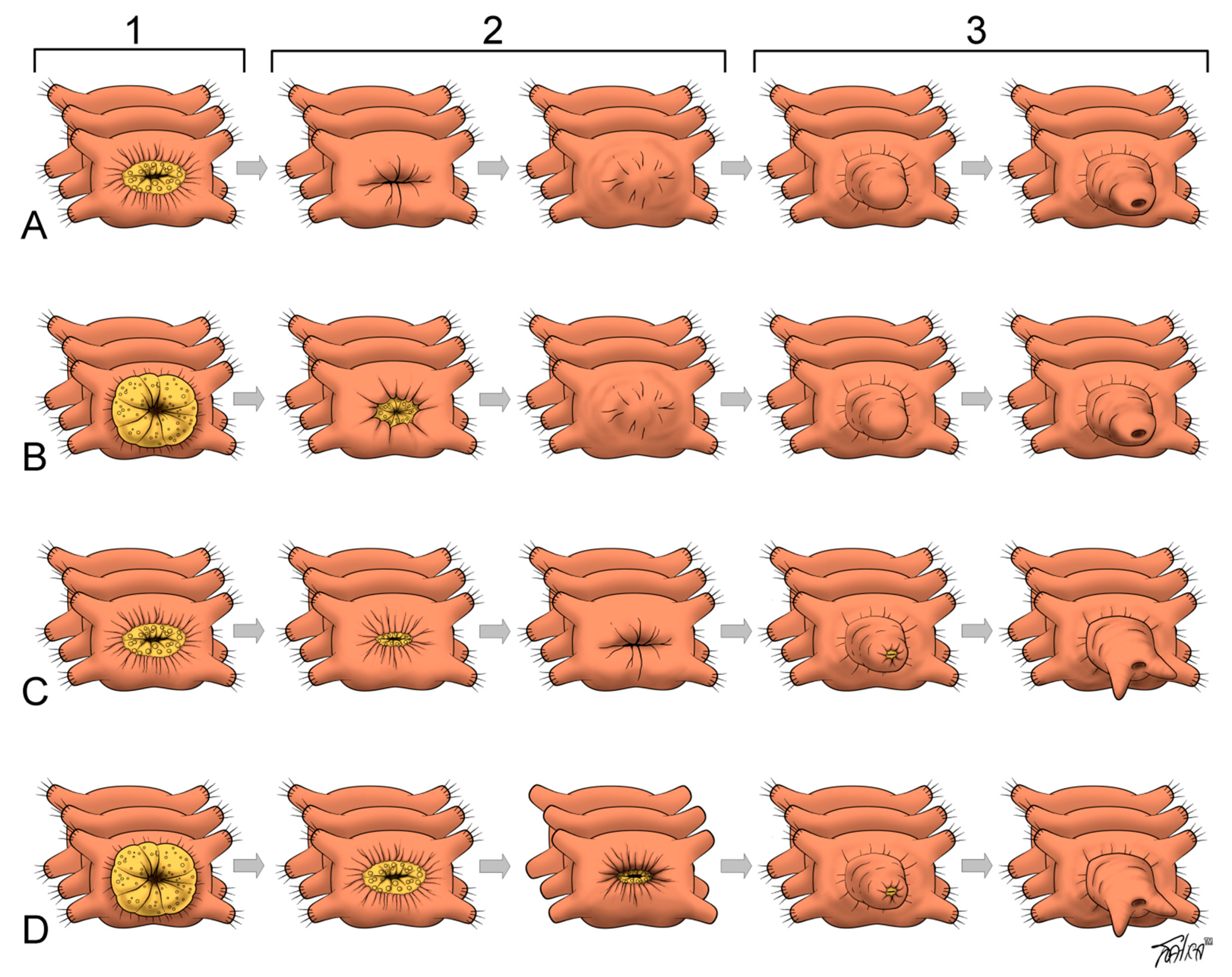
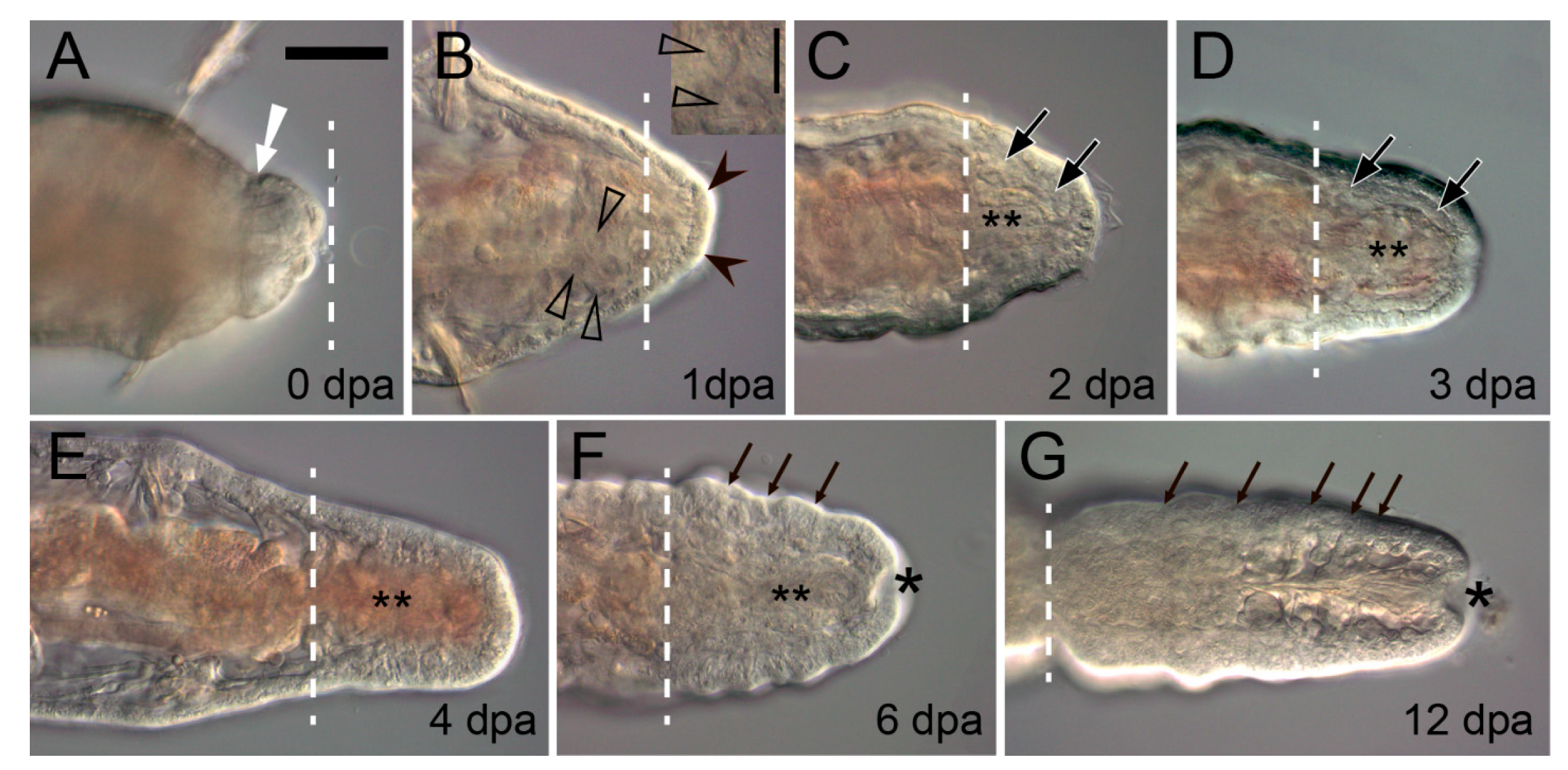
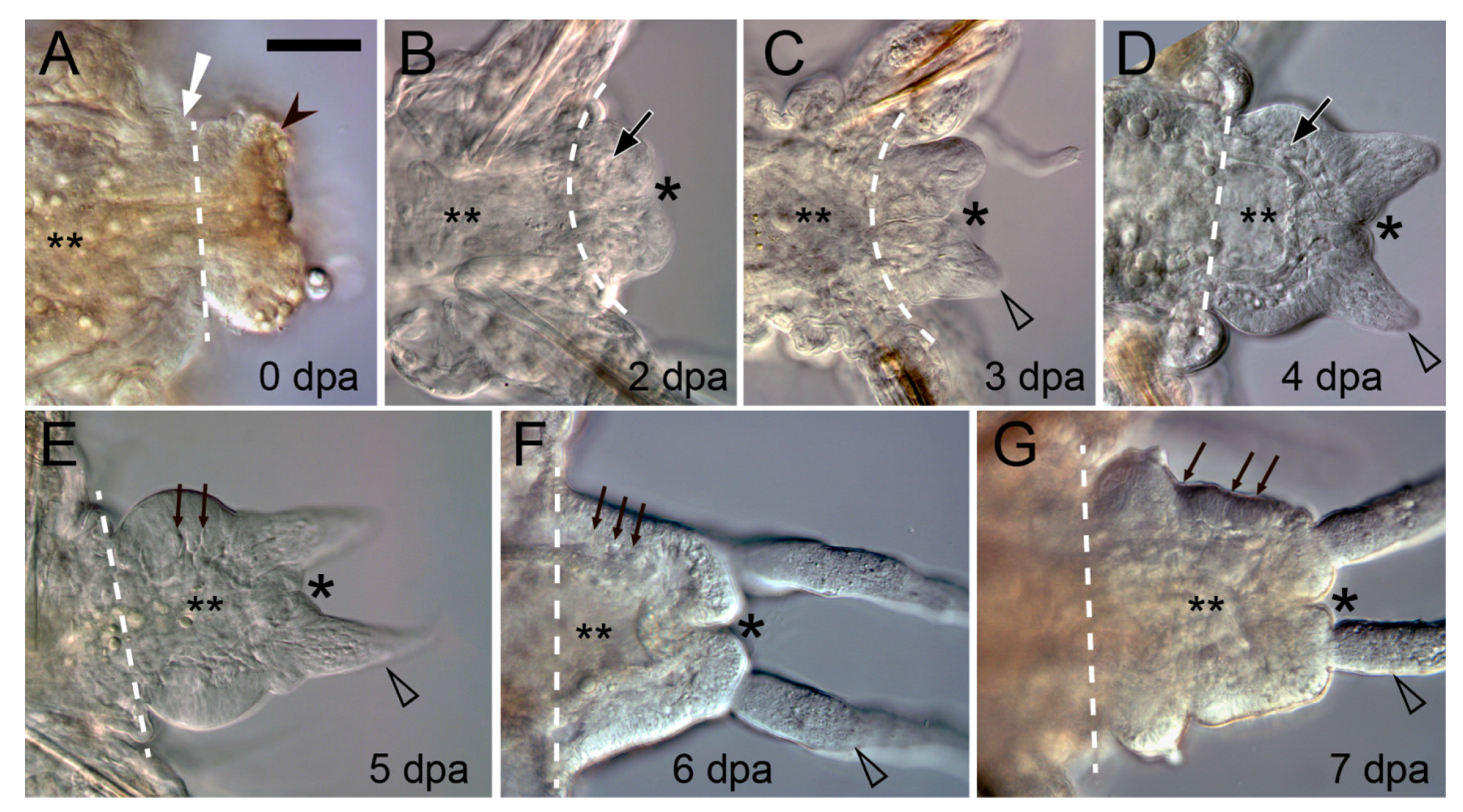
3.1. Early Events during Regeneration: Wound Healing and Immune Response
3.2. Cellular Sources of the Blastema
3.3. Regeneration of Specific Tissues
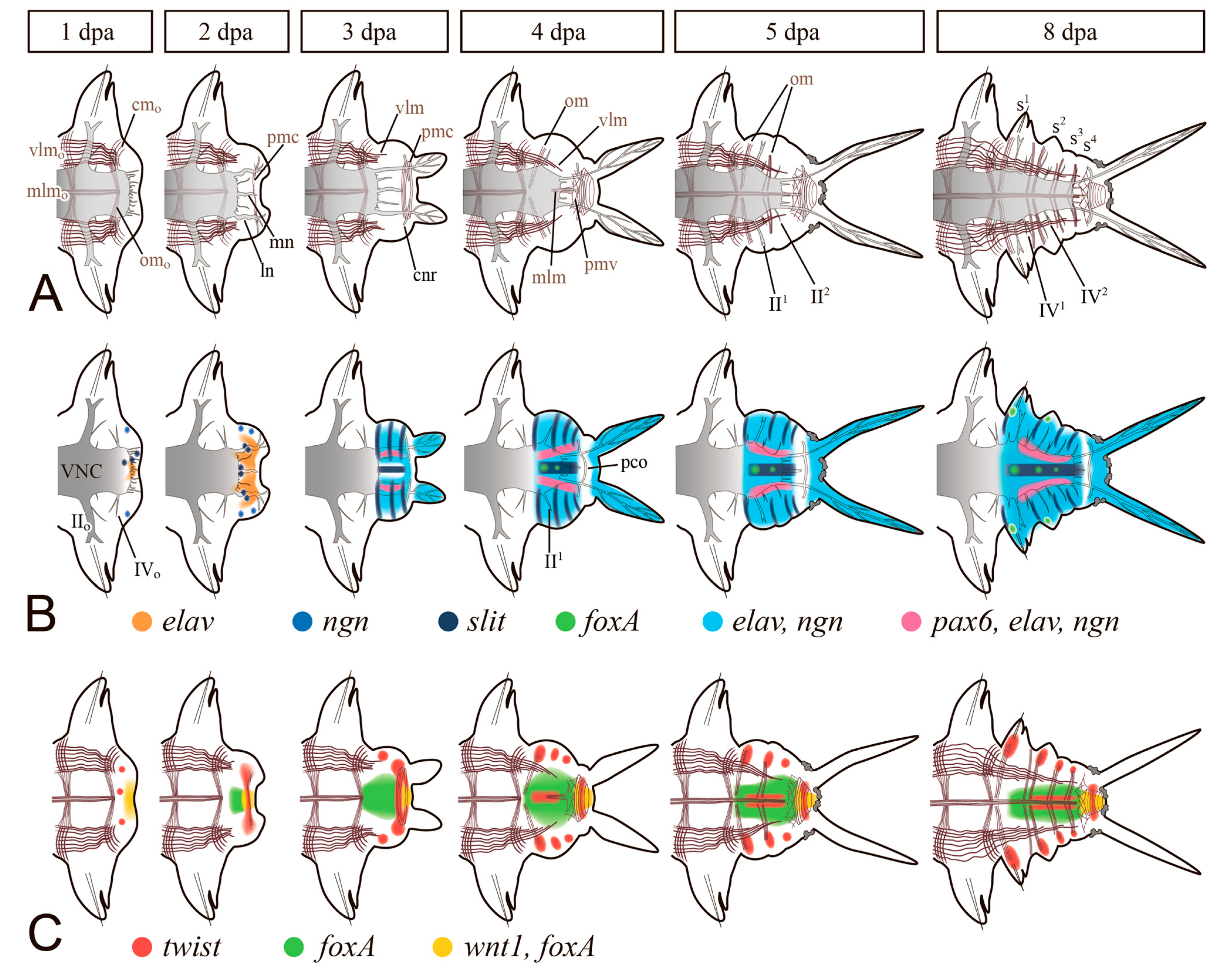
3.3.1. Nervous System
3.3.2. Coelom Wall and Muscular System
3.3.3. Digestive System
3.3.4. Gonad and Germ Cell De Novo Formation during Regeneration and Asexual Reproduction in Annelids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgan, T.H. Regeneration; Columbia Univ. Biol. Ser.; Macmillan: Norwood, MA, USA, 1901; 376р. [Google Scholar]
- Vorontsova, M.A.; Liosner, L.D. Asexual Propagation and Regeneration; Pergamon Press: London, UK, 1960; 489p. [Google Scholar]
- Carlson, B.M. Principles of Regenerative Biology; Elsevier/Academic Press: Amsterdam, The Netherlands; Burlington, MA, USA, 2007; ISBN 978-0-12-369439-3. [Google Scholar]
- Kostyuchenko, R.P.; Kozin, V.V.; Kupriashova, E.E. Regeneration and asexual reproduction in annelids: Cells, genes, and evolution. Biol. Bull. 2016, 43, 185–194. [Google Scholar] [CrossRef]
- Stocum, D.L. Regenerative Biology and Medicine, 2nd ed.; Elsevier Science Publ. Co. Inc.: Amsterdam, The Netherlands, 2012; 474p. [Google Scholar]
- Korotkova, G.P. Regeneration in Animals; Saint-Petersburg University Press: Saint-Petersburg, Russia, 1997; ISBN 5-288-01322-5. [Google Scholar]
- Bely, A.E.; Nyberg, G.K. Evolution of animal regeneration: Re-emergence of a field. Trends Ecol. Evol. 2010, 25, 161–170. [Google Scholar] [CrossRef]
- Lai, A.G.; Aboobaker, A.A. EvoRegen in animals: Time to uncover deep conservation or convergence of adult stem cell evolution and regenerative processes. Dev. Biol. 2018, 433, 118–131. [Google Scholar] [CrossRef]
- Bely, A.E. Distribution of segment regeneration ability in the Annelida. Integr. Comp. Biol. 2006, 46, 508–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bely, A.E. Early Events in Annelid Regeneration: A Cellular Perspective. Integr. Comp. Biol. 2014, 54, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Nikanorova, D.D.; Kupriashova, E.E.; Kostyuchenko, R.P. Regeneration in Annelids: Cell Sources, Tissue Remodeling, and Differential Gene Expression. Russ. J. Dev. Biol. 2020, 51, 148–161. [Google Scholar] [CrossRef]
- Zattara, E. Axial Regeneration in Segmented Animals A Post-Embryonic Reboot of the Segmentation Process. In Cellular Processes in Segmentation; CRC Press Taylor & Francis Group: Abingdon Oxfordshire, UK, 2020; pp. 255–292. ISBN 978-0-429-42360-4. [Google Scholar]
- Bely, A.E. Decoupling of fission and regenerative capabilities in an asexual oligochaete. Hydrobiologia 1999, 406, 243–251. [Google Scholar] [CrossRef]
- Kharin, A.V.; Zagainova, I.V.; Kostyuchenko, R.P. Formation of the paratomic fission zone in freshwater oligochaetes. Russ. J. Dev. Biol. 2006, 37, 354–365. [Google Scholar] [CrossRef]
- Zattara, E.E.; Bely, A.E. Phylogenetic distribution of regeneration and asexual reproduction in Annelida: Regeneration is ancestral and fission evolves in regenerative clades. Invert. Biol. 2016, 135, 400–414. [Google Scholar] [CrossRef] [Green Version]
- Bely, A.E.; Sikes, J.M. Latent regeneration abilities persist following recent evolutionary loss in asexual annelids. Proc. Natl. Acad. Sci. USA 2010, 107, 1464–1469. [Google Scholar] [CrossRef] [Green Version]
- Iwanoff, P.P. Die Entwiklung der Larvalsegmente bei den Annelide. Z. Morph. Oekol. Tiere. 1928, 10, 62–161. [Google Scholar] [CrossRef]
- Nengwen, X.; Feng, G.; Edwards, C.A. The regeneration capacity of an earthworm, Eisenia fetida, in relation to the site of amputation along the body. Acta Ecol. Sinica. 2011, 31, 197–204. [Google Scholar] [CrossRef]
- Kostyuchenko, R.P.; Kozin, V.V. Morphallaxis versus Epimorphosis? Cellular and Molecular Aspects of Regeneration and Asexual Reproduction in Annelids. Biol. Bull. 2020, 47, 237–246. [Google Scholar] [CrossRef]
- Hyman, L.H. Aspects of Regeneration in Annelids. Am. Nat. 1940, 74, 513–527. [Google Scholar] [CrossRef]
- Herlant-Meewis, H. Regeneration in Annelids. Adv. Morphog. 1964, 4, 155–215. [Google Scholar]
- Ribeiro, R.P.; Ponz-Segrelles, G.; Bleidorn, C.; Aguado, M.T. Comparative Transcriptomics in Syllidae (Annelida) Indicates That Posterior Regeneration and Regular Growth Are Comparable, While Anterior Regeneration Is a Distinct Process. BMC Genom. 2019, 20, 855. [Google Scholar] [CrossRef]
- Myohara, M.; Niva, C.C.; Lee, J.M. Molecular Approach to Annelid Regeneration: CDNA Subtraction Cloning Reveals Various Novel Genes That Are Upregulated during the Large-Scale Regeneration of the Oligochaete, Enchytraeus Japonensis. Dev. Dyn. 2006, 235, 2051–2070. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-J.; Lee, M.S.; Tak, E.S.; Lee, E.; Koh, K.S.; Ahn, C.H.; Park, S.C. Gene expression profile in the anterior regeneration of the earthworm using expressed sequence tags. Biosci. Biotechnol. Biochem. 2009, 73, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyberg, K.G.; Conte, M.A.; Kostyun, J.L.; Forde, A.; Bely, A.E. Transcriptome characterization via 454 pyrosequencing of the annelid Pristina leidyi, an emerging model for studying the evolution of regeneration. BMC Genom. 2012, 13, 287. [Google Scholar] [CrossRef] [Green Version]
- Bhambri, A.; Dhaunta, N.; Patel, S.S.; Hardikar, M.; Bhatt, A.; Srikakulam, N.; Shridhar, S.; Vellarikkal, S.; Pandey, R.; Jayarajan, R.; et al. Large Scale Changes in the Transcriptome of Eisenia Fetida during Regeneration. PLoS ONE 2018, 13, e0204234. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Zunjarrao, S.; Pillai, B. Neev, a novel long non-coding RNA, is expressed in chaetoblasts during regeneration of Eisenia fetida. J. Exp. Biol. 2020, 223, jeb216754. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wang, X.-B.; Zhang, J.-J.; Li, M.-L.; Wu, S.-S.; Ma, X.-Y.; Wang, X.; Zhao, H.-F.; Li, Y.; Zhu, H.H.; et al. Genome and Single-Cell RNA-Sequencing of the Earthworm Eisenia Andrei Identifies Cellular Mechanisms Underlying Regeneration. Nat. Commun. 2020, 11, 2656. [Google Scholar] [CrossRef]
- Paul, S.; Balakrishnan, S.; Arumugaperumal, A.; Lathakumari, S.; Syamala, S.S.; Arumugaswami, V.; Sivasubramaniam, S. The Transcriptome of Anterior Regeneration in Earthworm Eudrilus Eugeniae. Mol. Biol. Rep. 2021, 48, 259–283. [Google Scholar] [CrossRef] [PubMed]
- Tellez-Garcia, A.A.; Álvarez-Martínez, R.; MaríaLópez-Martínez, J.; Arellano-Carbajal, F. Transcriptome analysis during early regeneration of Lumbriculus variegatus. Gene Rep. 2021, 23, 101050. [Google Scholar] [CrossRef]
- Cornec, J.-P.; Cresp, J.; Delye, P.; Hoarau, F.; Reynaud, G. Tissue Responses and Organogenesis during Regeneration in the Oligochete Limnodrilus hoffmeisteri (Clap.). Can. J. Zool. 1987, 65, 403–414. [Google Scholar] [CrossRef]
- Huguet, G.; Molinas, M. Myofibroblast-like Cells and Wound Contraction in Leech Wound Healing. J. Exp. Zool. 1996, 275, 308–316. [Google Scholar] [CrossRef]
- Bodó, K.; Kellermayer, Z.; László, Z.; Boros, Á.; Kokhanyuk, B.; Németh, P.; Engelmann, P. Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia Andrei. Int. J. Mol. Sci. 2021, 22, 2363. [Google Scholar] [CrossRef]
- Kostyuchenko, R.P.; Kozin, V.V.; Filippova, N.A.; Sorokina, E.V. FoxA expression pattern in two polychaete species, Alitta virens and Platynereis dumerilii: Examination of the conserved key regulator of the gut development from cleavage through larval life, postlarval growth, and regeneration. Dev. Dyn. 2019, 248, 728–743. [Google Scholar] [CrossRef]
- Brockes, J.P.; Kumar, A. Comparative aspects of animal regeneration. Annu. Rev. Cell Dev. Biol. 2008, 24, 25–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupin, F.; Coulon, J.; Parco, Y.L.; Fontes, M.; Thouveny, Y. Formation of the extracellular matrix during the epimorphic anterior regeneration of Owenia fusiformis: Autoradiographical and in situ hybridization studies. Int. J. Dev. Biol. 1991, 35, 109–119. [Google Scholar]
- Fontés, M.; Coulon, J.; Delgross, M.-H.; Thouveny, Y. Muscle Dedifferentiation and Contractile Protein Synthesis during Post-Traumatic Regeneration by Owenia Fusiformis (Polychaete Annelid). Cell Differ. 1983, 13, 267–282. [Google Scholar] [CrossRef]
- Zattara, E.E.; Bely, A.E. Evolution of a Novel Developmental Trajectory: Fission Is Distinct from Regeneration in the Annelid Pristina Leidyi. Evol. Dev. 2011, 13, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Planques, A.; Malem, J.; Parapar, J.; Vervoort, M.; Gazave, E. Morphological, Cellular and Molecular Characterization of Posterior Regeneration in the Marine Annelid Platynereis Dumerilii. Dev. Biol. 2019, 445, 189–210. [Google Scholar] [CrossRef]
- de Jong, D.M.; Seaver, E.C. Investigation into the Cellular Origins of Posterior Regeneration in the Annelid Capitella Teleta. Regeneration 2018, 5, 61–77. [Google Scholar] [CrossRef]
- Kozin, V.V.; Kostyuchenko, R.P. Vasa, PL10, and Piwi Gene Expression during Caudal Regeneration of the Polychaete Annelid Alitta Virens. Dev. Genes Evol. 2015, 225, 129–138. [Google Scholar] [CrossRef]
- Sugio, M.; Yoshida-noro, C.; Ozawa, K. Tochinai S. Stem cells in asexual reproduction of Enchytraeus japonensis (Oligochaeta, Annelid): Proliferation and migration of neoblasts. Dev. Growth Differ. 2012, 54, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Paulus, T.; Müller, M.C.M. Cell proliferation dynamics and morphological differentiation during regeneration in Dorvillea bermudensis (Polychaeta, Dorvilleidae). J. Morphol. 2006, 267, 393–403. [Google Scholar] [CrossRef]
- Bilello, A.A.; Potswald, H.E. A cytological and quantitative study of neoblasts in the naid Ophidonais serpentina (Oligochaeta). Wilhelm Roux. Arch. Entwickl. Mech. Org. 1974, 174, 234–249. [Google Scholar] [CrossRef]
- Ribeiro, R.P.; Egger, B.; Ponz-Segrelles, G.; Aguado, M.T. Cellular proliferation dynamics during regeneration in Syllis malaquini (Syllidae, Annelida). Front. Zool. 2021, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Shalaeva, A.Y.; Kostyuchenko, R.P.; Kozin, V.V. Structural and Functional Characterization of the FGF Signaling Pathway in Regeneration of the Polychaete Worm Alitta Virens (Annelida, Errantia). Genes 2021, 12, 788. [Google Scholar] [CrossRef] [PubMed]
- Juliano, C.E.; Swartz, S.Z.; Wessel, G.M. A conserved germline multipotency program. Development 2010, 137, 4113–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özpolat, B.D.; Bely, A.E. Gonad Establishment during Asexual Reproduction in the Annelid Pristina Leidyi. Dev. Biol. 2015, 405, 123–136. [Google Scholar] [CrossRef]
- Randolph, H. The Regeneration of the Tail in Lumbriculus. J. Morphol. 1892, 7, 317–344. [Google Scholar] [CrossRef] [Green Version]
- Newmark, P.A.; Alvarado, A.S. Regeneration in Planaria. eLS 2001, 1–7. [Google Scholar]
- Zattara, E.E.; Turlington, K.W.; Bely, A.E. Long-Term Time-Lapse Live Imaging Reveals Extensive Cell Migration during Annelid Regeneration. BMC Dev. Biol. 2016, 16, 6. [Google Scholar] [CrossRef] [Green Version]
- Sugio, M.; Takeuchi, K.; Kutsuna, J.; Tadokoro, R.; Takahashi, Y.; Yoshida-noro, C.; Tochinai, S. Exploration of embryonic origins of germline stem cells and neoblasts in Enchytraeus japonensis (Oligochaeta, Annelida). Gene Expr. Patterns. 2008, 8, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, R.; Sugio, M.; Kutsuna, J.; Tochinai, S.; Takahashi, Y. Early Segregation of Germ and Somatic Lineages during Gonadal Regeneration in the Annelid Enchytraeus Japonensis. Curr. Biol. 2006, 16, 1012–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida-Noro, C.; Tochinai, S. Stem cell system in asexual and sexual reproduction of Enchytraeus japonensis (Oligochaeta, Annelida). Dev. Growth Differ. 2010, 52, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Myohara, M. What Role Do Annelid Neoblasts Play? A Comparison of the Regeneration Patterns in a Neoblast-Bearing and a Neoblast-Lacking Enchytraeid Oligochaete. PLoS ONE 2012, 7, e37319. [Google Scholar] [CrossRef]
- Özpolat, B.D.; Bely, A.E. Developmental and Molecular Biology of Annelid Regeneration: A Comparative Review of Recent Studies. Curr. Opin. Genet. Dev. 2016, 40, 144–153. [Google Scholar] [CrossRef]
- Tweeten, K.A.; Anderson, A. Analysis of cell proliferation and migration during regeneration in Lumbriculus variegatus (Clitellata: Lumbriculidae). Bione 2008, 79, 183–190. [Google Scholar] [CrossRef]
- Gazave, E.; Behague, J.; Laplane, L.; Guillou, A.; Preau, L.; Demilly, A.; Balavoine, G.; Vervoort, M. Posterior elongation in the annelid Platynereis dumerilii involves stem cells molecularly related to primordial germ cells. Dev. Biol. 2013, 382, 246–267. [Google Scholar] [CrossRef] [PubMed]
- Giani, V.C.; Yamaguchi, E.; Boyle, M.J.; Seaver, E.C. Somatic and germline expression of piwi during development and regeneration in the marine polychaete annelid Capitella teleta. EvoDevo 2011, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Oyama, A.; Shimizu, T. Transient occurrence of vasa-expressing cells in nongenital segments during embryonic development in the oligochaete annelid Tubifex tubifex. Dev. Genes Evol. 2007, 217, 675–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyama, A.; Yoshida, H.; Shimizu, T. Embryonic expression of p68, a DEAD-box RNA helicase, in the oligochaete annelid Tubifex tubifex. Gene Expr. Patterns. 2008, 8, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Berrill, N.J. Regeneraton and Budding in Worms. Biol. Rev. 1952, 27, 401–438. [Google Scholar] [CrossRef]
- Clark, M.E. Later Stages of Regeneration in the Polychaete, Nephtys. J. Morphol. 1968, 124, 483–510. [Google Scholar] [CrossRef]
- Müller, M.C.M. Nerve Development, Growth and Differentiation during Regeneration in Enchytraeus Fragmentosus and Stylaria Lacustris (Oligochaeta). Dev. Growth Differ. 2004, 46, 471–478. [Google Scholar] [CrossRef]
- Goss, R.J. Principles of Regeneration; Elsevier: Amsterdam, The Netherlands, 1969; ISBN 978-1-4832-3250-8. [Google Scholar]
- Hulsebosch, C.E.; Bittner, G.D. Regeneration of Axons and Nerve Cell Bodies in the CNS of Annelids. J. Comp. Neurol. 1981, 198, 77–88. [Google Scholar] [CrossRef]
- Moffett, S.B. Nervous System Regeneration in the Invertebrates; Zoophysiology; Springer: Berlin/Heidelberg, Germany, 1996; Volume 34, ISBN 978-3-642-79841-2. [Google Scholar]
- Birse, S.C.; Bittner, G.D. Regeneration of Earthworm Giant Axons Following Transection or Ablation. J. Neurophysiol. 1981, 45, 724–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewes, C.D.; Vining, E.P.; Zoran, M.J. Regeneration of Rapid Escape Reflex Pathways in Earthworms. Am. Zool. 1988, 28, 1077–1089. [Google Scholar] [CrossRef]
- Lyckman, A.W.; Heidelbaugh, S.M.; Bittner, G.D. Analysis of Neuritic Outgrowth from Severed Giant Axons in Lumbricus Terrestris. J. Comp. Neurol. 1992, 318, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Lesiuk, N.M.; Drewes, C.D. Behavioral Plasticity and Central Regeneration of Locomotor Reflexes in the Freshwater Oligochaete, Lumbriculus Variegatus. II: Ablation Studies. Invertebr. Biol. 2001, 120, 259–268. [Google Scholar] [CrossRef]
- Baylor, D.A.; Nicholls, J.G. Patterns of Regeneration between Individual Nerve Cells in the Central Nervous System of the Leech. Nature 1971, 232, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Muller, K.J.; McGlade-McCulloh, E.; Mason, A. Tinkering with Successful Synapse Regeneration in the Leech: Adding Insult to Injury. J. Exp. Biol. 1987, 132, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, B.; Pellegrino, M. Remodelling of an Intact Neurone in the Central Nervous System of the Leech. J. Exp. Biol. 1995, 198, 1989–1994. [Google Scholar] [CrossRef]
- Blackshaw, S.E.; Arkison, S.; Cameron, C.; Davies, J.A. Promotion of Regeneration and Axon Growth Following Injury in an Invertebrate Nervous System by the Use of Three–Dimensional Collagen Gels. Proc. R. Soc. Lond. B Biol. Sci. 1997, 264, 657–661. [Google Scholar] [CrossRef] [Green Version]
- Burrell, B.D.; Sahley, C.L.; Muller, K.J. Progressive Recovery of Learning during Regeneration of a Single Synapse in the Medicinal Leech. J. Comp. Neurol. 2003, 457, 67–74. [Google Scholar] [CrossRef]
- Muller, K.J.; Scott, S.A. Correct Axonal Regeneration after Target Cell Removal in the Central Nervous System of the Leech. Science 1979, 206, 87–89. [Google Scholar] [CrossRef]
- de Jong, D.M.; Seaver, E.C. A Stable Thoracic Hox Code and Epimorphosis Characterize Posterior Regeneration in Capitella Teleta. PLoS ONE 2016, 11, e0149724. [Google Scholar] [CrossRef] [Green Version]
- Kozin, V.V.; Filippova, N.A.; Kostyuchenko, R.P. Regeneration of the Nervous and Muscular System after Caudal Amputation in the Polychaete Alitta Virens (Annelida: Nereididae). Russ. J. Dev. Biol. 2017, 48, 198–210. [Google Scholar] [CrossRef]
- Avel, M. L’influence Du Système Nerveux Sur La Régénération Chez Les Urodèles et Les Oligochètes. Bull. Soc. Zool. Fr. 1961, 86, 464–483. [Google Scholar]
- Bouc-Lassalle, A.M. The cicatrization of wounds of the body wall of the Lombrician Eisenia foetida Sav., in the presence and absence of innervation. Comptes Rendus Hebd. Seances Acad. Sci. Ser. Sci. Nat. 1968, 267, 2167–2169. [Google Scholar]
- Müller, M.C.M.; Berenzen, A.; Westheide, W. Experiments on Anterior Regeneration in Eurythoe Complanata (“Polychaeta”, Amphinomidae): Reconfiguration of the Nervous System and Its Function for Regeneration. Zoomorphology 2003, 122, 95–103. [Google Scholar] [CrossRef]
- Weidhase, M.; Beckers, P.; Bleidorn, C.; Aguado, M.T. Nervous System Regeneration in Typosyllis Antoni (Annelida: Syllidae). Zool. Anz. J. Comp. Zool. 2017, 269, 57–67. [Google Scholar] [CrossRef]
- Starunov, V.V.; Barmasova, G.A.; Nesterenko, M.A.; Kulakova, M.A.; Novikova, E.L. Pygospio Elegans (Annelida: Spionidae)—An Annelid Model for Regeneration Studies. Invertebr. Zool. 2020, 17, 247–266. [Google Scholar] [CrossRef]
- Yoshida-Noro, C.; Myohara, M.; Kobari, F.; Tochinai, S. Nervous System Dynamics during Fragmentation and Regeneration in Enchytraeus Japonensis (Oligochaeta, Annelida). Dev. Genes Evol. 2000, 210, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.C.M.; Henning, L. Ground Plan of the Polychaete Brain—I. Patterns of Nerve Development during Regeneration in Dorvillea Bermudensis (Dorvilleidae). J. Comp. Neurol. 2004, 471, 49–58. [Google Scholar] [CrossRef]
- Myohara, M. Differential Tissue Development during Embryogenesis and Regeneration in an Annelid. Dev. Dyn. 2004, 231, 349–358. [Google Scholar] [CrossRef]
- Weidhase, M.; Bleidorn, C.; Helm, C. Structure and Anterior Regeneration of Musculature and Nervous System in Cirratulus Cf. Cirratus (Cirratulidae, Annelida). J. Morphol. 2014, 275, 1418–1430. [Google Scholar] [CrossRef]
- Weidhase, M.; Helm, C.; Bleidorn, C. Morphological Investigations of Posttraumatic Regeneration in Timarete Cf. Punctata (Annelida: Cirratulidae). Zool. Lett. 2015, 1, 20. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.S.; Kim, J.; Yi, J.; Park, S.C.; Lee, H.-Y.; Cho, S.-J. Characterization of Perionyx Excavatus Development and Its Head Regeneration. Biology 2020, 9, 273. [Google Scholar] [CrossRef]
- Pfeifer, K.; Dorresteijn, A.W.C.; Fröbius, A.C. Activation of Hox Genes during Caudal Regeneration of the Polychaete Annelid Platynereis Dumerilii. Dev. Genes Evol. 2012, 222, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Drewes, C.D.; Fourtner, C.R. Morphallaxis in an Aquatic Oligochaete, Lumbriculus Variegatus: Reorganization of Escape Reflexes in Regenerating Body Fragments. Dev. Biol. 1990, 138, 94–103. [Google Scholar] [CrossRef]
- Martinez, V.G.; Reddy, P.K.; Zoran, M.J. Asexual Reproduction and Segmental Regeneration, but Not Morphallaxis, Are Inhibited by Boric Acid in Lumbriculus Variegatus (Annelida: Clitellata: Lumbriculidae). Hydrobiologia 2006, 564, 73–86. [Google Scholar] [CrossRef]
- Martinez-Acosta, V.G.; Zoran, M.J. Evolutionary Aspects of Annelid Regeneration. eLS 2015. [Google Scholar] [CrossRef]
- Lybrand, Z.R.; Zoran, M.J. Rapid Neural Circuit Switching Mediated by Synaptic Plasticity during Neural Morphallactic Regeneration. Dev. Neurobiol. 2012, 72, 1256–1266. [Google Scholar] [CrossRef]
- Zoran, M.J.; Martinez, V.G. Lumbriculus variegatus and the Need for Speed: A Model System for Rapid Escape, Regeneration and Asexual Reproduction. In Annelids in Modern Biology; Shain, D.H., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; pp. 185–202. ISBN 978-0-470-45520-3. [Google Scholar]
- Martinez, V.G.; Manson, J.M.B.; Zoran, M.J. Effects of Nerve Injury and Segmental Regeneration on the Cellular Correlates of Neural Morphallaxis. J. Exp. Zoolog. B Mol. Dev. Evol. 2008, 310B, 520–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackshaw, S.E.; Babington, E.J.; Emes, R.D.; Malek, J.; Wang, W.-Z. Identifying Genes for Neuron Survival and Axon Outgrowth in Hirudo Medicinalis. J. Anat. 2004, 204, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.G.; Menger, G.J.; Zoran, M.J. Regeneration and Asexual Reproduction Share Common Molecular Changes: Upregulation of a Neural Glycoepitope during Morphallaxis in Lumbriculus. Mech. Dev. 2005, 122, 721–732. [Google Scholar] [CrossRef]
- Niva, C.C.; Lee, J.M.; Myohara, M. Glutamine Synthetase Gene Expression during the Regeneration of the Annelid Enchytraeus Japonensis. Dev. Genes Evol. 2008, 218, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Novikova, E.L.; Bakalenko, N.I.; Nesterenko, A.Y.; Kulakova, M.A. Expression of Hox Genes during Regeneration of Nereid Polychaete Alitta (Nereis) Virens (Annelida, Lophotrochozoa). EvoDevo 2013, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, T.H. Experimental Studies of the Internal Factors of Regeneration in the Earthworm. Arch. Für Entwickl. Org. 1902, 14, 562–591. [Google Scholar] [CrossRef]
- von Haffner, K. Die überzähligen Bildungen des Körperstammes von Lumbriculus variegatus Müll. und ihre kausale Analyse. Wilhelm Roux Arch. Für Entwickl. Org. 1931, 123, 649–681. [Google Scholar] [CrossRef] [PubMed]
- von Haffner, K. Über Die Regeneration der Vordersten Segmente von Lumbriculus Und Ihre Fahigkeit, Ein Hinterende Zu Regenerieren. Z. Wk. Zool. 1928, 132, 37–72. [Google Scholar]
- Janda, V. Über die Lebensdauer und reparativen Potenzen bauchstrangloser Fragmente und künstlich vereinigter dorsaler Körperhälften von Criodrilus lacuum Hoffm. Wilhelm Roux Arch. Für Entwickl. Org. 1930, 122, 432–450. [Google Scholar] [CrossRef]
- Holmes, G.E. The Influence of the Nervous System on Regeneration in Nereis Virens, Sars. J. Exp. Zool. 1931, 60, 485–509. [Google Scholar] [CrossRef]
- Zhinkin, L. The Influence of the Nervous System on Regeneration in Rhynchelmis Limosella Hof. J. Exp. Zool. 1936, 73, 43–65. [Google Scholar] [CrossRef]
- Sayles, L.P. Buds Induced by Implants of Anterior Nerve Cord and Neighboring Tissues Inserted at Various Levels in Clymenella Torquata. Biol. Bull. 1940, 78, 298–311. [Google Scholar] [CrossRef]
- Kiortsis, V.; Moraitou, M. Factors of regeneration in Spirographis spallanzani. In Regeneration in Animals and Related Problems; Kiortsis, V., Trampusch, H.A.L., Eds.; North Holland Publishing Co.: Amsterdam, The Netherlands, 1965; pp. 250–261. [Google Scholar]
- Combaz, A.; Boilly, B. Étude Expérimentale et Histologique de La Régénération Caudale En l’absence de Chaine Nerveuse Chez Les Nereidae (Annélides Polychètes). Ann. Embryol. Morphog. 1974, 7, 171–197. [Google Scholar]
- Pfannenstiel, H.-D. The Ventral Nerve Cord Signals Positional Information during Segment Formation in an Annelid (Ophryotrocha Puerilis, Polychaeta). Wilhelm Rouxs Arch. Dev. Biol. 1984, 194, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Zoran, M.J. Regeneration in Annelids. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; ISBN 978-0-470-01590-2. [Google Scholar]
- Sinigaglia, C.; Averof, M. The Multifaceted Role of Nerves in Animal Regeneration. Curr. Opin. Genet. Dev. 2019, 57, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Combaz, A.; Boilly-Marer, Y. Mise En Évidence de La Nature Dorsale de La Paroi de Corps Du Régénérat Aneurogénique de Nereis Pelagica Linné (Annélide Polychète). Comptes Rendus L’Académie Sci. 1976, 283, 785–788. [Google Scholar]
- Pirotte, N.; Leynen, N.; Artois, T.; Smeets, K. Do You Have the Nerves to Regenerate? The Importance of Neural Signalling in the Regeneration Process. Dev. Biol. 2016, 409, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Farkas, J.E.; Monaghan, J.R. A Brief History of the Study of Nerve Dependent Regeneration. Neurogenesis 2017, 4, e1302216. [Google Scholar] [CrossRef] [Green Version]
- Stocum, D.L. Nerves and Proliferation of Progenitor Cells in Limb Regeneration. Dev. Neurobiol. 2019, 79, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Ponesakki, V.; Paul, S.; Mani, D.K.S.; Rajendiran, V.; Kanniah, P.; Sivasubramaniam, S. Annotation of Nerve Cord Transcriptome in Earthworm Eisenia Fetida. Genom. Data 2017, 14, 91–105. [Google Scholar] [CrossRef]
- Bideau, L.; Kerner, P.; Hui, J.; Vervoort, M.; Gazave, E. Animal Regeneration in the Era of Transcriptomics. Cell. Mol. Life Sci. 2021, 78, 3941–3956. [Google Scholar] [CrossRef]
- Nagabhushanam, R.; Hanumante, M.M. Influence of Neurohumors on Caudal Regeneration of the Earthworm Perionyx Excavatus Perrier. Indian J. Exp. Biol. 1977, 15, 156. [Google Scholar]
- Vargas, J.; Alfaro-Rodríguez, A.; Perez-Orive, J.; Vargas, J.; Alfaro-Rodríguez, A.; Perez-Orive, J. Serotonin Induces or Inhibits Neuritic Regeneration of Leech CNS Neurons Depending on Neuronal Identity. Braz. J. Med. Biol. Res. 2019, 52. [Google Scholar] [CrossRef]
- Durchon, M. Neurosecretion and Hormonal Control of Reproduction in Annelida. Gen. Comp. Endocrinol. 1962, 1, 227–240. [Google Scholar] [CrossRef]
- Olive, P.J.W. Cellular Aspects of Regeneration Hormone Influence in Nereis Diversicolor. J. Embryol. Exp. Morphol. 1974, 32, 111–131. [Google Scholar]
- Hofmann, D.K. Regeneration and Endocrinology in the Polychaete Platynereis Dumerilii. Wilhelm Rouxs Arch. Dev. Biol. 1976, 180, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Schenk, S.; Krauditsch, C.; Frühauf, P.; Gerner, C.; Raible, F. Discovery of Methylfarnesoate as the Annelid Brain Hormone Reveals an Ancient Role of Sesquiterpenoids in Reproduction. eLife 2016, 5, e17126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Campos, P.; Kenny, N.J.; Verdes, A.; Fernández, R.; Novo, M.; Giribet, G.; Riesgo, A. Delegating Sex: Differential Gene Expression in Stolonizing Syllids Uncovers the Hormonal Control of Reproduction. Genome Biol. Evol. 2019, 11, 295–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreatta, G.; Broyart, C.; Borghgraef, C.; Vadiwala, K.; Kozin, V.; Polo, A.; Bileck, A.; Beets, I.; Schoofs, L.; Gerner, C.; et al. Corazonin Signaling Integrates Energy Homeostasis and Lunar Phase to Regulate Aspects of Growth and Sexual Maturation in Platynereis. Proc. Natl. Acad. Sci. USA 2020, 117, 1097–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dales, R.P. Defence mechanisms. In Physiology of Annelida; Mill, P.J., Ed.; Academic Press: London, UK; New York, NY, USA, 1978; pp. 479–507. [Google Scholar]
- LeGore, R.S.; Sparks, A.K. Repair of Body Wall Incision in the Rhynchobdellid Leech Piscicola Salmositica. J. Invertebr. Pathol. 1971, 18, 40–45. [Google Scholar] [CrossRef]
- Grimaldi, A.; Banfi, S.; Bianchi, C.; Greco, G.; Tettamanti, G.; Noonan, D.M.; Valvassori, R.; de Eguileor, M. The Leech: A Novel Invertebrate Model for Studying Muscle Regeneration and Diseases. Curr. Pharm. Des. 2010, 16, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.D. Origin of the Regeneration Blastema in Polychaete Annelids. Am. Zool. 1970, 10, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Bartolomaeus, T. On the Ultrastructure of the Coelomic Lining in the Annelida, Sipuncula and Echiura. Microfauna Mar. 1994, 9, 171–220. [Google Scholar]
- Linthicum, D.S.; Marks, D.H.; Stein, E.A.; Cooper, E.L. Graft Rejection in Earthworms: An Electron Microscopic Study. Eur. J. Immunol. 1977, 7, 871–876. [Google Scholar] [CrossRef]
- Stein, E.A.; Cooper, E.L. Inflammatory Responses in Annelids. Am. Zool. 1983, 23, 145–156. [Google Scholar] [CrossRef]
- Tettamanti, G.; Grimaldi, A.; Rinaldi, L.; Arnaboldi, F.; Congiu, T.; Valvassori, R.; Eguileor, M.D. The Multifunctional Role of Fibroblasts during Wound Healing in Hirudo Medicinalis (Annelida, Hirudinea). Biol. Cell 2004, 96, 443–455. [Google Scholar] [CrossRef]
- Boilly, B.; Boilly-Marer, Y.; Bely, A.E. Regulation of Dorso-Ventral Polarity by the Nerve Cord during Annelid Regeneration: A Review of Experimental Evidence. Regeneration 2017, 4, 54–68. [Google Scholar] [CrossRef]
- Raz, A.A.; Srivastava, M.; Reddien, P.W.; Salvamoser, R. Acoel Regeneration Mechanisms Indicate an Ancient Role for Muscle in Regenerative Patterning. Nat. Commun. 2017, 8, 1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rand, H.W. The Behavior of the Epidermis of the Earthworm in Regeneration. Arch. Für Entwickl. Org. 1905, 19, 16–57. [Google Scholar] [CrossRef]
- Clarck, M.E.; Clark, R.B. Growth and Regeneration in Nephtys. Zool. Physiol. 1962, 70, 24–90. [Google Scholar]
- Burke, D.J.M. Wound Healing in Eisenia Foetida (Oligochaeta). III. A Fine Structural Study of the Role of Non-Epidermal Tissues. Cell Tissue Res. 1974, 154, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Emberts, Z.; Escalante, I.; Bateman, P.W. The Ecology and Evolution of Autotomy. Biol. Rev. 2019, 94, 1881–1896. [Google Scholar] [CrossRef]
- Lesiuk, N.M.; Drewes, C.D. Autotomy Reflex in a Freshwater Oligochaete, Lumbriculus Variegatus (Clitellata: Lumbriculidae). Hydrobiologia 1999, 406, 253–261. [Google Scholar] [CrossRef]
- Kawamoto, S.; Yoshida-Noro, C.; Tochinai, S. Bipolar Head Regeneration Induced by Artificial Amputation in Enchytraeus Japonensis (Annelida, Oligochaeta). J. Exp. Zoolog. A Comp. Exp. Biol. 2005, 303A, 615–627. [Google Scholar] [CrossRef]
- Boilly, B. Origin of regeneration cells in Aricia foetida Clap. (Polychaete Annelid). Arch. Anat. Microsc. Morphol. Exp. 1968, 57, 297–308. [Google Scholar]
- Park, S.K.; Cho, S.-J.; Park, S.C. Histological Observations of Blastema Formation during Earthworm Tail Regeneration. Invertebr. Reprod. Dev. 2012, 57, 165–169. [Google Scholar] [CrossRef]
- Zattara, E.E. Regeneration, Fission and the Evolution of Developmental Novelty in Naid Annelids. Ph.D. Thesis, University of Maryland, College Park, MA, USA, 2012. [Google Scholar]
- Weidhase, M.; Bleidorn, C.; Beckers, P.; Helm, C. Myoanatomy and Anterior Muscle Regeneration of the Fireworm Eurythoe Cf. Complanata (Annelida: Amphinomidae). J. Morphol. 2016, 277, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Lindner, E.; Fischer, A. Zur Feinstruktur nereider und heteronereider Muskulatur von Platynereis dumerillii. Naturwissenschaften 1964, 51, 410. [Google Scholar] [CrossRef]
- Wissocq, J.C. Etude ultrastructurale de l’évolution des muscles longitudinaux lors de la stolonisation expérimentale de Syllis amica (Quatrefages) (Annélide Polychète). Z. Für Zellforsch. Mikrosk. Anat. 1967, 83, 449–467. [Google Scholar] [CrossRef]
- Pfeifer, K.; Schaub, C.; Wolfstetter, G.; Dorresteijn, A. Identification and Characterization of a Twist Ortholog in the Polychaete Annelid Platynereis Dumerilii Reveals Mesodermal Expression of Pdu-Twist. Dev. Genes Evol. 2013, 223, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Kozin, V.V.; Kostyuchenko, R.P. Evolutionary Conservation and Variability of the Mesoderm Development in Spiralia: A Peculiar Pattern of Nereid Polychaetes. Biol. Bull. 2016, 43, 216–225. [Google Scholar] [CrossRef]
- Myohara, M.; Yoshida-noro, C.; Kobari, F. Fragmenting oligochaete Enchytraeus japonensis: A new material for regeneration study. Dev. Growth Differ. 1999, 41, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Tweeten, K.A.; Reiner, A. Characterization of serine proteases of Lumbriculus variegatus and their role in regeneration. Invertebr. Biol. 2012, 131, 322–332. [Google Scholar] [CrossRef]
- Boilly, B. Sur la régénération d’un intestin dans la zone pharyngienne chez Syllis amica Quatrefages (Annélide Polychète). Cah. Biol. Mar. 1967, 8, 221–231. [Google Scholar]
- Boilly, B. Origine des cellules régénératrices chez Nereis diversicolor O. F. Müller (Annélide Polychète). Wilhelm Roux. Arch. Entwickl. Mech. Org. 1969, 162, 286–305. [Google Scholar] [CrossRef]
- Okada, Y.K. Regeneration and fragmentation in the Syllidian Polychaetes. Wilhelm Roux. Arch. Entwickl. 1929, 115, 542–600. [Google Scholar] [CrossRef] [PubMed]
- Bely, A.E.; Wray, G.A. Evolution of regeneration and fission in annelids: Insights from engrailed- and orthodenticle-class gene expression. Development 2001, 128, 2781–2791. [Google Scholar] [CrossRef] [PubMed]
- Takeo, M.; Yoshida-Noro, C.; Tochinai, S. Morphallactic regeneration as revealed by region-specific gene expression in the digestive tract of Enchytraeus japonensis (Oligochaeta, Annelida). Dev. Dyn. 2008, 237, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Özpolat, B.D.; Sloane, E.S.; Zattara, E.E.; Bely, A.E. Plasticity and regeneration of gonads in the annelid Pristina leidyi. EvoDevo 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Sampling reference | [22] | [22] | [25] | [23] | [24] | [29] | [26] | [28] | [30] |
|---|---|---|---|---|---|---|---|---|---|
| anterior regeneration(AR) stages | 1, 3, 6, 8 dpa | 1, 5, 9, 12 dpa | mixed-stage regenerating and fissioning worms | mixed AR and PR samples (2, 3, 4, 11, 14, 16, 17, 18, 21, 22, 26, 27, 34, 38, 41, 47, 51, 53, 58 hpa) | mixed-stage (0.5, 1, 3, 6, 12, 18, and 24 hpa) | 4, 6 dpa | not sampled | 72 hpa (single-cell RNA-Seq) | 24, 48, 72 hpa |
| posterior regeneration(PR) stages | 1, 3, 6, 8 dpa | 1, 5, 9, 12 dpa | not sampled | not sampled | 15, 20, 30 dpa | 0, 6, 12, 24, 48, 72 hpa (RNA-Seq) | not sampled | ||
| time of complete regeneration in the experiments, temperature | complete AR after 14 dpa, 14 °C | incomplete prostomium after 14 dpa, 14 °C | no data | 4–5 dpa, 24 °C | no data, 22–24 °C | no data | no data, 22 °C | after 18 dpa, 25 °C | no data, 20 °C |
| technique | RNA-Seq | RNA-Seq | 454 pyrosequencing | suppression subtractive hybridization | expressed sequence tags (ESTs) | RNA-Seq | RNA-Seq | single-cell RNA-Seq, RNA-Seq | RNA-Seq |
| number of identified/assembled sequences | 526,860 contigs | 315,224 contigs | 111,201 unigenes | 803 genes | 105,193 contigs | 125,896 contigs | up to 233,332 contigs | 164,769 transcripts | |
| number of specifically upregulated genes | 1940 (AR) and 33 (PR) differentially expressed genes | 4699 (AR) and 161 (PR) differentially expressed genes | no data | 165 | no data | 3986 | 3589, 1887, 617 (15, 20, 30 dpa) | 6048 differentially expressed genes | 111 |
| number of specifically downregulated genes | no data | no data | no data | 6882 | 5124, 3612, 614 (15, 20, 30 dpa) | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostyuchenko, R.P.; Kozin, V.V. Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration. Genes 2021, 12, 1148. https://doi.org/10.3390/genes12081148
Kostyuchenko RP, Kozin VV. Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration. Genes. 2021; 12(8):1148. https://doi.org/10.3390/genes12081148
Chicago/Turabian StyleKostyuchenko, Roman P., and Vitaly V. Kozin. 2021. "Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration" Genes 12, no. 8: 1148. https://doi.org/10.3390/genes12081148
APA StyleKostyuchenko, R. P., & Kozin, V. V. (2021). Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration. Genes, 12(8), 1148. https://doi.org/10.3390/genes12081148