
The Tentacular Spectacular: Evolution of Regeneration in Sea Anemones


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. What Is Regeneration?
Genetic Framework of Regeneration: Key Players Identified from Model Organisms
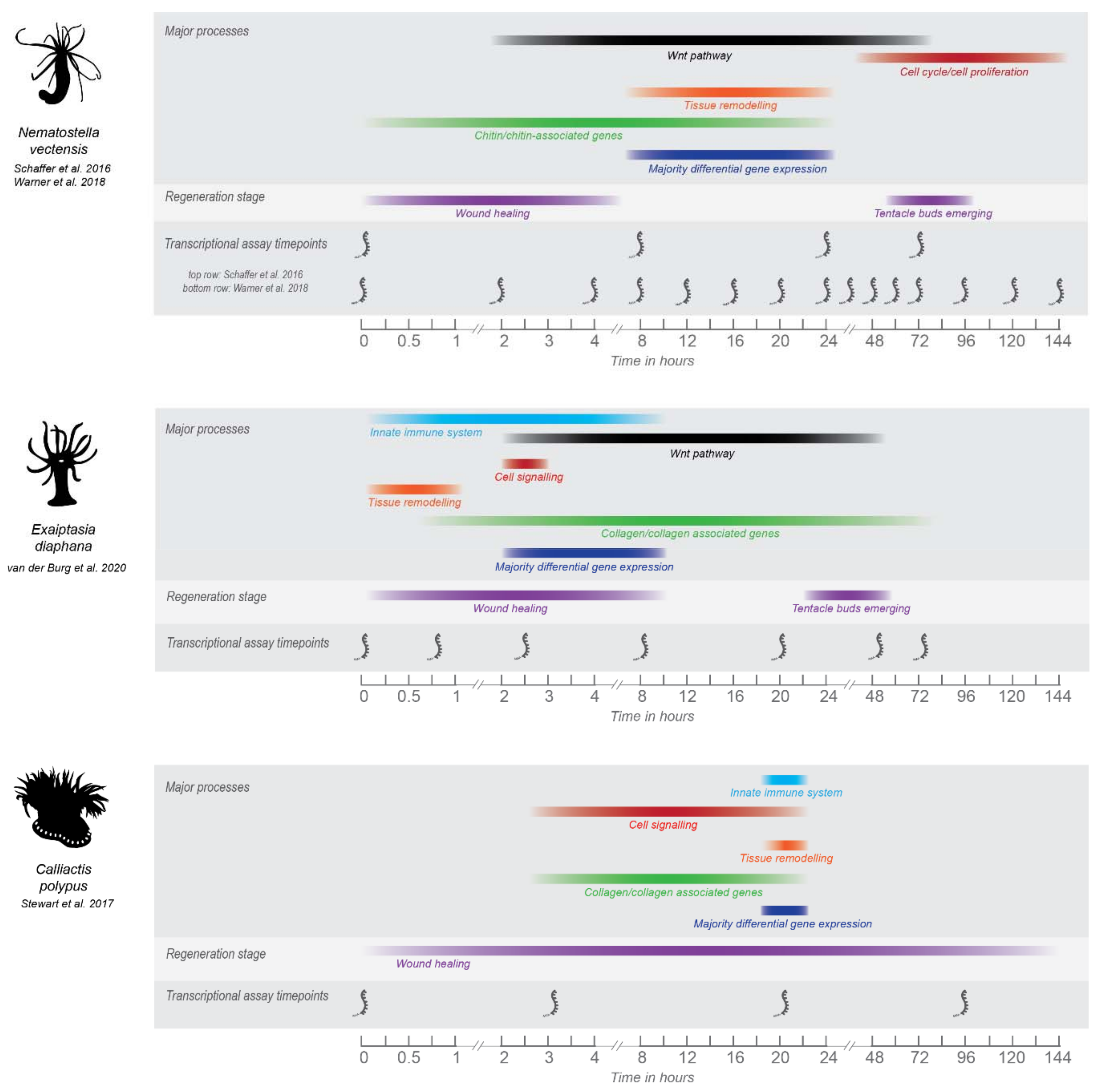
3. Overview of Sea Anemone Molecular Regeneration Studies
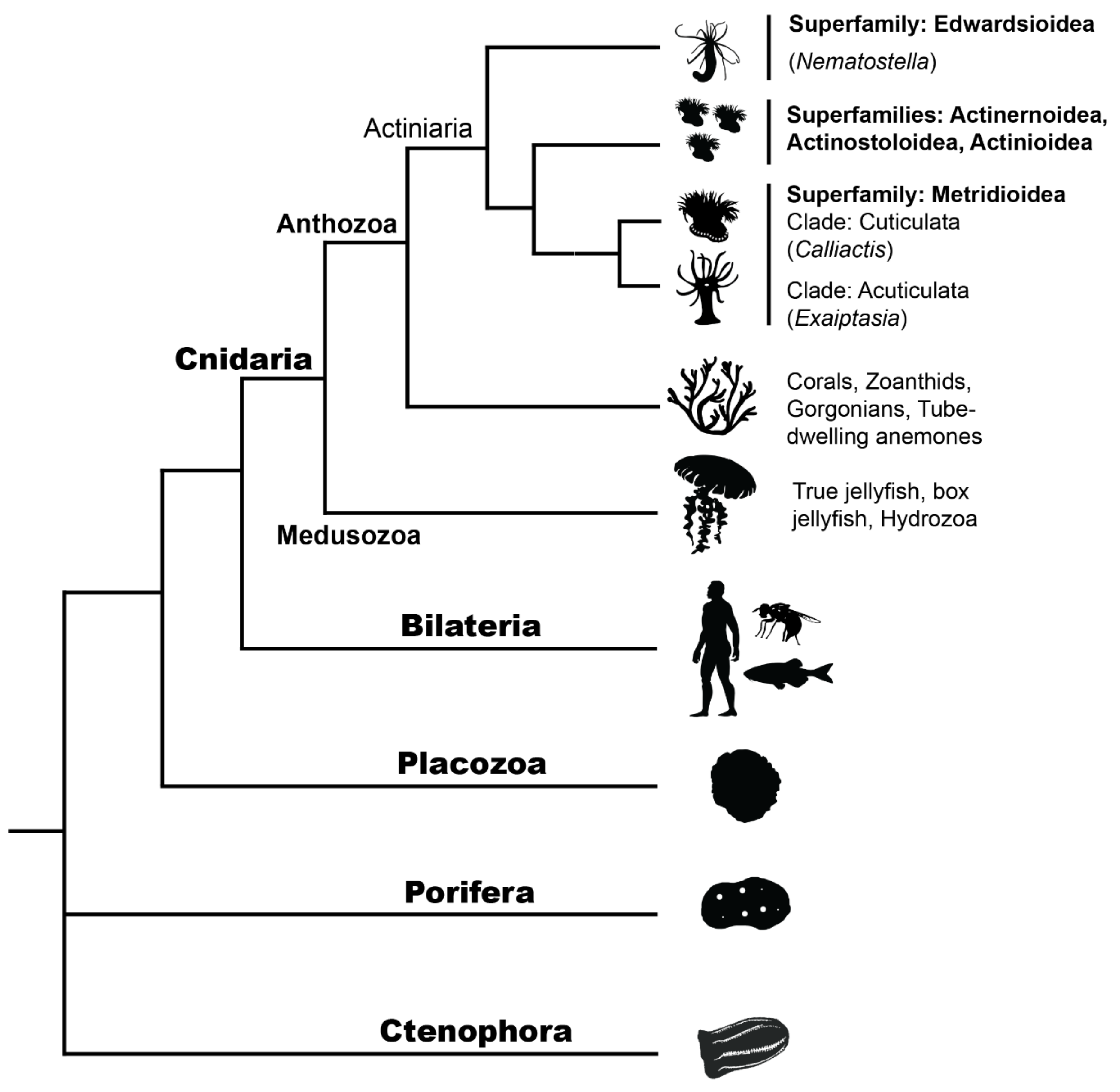
4. Ecological, Evolutionary and Genomic Context of Sea Anemone Regeneration
4.1. Linking Life History Strategies and Regeneration
4.2. Genomic Resources for Actiniaria
4.3. Regeneration Gene Set Evolution
4.4. Broad-Scale Evolution of Regeneration
4.5. Regeneration and the Immune System
4.6. Is Regeneration a Recapitulation of Development?
4.7. Other Observations
4.8. Conclusions and Future Areas of Exploration
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, E.; Hwang, D.-S.; Lee, J.-S.; Song, J.-I.; Seo, T.-K.; Won, Y.-J. Estimation of Divergence Times in Cnidarian Evolution Based on Mitochondrial Protein-Coding Genes and the Fossil Record. Mol. Phylogenet. Evol. 2012, 62, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.J.; Cotton, J.A.; Gehling, J.G.; Pisani, D. The Ediacaran Emergence of Bilaterians: Congruence between the Genetic and the Geological Fossil Records. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1435–1443. [Google Scholar] [CrossRef]
- Zapata, F.; Goetz, F.E.; Smith, S.A.; Howison, M.; Siebert, S.; Church, S.; Sanders, S.M.; Ames, C.L.; McFadden, C.S.; France, S.C.; et al. Phylogenomic Analyses Support Traditional Relationships within Cnidaria. bioRxiv 2015, 017632. [Google Scholar] [CrossRef] [Green Version]
- Kusserow, A.; Pang, K.; Sturm, C.; Hrouda, M.; Lentfer, J.; Schmidt, H.A.; Technau, U.; von Haeseler, A.; Hobmayer, B.; Martindale, M.Q.; et al. Unexpected Complexity of the Wnt Gene Family in a Sea Anemone. Nature 2005, 433, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J.; Ball, E.E.; Technau, U. Cnidarians and Ancestral Genetic Complexity in the Animal Kingdom. Trends Genet. 2005, 21, 536–539. [Google Scholar] [CrossRef]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea Anemone Genome Reveals Ancestral Eumetazoan Gene Repertoire and Genomic Organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.F.; Burton, P.M.; Mazza, M.E.; Kwong, G.K.; Mullikin, J.C.; Finnerty, J.R. The Cnidarian-Bilaterian Ancestor Possessed at Least 56 Homeoboxes: Evidence from the Starlet Sea Anemone, Nematostella vectensis. Genome Biol. 2006, 7, R64. [Google Scholar] [CrossRef] [Green Version]
- Technau, U.; Rudd, S.; Maxwell, P.; Gordon, P.M.K.; Saina, M.; Grasso, L.C.; Hayward, D.C.; Sensen, C.W.; Saint, R.; Holstein, T.W.; et al. Maintenance of Ancestral Complexity and Non-Metazoan Genes in Two Basal Cnidarians. Trends Genet. 2005, 21, 633–639. [Google Scholar] [CrossRef]
- Trembley, A. Mémoires Pour Servir a l’histoire d’un Genre de Polypes d’eau Douce, a Bras En Forme de Cornes; Jean & Herman Verbeek: Leiden, The Netherlands, 1744. [Google Scholar]
- Fraune, S.; Abe, Y.; Bosch, T.C.G. Disturbing Epithelial Homeostasis in the Metazoan Hydra Leads to Drastic Changes in Associated Microbiota. Environ. Microbiol. 2009, 11, 2361–2369. [Google Scholar] [CrossRef]
- Fraune, S.; Bosch, T.C.G. Long-Term Maintenance of Species-Specific Bacterial Microbiota in the Basal Metazoan Hydra. Proc. Natl. Acad. Sci. USA 2007, 104, 13146–13151. [Google Scholar] [CrossRef] [Green Version]
- Babonis, L.S.; Martindale, M.Q. Phylogenetic Evidence for the Modular Evolution of Metazoan Signalling Pathways. Philos. Trans. R. Soc. B 2017, 372, 20150477. [Google Scholar] [CrossRef] [Green Version]
- He, S.; del Viso, F.; Chen, C.-Y.; Ikmi, A.; Kroesen, A.E.; Gibson, M.C. An Axial Hox Code Controls Tissue Segmentation and Body Patterning in Nematostella vectensis. Science 2018, 361, 1377–1380. [Google Scholar] [CrossRef] [Green Version]
- Technau, U.; Steele, R.E. Evolutionary Crossroads in Developmental Biology: Cnidaria. Development 2011, 138, 1447–1458. [Google Scholar] [CrossRef] [Green Version]
- Tiozzo, S.; Copley, R.R. Reconsidering Regeneration in Metazoans: An Evo-Devo Approach. Front. Ecol. Evol. 2015, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C.G.; Augustin, R.; Anton-Erxleben, F.; Fraune, S.; Hemmrich, G.; Zill, H.; Rosenstiel, P.; Jacobs, G.; Schreiber, S.; Leippe, M.; et al. Uncovering the Evolutionary History of Innate Immunity: The Simple Metazoan Hydra Uses Epithelial Cells for Host Defence. Dev. Comp. Immunol. 2009, 33, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Hemmrich, G.; Miller, D.J.; Bosch, T.C.G. The Evolution of Immunity: A Low-Life Perspective. Trends Immunol. 2007, 28, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Sharoni, T.; Admoni, Y.; Aharoni, R.; Moran, Y. Functional Characterization of the Cnidarian Antiviral Immune Response Reveals Ancestral Complexity. Mol. Biol. Evol. 2021, msab197. [Google Scholar] [CrossRef]
- Poole, A.Z.; Weis, V.M. TIR-Domain-Containing Protein Repertoire of Nine Anthozoan Species Reveals Coral–Specific Expansions and Uncharacterized Proteins. Dev. Comp. Immunol. 2014, 46, 480–488. [Google Scholar] [CrossRef]
- Van der Burg, C.A.; Prentis, P.J.; Surm, J.M.; Pavasovic, A. Insights into the Innate Immunome of Actiniarians Using a Comparative Genomic Approach. BMC Genom. 2016, 17, 850. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.; Barbeitos, M.S.; Brugler, M.R.; Crowley, L.M.; Grajales, A.; Gusmão, L.; Häussermann, V.; Reft, A.; Daly, M. Hidden among Sea Anemones: The First Comprehensive Phylogenetic Reconstruction of The Order Actiniaria (Cnidaria, Anthozoa, Hexacorallia) Reveals a Novel Group of Hexacorals. PLoS ONE 2014, 9, e96998. [Google Scholar] [CrossRef] [PubMed]
- McFadden, C.S.; Quattrini, A.M.; Brugler, M.R.; Cowman, P.F.; Dueñas, L.F.; Kitahara, M.V.; Paz-García, D.A.; Reimer, J.D.; Rodríguez, E. Phylogenomics, Origin, and Diversification of Anthozoans (Phylum cnidaria). Syst. Biol. 2021. [Google Scholar] [CrossRef]
- Blair, J.E.; Hedges, S.B. Molecular Phylogeny and Divergence Times of Deuterostome Animals. Mol. Biol. Evol. 2005, 22, 2275–2284. [Google Scholar] [CrossRef] [Green Version]
- Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D.; Peterson, K.J. The Cambrian Conundrum: Early Divergence and Later Ecological Success in the Early History of Animals. Science 2011, 334, 1091–1097. [Google Scholar] [CrossRef] [Green Version]
- Goodheart, J.A.; Bely, A.E. Sequestration of Nematocysts by Divergent Cnidarian Predators: Mechanism, Function, and Evolution. Invertebr. Biol. 2017, 136, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Ottaway, J.R. Predators of Sea Anemones. Tuatara 1977, 22, 213–221. [Google Scholar]
- Edmunds, M.; Potts, G.W.; Swinfen, R.C.; Waters, V.L. Defensive Behaviour of Sea Anemones in Response to Predation by the Opisthobranch Mollusc Aeolidia papillosa (L.). J. Mar. Biol. Assoc. UK 1976, 56, 65–83. [Google Scholar] [CrossRef]
- Lawn, I.D.; Ross, D.M.; Denton, E.J. The Behavioural Physiology of the Swimming Sea Anemone Boloceroides mcmurrichi. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1982, 216, 315–334. [Google Scholar] [CrossRef]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of Venom Composition across a Complex Life Cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef] [PubMed]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surm, J.M.; Smith, H.L.; Madio, B.; Undheim, E.A.B.; King, G.F.; Hamilton, B.R.; van der Burg, C.A.; Pavasovic, A.; Prentis, P.J. A Process of Convergent Amplification and Tissue-Specific Expression Dominates the Evolution of Toxin and Toxin-like Genes in Sea Anemones. Mol. Ecol. 2019, 28, 2272–2289. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, P.G. Acquisition and Use of Nematocysts by Cnidarian Predators. Toxicon 2009, 54, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Dungan, A.M.; Hartman, L.M.; Tortorelli, G.; Belderok, R.; Lamb, A.M.; Pisan, L.; McFadden, G.I.; Blackall, L.L.; van Oppen, M.J.H. Exaiptasia diaphana from the Great Barrier Reef: A Valuable Resource for Coral Symbiosis Research. Symbiosis 2020, 80, 195–206. [Google Scholar] [CrossRef]
- Clayton, W.S. Pedal Laceration by the Anemone Aiptasia pallida. Mar. Ecol. Prog. Ser. 1985, 21, 75–80. [Google Scholar] [CrossRef]
- Schlesinger, A.; Kramarsky-Winter, E.; Rosenfeld, H.; Armoza-Zvoloni, R.; Loya, Y. Sexual Plasticity and Self-Fertilization in the Sea Anemone Aiptasia diaphana. PLoS ONE 2010, 5, e11874. [Google Scholar] [CrossRef]
- Bocharova, E.S.; Kozevich, I.A. Modes of Reproduction in Sea Anemones (Cnidaria, Anthozoa). Biol Bull. Russ. Acad. Sci. 2011, 38, 849–860. [Google Scholar] [CrossRef]
- Layden, M.J.; Rentzsch, F.; Röttinger, E. The Rise of the Starlet Sea Anemone Nematostella vectensis as a Model System to Investigate Development and Regeneration. WIREs Dev. Biol. 2016, 5, 408–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebens, K.P. Morphological Variability during Longitudinal Fission of the Intertidal Sea Anemone, Anthopleura elegantissima (Brandt). Pac. Sci. 1983, 37, 12. [Google Scholar]
- Schmidt, H. Anthopleura stellula (Actiniaria, Actiniidae) and Its Reproduction by Transverse Fission. Mar. Biol. 1970, 5, 245–255. [Google Scholar] [CrossRef]
- Young, J. The Nature of Tissue Regeneration after Wounding in the Sea Anemone Calliactis parasitica (Couch). J. Mar. Biol. Ass. UK 1974, 54, 599–617. [Google Scholar] [CrossRef]
- Shick, J.M.; Lamb, A.N. Asexual Reproduction and Genetic Population Structure in the Colonizing Sea Anemone Haliplanella luciae. Biol. Bull. 1977, 153, 604–617. [Google Scholar] [CrossRef]
- Singer, I.I. Tentacular and Oral-Disc Regeneration in the Sea Anemone, Aiptasia diaphana III. J. Embryol. Exp. Morphol. 1971, 26, 253–270. [Google Scholar]
- Singer, I.I. An Electron Microscopic and Autoradiographic Study of Mesogleal Organization and Collagen Synthesis in the Sea Anemone Aiptasia diaphana. Cell Tissue Res. 1974, 149, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Singer, I.I.; Palmer, J.D. Tentacular and Oral-Disc Regeneration in the Sea Anemone, Aiptasia diaphana. I. Sequential Morphological Events in Distal-End Restitution. J. Morphol. 1969, 127, 373–381. [Google Scholar] [CrossRef]
- Child, C.M. Factors of Form Regulation in Harenactis attenuata. I. Wound Reaction and Restitution in General and the Regional Factors in Oral Restitution. J. Exp. Zool. 1909, 6, 471–506. [Google Scholar] [CrossRef]
- Bucklin, A. Biochemical Genetic Variation, Growth and Regeneration of the Sea Anemone, Metridium, of British Shores. J. Mar. Biol. Assoc. UK 1985, 65, 141–157. [Google Scholar] [CrossRef]
- Pearse, V.B. Prodigies of Propagation: The Many Modes of Clonal Replication in Boloceroidid Sea Anemones (Cnidaria, Anthozoa, Actiniaria). Invertebr. Reprod. Dev. 2002, 41, 201–213. [Google Scholar] [CrossRef]
- Alvarado, A.S.; Tsonis, P.A. Bridging the Regeneration Gap: Genetic Insights from Diverse Animal Models. Nat. Rev. Genet. 2006, 7, 873–884. [Google Scholar] [CrossRef]
- Brockes, J.P.; Kumar, A.; Velloso, C.P. Regeneration as an Evolutionary Variable. J. Anat. 2001, 199, 3–11. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Selck, C.; Friedrich, B.; Lutz, R.; Vila-Farré, M.; Dahl, A.; Brandl, H.; Lakshmanaperumal, N.; Henry, I.; Rink, J.C. Reactivating Head Regrowth in a Regeneration-Deficient Planarian Species. Nature 2013, 500, 81–84. [Google Scholar] [CrossRef]
- Poss, K.D. Advances in Understanding Tissue Regenerative Capacity and Mechanisms in Animals. Nat. Rev. Genet. 2010, 11, 710–722. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, E.M.; Reddien, P.W. The Cellular Basis for Animal Regeneration. Dev. Cell 2011, 21, 172–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockes, J.P.; Kumar, A. Comparative Aspects of Animal Regeneration. Annu. Rev. Cell Dev. Biol. 2008, 24, 525–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound Repair and Regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Darnet, S.; Dragalzew, A.C.; Amaral, D.B.; Sousa, J.F.; Thompson, A.W.; Cass, A.N.; Lorena, J.; Pires, E.S.; Costa, C.M.; Sousa, M.P.; et al. Deep Evolutionary Origin of Limb and Fin Regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 15106–15115. [Google Scholar] [CrossRef] [Green Version]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The Zebrafish as a Model for Complex Tissue Regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Guzmán, T.; Wang, H.; Khattak, S.; Schuez, M.; Roensch, K.; Nacu, E.; Tazaki, A.; Joven, A.; Tanaka, E.M.; Simon, A. Fundamental Differences in Dedifferentiation and Stem Cell Recruitment during Skeletal Muscle Regeneration in Two Salamander Species. Cell Stem Cell 2014, 14, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, B.; Thompson, K.; Frank, U. Distinct Mechanisms Underlie Oral vs. Aboral Regeneration in the Cnidarian Hydractinia echinata. eLife 2015, 4, e05506. [Google Scholar] [CrossRef]
- Guedelhoefer, O.C.; Alvarado, A.S. Amputation Induces Stem Cell Mobilization to Sites of Injury during Planarian Regeneration. Development 2012, 139, 3510–3520. [Google Scholar] [CrossRef] [Green Version]
- Henry, J.Q.; Martindale, M.Q. Regulation and Regeneration in the Ctenophore Mnemiopsis leidyi. Dev. Biol. 2000, 227, 720–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK Cascades: Signaling Components, Nuclear Roles and Mechanisms of Nuclear Translocation. Biochim. Biophys. Acta 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [Green Version]
- Cary, G.A.; Wolff, A.; Zueva, O.; Pattinato, J.; Hinman, V.F. Analysis of Sea Star Larval Regeneration Reveals Conserved Processes of Whole-Body Regeneration across the Metazoa. BMC Biol. 2019, 17, 16. [Google Scholar] [CrossRef] [Green Version]
- DuBuc, T.Q.; Traylor-Knowles, N.; Martindale, M.Q. Initiating a Regenerative Response; Cellular and Molecular Features of Wound Healing in the Cnidarian Nematostella vectensis. BMC Biol. 2014, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A. Evolution of Immune Pathways in Regeneration and Repair: Recent Concepts and Translational Perspectives. Semin. Immunol. 2014, 26, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Leiper, L.J.; Walczysko, P.; Kucerova, R.; Ou, J.; Shanley, L.J.; Lawson, D.; Forrester, J.V.; McCaig, C.D.; Zhao, M.; Collinson, J.M. The Roles of Calcium Signaling and ERK1/2 Phosphorylation in a Pax6+/− Mouse Model of Epithelial Wound-Healing Delay. BMC Biol. 2006, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H.O.; Höger, S.K.; Looso, M.; Lengfeld, T.; Kuhn, A.; Warnken, U.; Nishimiya-Fujisawa, C.; Schnölzer, M.; Krüger, M.; Özbek, S.; et al. A Comprehensive Transcriptomic and Proteomic Analysis of Hydra Head Regeneration. Mol. Biol. Evol. 2015, 32, 1928–1947. [Google Scholar] [CrossRef]
- Yoo, S.K.; Freisinger, C.M.; LeBert, D.C.; Huttenlocher, A. Early Redox, Src Family Kinase, and Calcium Signaling Integrate Wound Responses and Tissue Regeneration in Zebrafish. J. Cell Biol. 2012, 199, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Chera, S.; Ghila, L.; Dobretz, K.; Wenger, Y.; Bauer, C.; Buzgariu, W.; Martinou, J.-C.; Galliot, B. Apoptotic Cells Provide an Unexpected Source of Wnt3 Signaling to Drive Hydra Head Regeneration. Dev. Cell 2009, 17, 279–289. [Google Scholar] [CrossRef]
- Clevers, H.; Loh, K.M.; Nusse, R. An Integral Program for Tissue Renewal and Regeneration: Wnt Signaling and Stem Cell Control. Science 2014, 346, 1248012. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.J.; Plickert, G.; Kuenzel, T.; Tilmann, W.; Frank, U. Wnt Signaling Promotes Oral but Suppresses Aboral Structures in Hydractinia Metamorphosis and Regeneration. Development 2010, 137, 3057–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gufler, S.; Artes, B.; Bielen, H.; Krainer, I.; Eder, M.-K.; Falschlunger, J.; Bollmann, A.; Ostermann, T.; Valovka, T.; Hartl, M.; et al. β-Catenin Acts in a Position-Independent Regeneration Response in the Simple Eumetazoan Hydra. Dev. Biol. 2018, 433, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Wikramanayake, A.H.; Hong, M.; Lee, P.N.; Pang, K.; Byrum, C.A.; Bince, J.M.; Xu, R.; Martindale, M.Q. An Ancient Role for Nuclear β-Catenin in the Evolution of Axial Polarity and Germ Layer Segregation. Nature 2003, 426, 446–450. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Bazarsky, M.; Levy, K.; Chalifa-Caspi, V.; Gat, U. A Transcriptional Time-Course Analysis of Oral vs. Aboral Whole-Body Regeneration in the Sea Anemone Nematostella vectensis. BMC Genom. 2016, 17, 718. [Google Scholar] [CrossRef] [Green Version]
- Love, N.R.; Chen, Y.; Ishibashi, S.; Kritsiligkou, P.; Lea, R.; Koh, Y.; Gallop, J.L.; Dorey, K.; Amaya, E. Amputation-Induced Reactive Oxygen Species Are Required for Successful Xenopus Tadpole Tail Regeneration. Nat. Cell Biol. 2013, 15, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Warner, J.F.; Amiel, A.R.; Johnston, H.; Röttinger, E. Regeneration Is a Partial Redeployment of the Embryonic Gene Network. bioRxiv 2019, 658930. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, M.R.; Zapperi, S.; Porta, C.A.M.L. Regeneration in Distantly Related Species: Common Strategies and Pathways. NPJ Syst. Biol. Appl. 2018, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A Tissue-Scale Gradient of Hydrogen Peroxide Mediates Rapid Wound Detection in Zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef]
- Stewart, Z.K.; Pavasovic, A.; Hock, D.H.; Prentis, P.J. Transcriptomic Investigation of Wound Healing and Regeneration in the Cnidarian Calliactis polypus. Sci. Rep. 2017, 7, 41458. [Google Scholar] [CrossRef]
- Wenger, Y.; Buzgariu, W.; Reiter, S.; Galliot, B. Injury-Induced Immune Responses in Hydra. Semin. Immunol. 2014, 26, 277–294. [Google Scholar] [CrossRef] [PubMed]
- Krens, S.F.G.; Spaink, H.P.; Snaar-Jagalska, B.E. Functions of the MAPK Family in Vertebrate-Development. FEBS Lett. 2006, 580, 4984–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.; Anavy, L.; Cole, A.G.; Winter, E.; Mostov, N.; Khair, S.; Senderovich, N.; Kovalev, E.; Silver, D.H.; Feder, M.; et al. The Mid-Developmental Transition and the Evolution of Animal Body Plans. Nature 2016, 531, 637–641. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Tautz, D.; Domazet-Lošo, T. The Evolutionary Origin of Orphan Genes. Nat. Rev. Genet. 2011, 12, 692–702. [Google Scholar] [CrossRef]
- Echeverri, K.; Tanaka, E.M. Proximodistal Patterning during Limb Regeneration. Dev. Biol. 2005, 279, 391–401. [Google Scholar] [CrossRef]
- Garza-Garcia, A.; Harris, R.; Esposito, D.; Gates, P.B.; Driscoll, P.C. Solution Structure and Phylogenetics of Prod1, a Member of the Three-Finger Protein Superfamily Implicated in Salamander Limb Regeneration. PLoS ONE 2009, 4, e7123. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, S.M.; Gates, P.B.; Brockes, J.P. The Newt Ortholog of CD59 Is Implicated in Proximodistal Identity during Amphibian Limb Regeneration. Dev. Cell 2002, 3, 547–555. [Google Scholar] [CrossRef]
- Garza-Garcia, A.A.; Driscoll, P.C.; Brockes, J.P. Evidence for the Local Evolution of Mechanisms Underlying Limb Regeneration in Salamanders. Integr. Comp. Biol. 2010, 50, 528–535. [Google Scholar] [CrossRef] [Green Version]
- Babonis, L.S.; Martindale, M.Q.; Ryan, J.F. Do Novel Genes Drive Morphological Novelty? An Investigation of the Nematosomes in the Sea Anemone Nematostella vectensis. BMC Evol. Biol. 2016, 16, 114. [Google Scholar] [CrossRef] [Green Version]
- Forêt, S.; Knack, B.; Houliston, E.; Momose, T.; Manuel, M.; Quéinnec, E.; Hayward, D.C.; Ball, E.E.; Miller, D.J. New Tricks with Old Genes: The Genetic Bases of Novel Cnidarian Traits. Trends Genet. 2010, 26, 154–158. [Google Scholar] [CrossRef]
- Warner, J.F.; Guerlais, V.; Amiel, A.R.; Johnston, H.; Nedoncelle, K.; Röttinger, E. NvERTx: A Gene Expression Database to Compare Embryogenesis and Regeneration in the Sea Anemone Nematostella vectensis. Development 2018, 145, dev162867. [Google Scholar] [CrossRef] [Green Version]
- Van der Burg, C.A.; Pavasovic, A.; Gilding, E.K.; Pelzer, E.S.; Surm, J.M.; Smith, H.L.; Walsh, T.P.; Prentis, P.J. The Rapid Regenerative Response of a Model Sea Anemone Species Exaiptasia pallida Is Characterised by Tissue Plasticity and Highly Coordinated Cell Communication. Mar. Biotechnol. 2020, 22, 285–307. [Google Scholar] [CrossRef] [PubMed]
- Philipp, I.; Aufschnaiter, R.; Özbek, S.; Pontasch, S.; Jenewein, M.; Watanabe, H.; Rentzsch, F.; Holstein, T.W.; Hobmayer, B. Wnt/β-Catenin and Noncanonical Wnt Signaling Interact in Tissue Evagination in the Simple Eumetazoan Hydra. Proc. Natl. Acad. Sci. USA 2009, 106, 4290–4295. [Google Scholar] [CrossRef] [Green Version]
- Leclère, L.; Bause, M.; Sinigaglia, C.; Steger, J.; Rentzsch, F. Development of the Aboral Domain in Nematostella Requires β-Catenin and the Opposing Activities of Six3/6 and Frizzled5/8. Development 2016, 143, 1766–1777. [Google Scholar] [CrossRef] [Green Version]
- Trevino, M.; Stefanik, D.J.; Rodriguez, R.; Harmon, S.; Burton, P.M. Induction of Canonical Wnt Signaling by Alsterpaullone Is Sufficient for Oral Tissue Fate during Regeneration and Embryogenesis in Nematostella vectensis. Dev. Dyn. 2011, 240, 2673–2679. [Google Scholar] [CrossRef] [Green Version]
- Abrams, M.J.; Basinger, T.; Yuan, W.; Guo, C.-L.; Goentoro, L. Self-Repairing Symmetry in Jellyfish through Mechanically Driven Reorganization. Proc. Natl. Acad. Sci. USA 2015, 112, E3365–E3373. [Google Scholar] [CrossRef] [Green Version]
- Van de Water, J.A.J.M.; Ainsworth, T.D.; Leggat, W.; Bourne, D.G.; Willis, B.L.; van Oppen, M.J.H. The Coral Immune Response Facilitates Protection against Microbes during Tissue Regeneration. Mol. Ecol. 2015, 24, 3390–3404. [Google Scholar] [CrossRef]
- Horricks, R.A.; Herbinger, C.M.; Vickaryous, M.K.; Taylor, P.; Lumsden, J.S. Differential Protein Abundance Associated with Delayed Regeneration of the Scleractinian Coral Montastraea cavernosa. Coral Reefs 2020, 39, 1175–1186. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Piraino, S.; Miglietta, M.P. Transcriptome Characterization of Reverse Development in Turritopsis dohrnii (Hydrozoa, Cnidaria). G3 Genes Genomes Genet. 2019, 9, 4127–4138. [Google Scholar] [CrossRef] [Green Version]
- Morgan, T.H. Regeneration in the Egg, Embryo, and Adult. Am. Nat. 1901, 35, 949–973. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.A.; Kirkness, E.F.; Simakov, O.; Hampson, S.E.; Mitros, T.; Weinmaier, T.; Rattei, T.; Balasubramanian, P.G.; Borman, J.; Busam, D.; et al. The Dynamic Genome of Hydra. Nature 2010, 464, 592–596. [Google Scholar] [CrossRef]
- Park, H.D.; Ortmeyer, A.B.; Blankenbaker, D.P. Cell Division during Regeneration in Hydra. Nature 1970, 227, 617–619. [Google Scholar] [CrossRef]
- Cummings, S.G.; Bode, H.R. Head Regeneration and Polarity Reversal in Hydra Attenuata Can Occur in the Absence of DNA Synthesis. Wilhelm Roux’ Archiv. 1984, 194, 79–86. [Google Scholar] [CrossRef]
- Gierer, A.; Berking, S.; Bode, H.; David, C.N.; Flick, K.; Hansmann, G.; Schaller, H.; Trenkner, E. Regeneration of Hydra from Reaggregated Cells. Nat. New Biol. 1972, 239, 98–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitzel, A.M.; Burton, P.M.; Krone, C.; Finnerty, J.R. Comparison of Developmental Trajectories in the Starlet Sea Anemone Nematostella vectensis: Embryogenesis, Regeneration, and Two Forms of Asexual Fission. Invertebr. Biol. 2007, 126, 99–112. [Google Scholar] [CrossRef]
- Hand, C.; Uhlinger, K.R. Asexual Reproduction by Transverse Fission and Some Anomalies in the Sea Anemone Nematostella vectensis. Invertebr. Biol. 1995, 114, 9–18. [Google Scholar] [CrossRef]
- Darling, J.A.; Reitzel, A.R.; Burton, P.M.; Mazza, M.E.; Ryan, J.F.; Sullivan, J.C.; Finnerty, J.R. Rising Starlet: The Starlet Sea Anemone, Nematostella vectensis. Bioessays 2005, 27, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, D.J.; Xiang, Y.; Pettay, D.T.; Zhong, M.; Santos, S.R. Population Genetic Data of a Model Symbiotic Cnidarian System Reveal Remarkable Symbiotic Specificity and Vectored Introductions across Ocean Basins. Mol. Ecol. 2013, 22, 4499–4515. [Google Scholar] [CrossRef] [PubMed]
- Brooks, W.R.; Mariscal, R.N. Asexual Reproduction by the Symbiotic Sea Anemone Calliactis Tricolor (Lesueur). Bull. Mar. Sci. 1985, 36, 432–435. [Google Scholar]
- Bossert, P.E.; Dunn, M.P.; Thomsen, G.H. A Staging System for the Regeneration of a Polyp from the Aboral Physa of the Anthozoan Cnidarian Nematostella vectensis. Dev. Dyn. 2013, 242, 1320–1331. [Google Scholar] [CrossRef]
- Amiel, A.R.; Johnston, H.T.; Nedoncelle, K.; Warner, J.F.; Ferreira, S.; Röttinger, E. Characterization of Morphological and Cellular Events Underlying Oral Regeneration in the Sea Anemone, Nematostella vectensis. Int. J. Mol. Sci. 2015, 16, 28449–28471. [Google Scholar] [CrossRef]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The Genome of Aiptasia, a Sea Anemone Model for Coral Symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef] [Green Version]
- Surm, J.M.; Stewart, Z.K.; Papanicolaou, A.; Pavasovic, A.; Prentis, P.J. The Draft Genome of Actinia Tenebrosa Reveals Insights into Toxin Evolution. Ecol. Evol. 2019, 9, 11314–11328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, S.I.; Urbarova, I.; Johansen, S.D. Expression of Homing Endonuclease Gene and Insertion-like Element in Sea Anemone Mitochondrial Genomes: Lesson Learned from Anemonia viridis. Gene 2018, 652, 78–86. [Google Scholar] [CrossRef]
- Wilding, C.S.; Fletcher, N.; Smith, E.K.; Prentis, P.; Weedall, G.D.; Stewart, Z. The Genome of the Sea Anemone Actinia equina (L.): Meiotic Toolkit Genes and the Question of Sexual Reproduction. Mar. Genom. 2020, 53, 100753. [Google Scholar] [CrossRef]
- Fredman, D.; Schwaiger, M.; Rentzsch, F.; Technau, U. Nematostella vectensis Transcriptome and Gene Models v2.0. Available online: https://doi.org/10.6084/m9.figshare.807696.v2 (accessed on 15 December 2020).
- Godwin, J.W.; Brockes, J.P. Regeneration, Tissue Injury and the Immune Response. J. Anat. 2006, 209, 423–432. [Google Scholar] [CrossRef]
- Eming, S.A.; Hammerschmidt, M.; Krieg, T.; Roers, A. Interrelation of Immunity and Tissue Repair or Regeneration. Semin. Cell Dev. Biol. 2009, 20, 517–527. [Google Scholar] [CrossRef]
- Abnave, P.; Ghigo, E. Role of the Immune System in Regeneration and Its Dynamic Interplay with Adult Stem Cells. Semin. Cell Dev. Biol. 2019, 87, 160–168. [Google Scholar] [CrossRef]
- Fukazawa, T.; Naora, Y.; Kunieda, T.; Kubo, T. Suppression of the Immune Response Potentiates Tadpole Tail Regeneration during the Refractory Period. Development 2009, 136, 2323–2327. [Google Scholar] [CrossRef] [Green Version]
- Peiris, T.H.; Hoyer, K.K.; Oviedo, N.J. Innate Immune System and Tissue Regeneration in Planarians: An Area Ripe for Exploration. Semin. Immunol. 2014, 26, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Altincicek, B.; Vilcinskas, A. Comparative Analysis of Septic Injury-Inducible Genes in Phylogenetically Distant Model Organisms of Regeneration and Stem Cell Research, the Planarian Schmidtea Mediterranea and the Cnidarian Hydra vulgaris. Front. Zool. 2008, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Godwin, J.W.; Pinto, A.R.; Rosenthal, N.A. Macrophages Are Required for Adult Salamander Limb Regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 9415–9420. [Google Scholar] [CrossRef] [Green Version]
- Kyritsis, N.; Kizil, C.; Zocher, S.; Kroehne, V.; Kaslin, J.; Freudenreich, D.; Iltzsche, A.; Brand, M. Acute Inflammation Initiates the Regenerative Response in the Adult Zebrafish Brain. Science 2012, 338, 1353–1356. [Google Scholar] [CrossRef]
- Brennan, J.J.; Gilmore, T.D. Evolutionary Origins of Toll-like Receptor Signaling. Mol. Biol. Evol. 2018, 35, 1576–1587. [Google Scholar] [CrossRef] [Green Version]
- Halanych, K.M.; Kocot, K.M. Repurposed Transcriptomic Data Facilitate Discovery of Innate Immunity Toll-like Receptor (TLR) Genes across Lophotrochozoa. Biol. Bull. 2014, 227, 201–209. [Google Scholar] [CrossRef]
- Miller, D.J.; Hemmrich, G.; Ball, E.E.; Hayward, D.C.; Khalturin, K.; Funayama, N.; Agata, K.; Bosch, T.C. The Innate Immune Repertoire in Cnidaria-Ancestral Complexity and Stochastic Gene Loss. Genome Biol. 2007, 8, R59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Jin, P.; Qin, S.; Chen, L.; Ma, F. The Evolution and Origin of Animal Toll-like Receptor Signaling Pathway Revealed by Network-Level Molecular Evolutionary Analyses. PLoS ONE 2012, 7, e51657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, L.M.; Fuess, L.E.; Brennan, J.J.; Mansfield, K.M.; Salas-Rodriguez, E.; Welsh, J.; Awtry, J.; Banic, S.; Chacko, C.; Chezian, A.; et al. A Conserved Toll-like Receptor-to-NF-ΚB Signaling Pathway in the Endangered Coral Orbicella Faveolata. Dev. Comp. Immunol. 2018, 79, 128–136. [Google Scholar] [CrossRef]
- Zárate-Potes, A.; Ocampo, I.D.; Cadavid, L.F. The Putative Immune Recognition Repertoire of the Model Cnidarian Hydractinia symbiolongicarpus Is Large and Diverse. Gene 2019, 684, 104–117. [Google Scholar] [CrossRef]
- Milde, S.; Hemmrich, G.; Anton-Erxleben, F.; Khalturin, K.; Wittlieb, J.; Bosch, T.C. Characterization of Taxonomically Restricted Genes in a Phylum-Restricted Cell Type. Genome Biol. 2009, 10, R8. [Google Scholar] [CrossRef] [Green Version]
- Wolenski, F.S.; Bradham, C.A.; Finnerty, J.R.; Gilmore, T.D. NF-ΚB Is Required for Cnidocyte Development in the Sea Anemone Nematostella vectensis. Dev. Biol. 2013, 373, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, J.J.; Messerschmidt, J.L.; Williams, L.M.; Matthews, B.J.; Reynoso, M.; Gilmore, T.D. Sea Anemone Model Has a Single Toll-like Receptor That Can Function in Pathogen Detection, NF-ΚB Signal Transduction, and Development. Proc. Natl. Acad. Sci. USA 2017, 114, E10122–E10131. [Google Scholar] [CrossRef] [Green Version]
- Babonis, L.S.; Ryan, J.F.; Enjolras, C.; Martindale, M.Q. Genomic Analysis of the Tryptome Reveals Molecular Mechanisms of Gland Cell Evolution. EvoDevo 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverri, K.; Zayas, R.M. Regeneration: From Cells to Tissues to Organisms. Dev. Biol. 2018, 433, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, S.M.; Gotting, K.; Ross, E.; Alvarado, A.S.; Swalla, B.J. Head Regeneration in Hemichordates Is Not a Strict Recapitulation of Development. Dev. Dyn. 2016, 245, 1159–1175. [Google Scholar] [CrossRef] [Green Version]
- Bellis, E.S.; Howe, D.K.; Denver, D.R. Genome-Wide Polymorphism and Signatures of Selection in the Symbiotic Sea Anemone Aiptasia. BMC Genom. 2016, 17, 160. [Google Scholar] [CrossRef] [Green Version]
- Bucher, M.; Wolfowicz, I.; Voss, P.A.; Hambleton, E.A.; Guse, A. Development and Symbiosis Establishment in the Cnidarian Endosymbiosis Model Aiptasia sp. Sci. Rep. 2016, 6, 19867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, C.A.; Ameismeier, M.F.; Peng, L.; Weis, V.M.; Grossman, A.R.; Davy, S.K. Symbiosis Induces Widespread Changes in the Proteome of the Model Cnidarian aiptasia. Cell Microbiol. 2016, 18, 1009–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, V.A.S.; Bucher, M.; Hambleton, E.A.; Guse, A. Microinjection to Deliver Protein, MRNA, and DNA into Zygotes of the Cnidarian Endosymbiosis Model Aiptasia sp. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Burg, C.A.; Prentis, P.J. The Tentacular Spectacular: Evolution of Regeneration in Sea Anemones. Genes 2021, 12, 1072. https://doi.org/10.3390/genes12071072
van der Burg CA, Prentis PJ. The Tentacular Spectacular: Evolution of Regeneration in Sea Anemones. Genes. 2021; 12(7):1072. https://doi.org/10.3390/genes12071072
Chicago/Turabian Stylevan der Burg, Chloé A., and Peter J. Prentis. 2021. "The Tentacular Spectacular: Evolution of Regeneration in Sea Anemones" Genes 12, no. 7: 1072. https://doi.org/10.3390/genes12071072
APA Stylevan der Burg, C. A., & Prentis, P. J. (2021). The Tentacular Spectacular: Evolution of Regeneration in Sea Anemones. Genes, 12(7), 1072. https://doi.org/10.3390/genes12071072