
QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica Napus L.)

 ,
,  , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
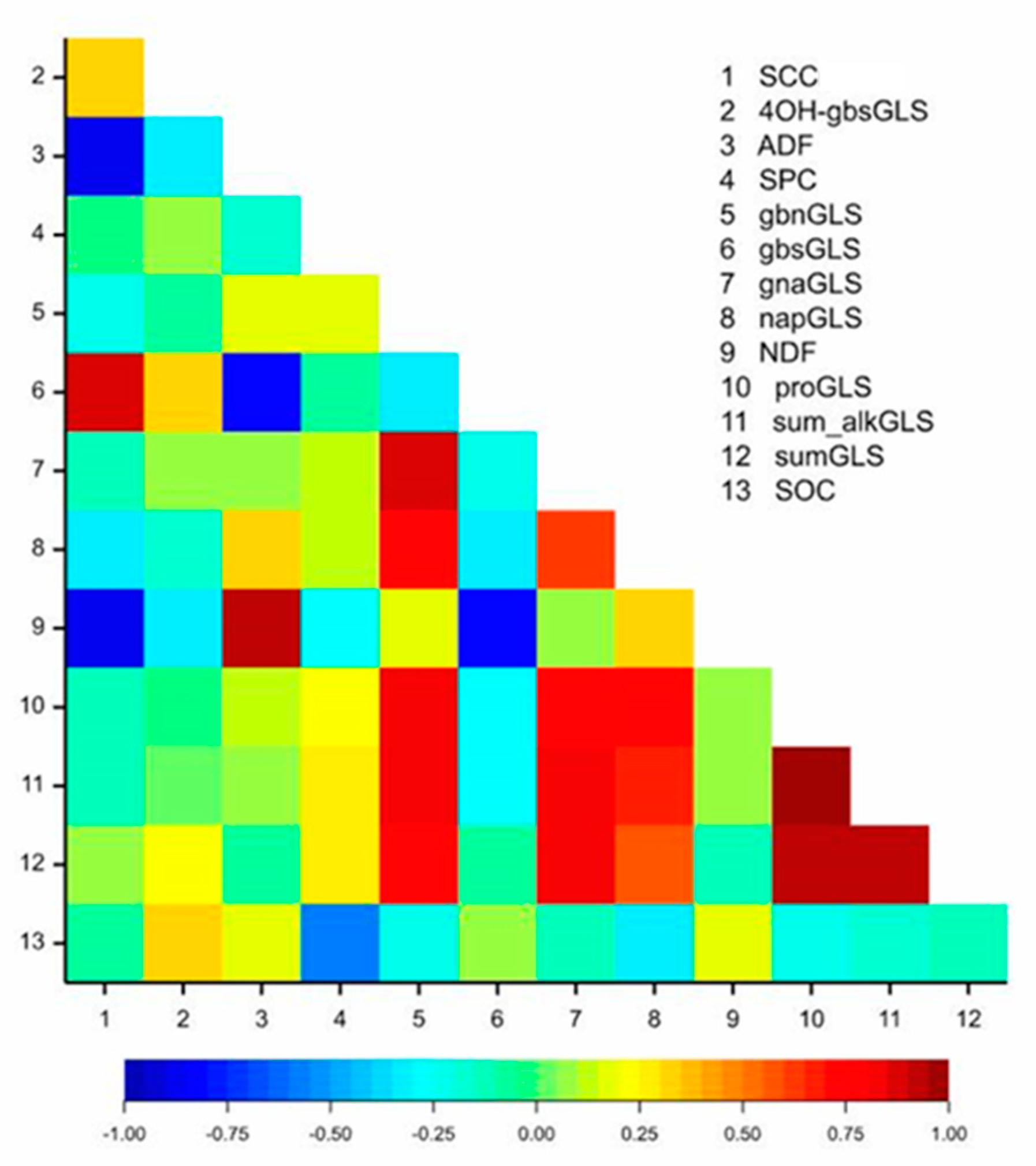
2.1. Phenotypic Analysis
2.2. Genome-Wide Genotyping of the Mapping Population
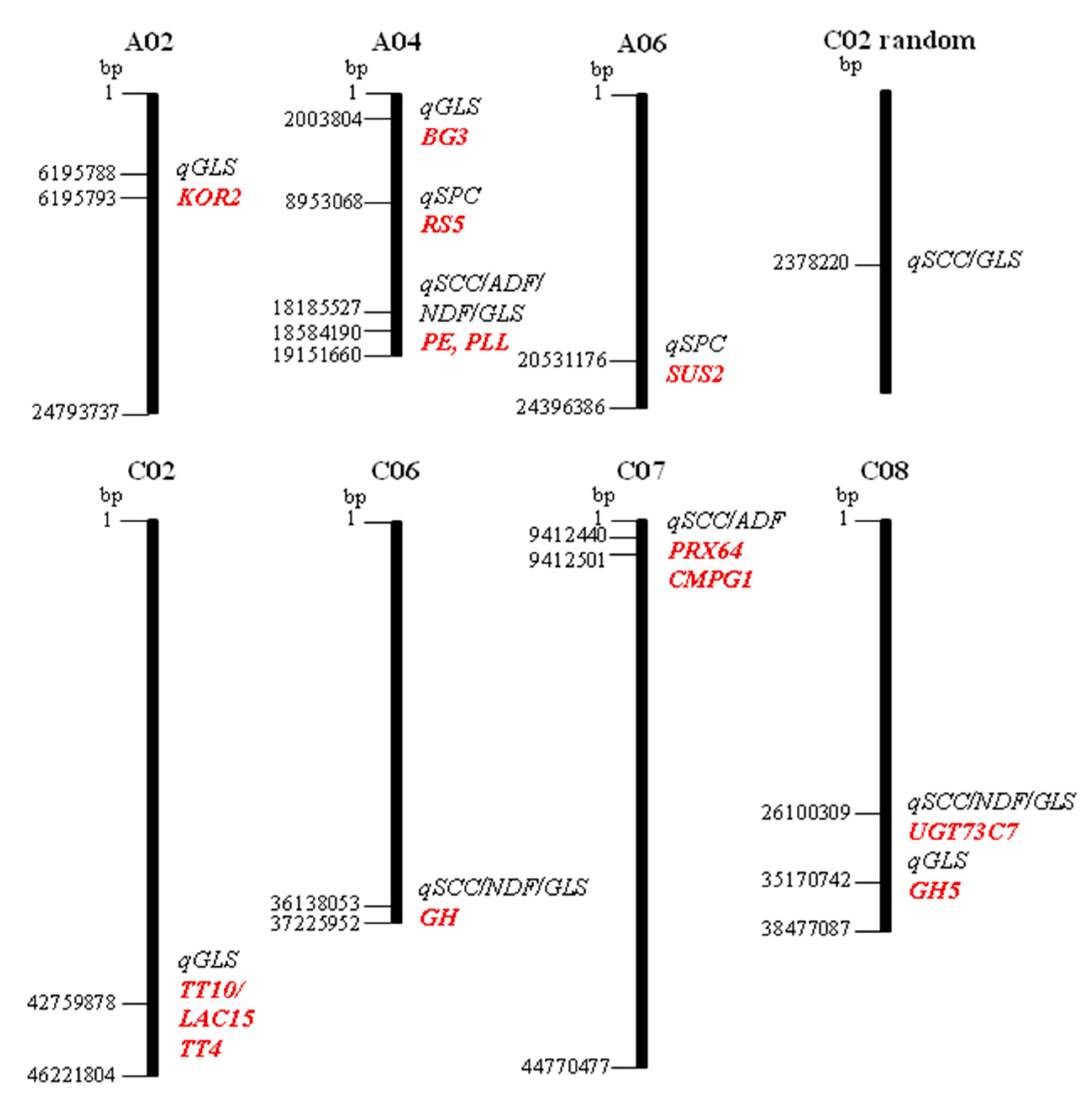
2.3. QTL Analysis for Seed Coat Colour, Seed Fibre, Glucosinolates, and Protein Content
2.4. Candidate Gene Prediction Underling QTL for Seed Coat Colour, Seed Fibre, Glucosinolates, and Protein Content
3. Discussion
4. Materials and Methods
4.1. Plant Material and Field Trials
4.2. Trait Analysis
4.3. Genotyping and SNP Marker Analysis
4.4. QTL Mapping
4.5. Identification of Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wittkop, B.; Snowdon, R.J.; Friedt, W. Status and perspectives of breeding for enhanced yield and quality of oilseed crops for Europe. Euphytica 2009, 170, 131–140. [Google Scholar] [CrossRef]
- Liu, L.Z.; Qu, C.M.; Wittkop, B.; Yi, B.; Xiao, Y.J.; He, Y.; Snowdon, R.J.; Li, J.N. A High-Density SNP Map for Accurate Mapping of Seed Fibre QTL in Brassica napus L. PLoS ONE 2013, 8, e83052. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jian, H.J.; Wei, L.J.; Qu, C.M.; Xu, X.F.; Lu, K.; Qian, W.; Li, J.N.; Li, M.T.; Liu, L.Z. Genome-Wide Analysis of Seed Acid Detergent Lignin (ADL) and Hull Content in Rapeseed (Brassica napus L.). PLoS ONE 2015, 10, e0145045. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xian, X.H.; Xu, X.F.; Qu, C.M.; Lu, K.; Li, J.N.; Liu, L.Z. Genome-Wide Association Mapping of Seed Coat Color in Brassica napus. J. Agric. Food Chem. 2017, 65, 5229–5237. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Wittkop, B.; Liu, L.Z.; Obermeier, C.; Friedt, W.; Snowdon, R.J. Dissection of a major QTL for seed colour and fibre content in Brassica napus reveals colocalization with candidate genes for phenylpropanoid biosynthesis and flavonoid deposi-tion. Plant Breed. 2013, 132, 382–389. [Google Scholar] [CrossRef]
- Badani, A.G.; Snowdon, R.J.; Wittkop, B.; Lipsa, F.D.; Baetzel, R.; Horn, R.; De Haro, A.; Font, R.; Luhs, W.; Friedt, W. Colocalization of a partially dominant gene for yellow seed colour with a major QTL influencing acid detergent fibre (ADF) content in different crosses of oilseed rape (Brassica napus). Genome 2006, 49, 1499–1509. [Google Scholar] [CrossRef]
- Snowdon, R.J.; Wittkop, B.; Rezaidad, A.; Hasan, M.; Lipsa, F.D.; Stein, A.; Friedt, W. Regional association analysis delineates a sequenced chromosome region influencing antinutritive seed meal compounds in oilseed rape. Genome 2010, 53, 917–928. [Google Scholar] [CrossRef]
- Liu, L.Z.; Stein, A.; Wittkop, B.; Sarvari, P.; Li, J.N.; Yan, X.Y.; Dreyer, F.; Frauen, M.; Friedt, W.; Snowdon, R.J. A knockout mutation in the lignin biosynthesis gene CCR1 explains a major QTL for acid detergent lignin content in Brassica napus seeds. Theor. Appl. Genet. 2012, 124, 1573–1586. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.Y.; Liu, L.-Z.; Chai, Y.-R.; Chen, L.; Yang, T.; Jin, M.-Y.; Ma, A.-F.; Yan, X.-Y.; Zhang, Z.-S.; Li, J.-N. Localization of QTLs for seed color using recombinant inbred lines of Brassica napus in different environments. Genome 2007, 50, 840–854. [Google Scholar] [CrossRef]
- Behnke, N.; Suprianto, E.; Möllers, C. A major QTL on chromosome C05 significantly reduces acid detergent lignin (ADL) content and increases seed oil and protein content in oilseed rape (Brassica napus L.). Theor. Appl. Genet. 2018, 131, 2477–2492. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.Y.; Chao, H.B.; Chen, L.; Wang, H.; Zhao, W.; Li, B.; Zhang, L.; Li, H.; Wang, B.; Li, M. Stable and novel QTL identification and new insights into the genetic networks affecting seed fiber traits in Brassica napus. Theor. Appl. Genet. 2019, 132, 1761–1775. [Google Scholar] [CrossRef]
- Marles, M.A.S.; Gruber, M.Y. Histochemical characterisation of unextractable seed coat pigments and quantification of extractable lignin in the Brassicaceae. J. Sci. Food Agric. 2004, 84, 251–262. [Google Scholar] [CrossRef]
- Stein, A.; Coriton, O.; Rousseau-Gueutin, M.; Samans, B.; Schiessl, S.V.; Obermeier, C.; Parkin, I.A.P.; Chevre, A.-M.; Snowdon, R.J. Mapping of homoeologous chromosome exchanges influencing quantitative trait variation in Brassica napus. Plant Biotechnol. J. 2017, 15, 1478–1489. [Google Scholar] [CrossRef] [Green Version]
- Hurgobin, B.; Golicz, A.A.; Bayer, P.E.; Chan, C.-K.; Tirnaz, S.; Dolatabadian, A.; Schiessl, S.V.; Samans, B.; Montenegro, J.D.; Parkin, I.A.P.; et al. Homoeologous exchange is a major cause of gene presence/absence variation in the amphidiploid Brassica napus. Plant Biotechnol. J. 2018, 16, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- Song, J.-M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef]
- Qu, C.M.; Fu, F.Y.; Lu, K.; Zhang, K.; Wang, R.; Xu, X.F.; Wang, M.; Lu, J.X.; Wan, H.F.; Tang, Z.L.; et al. Differential accumulation of phenolic compounds and expression of related genes in black- and yellow-seeded Brassica napus. J. Exp. Bot. 2013, 64, 2885–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libertini, E.; Li, Y.; McQueen-Mason, S.J. Phylogenetic analysis of the plant endo-beta-1,4-glucanase gene family. J. Mol. Evol. 2004, 58, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Minic, Z.; Jouanin, L. Plant glycoside hydrolases involved in cell wall polysaccharide degradation. Plant Physiol. Biochem. 2006, 44, 435–449. [Google Scholar] [CrossRef]
- Gangl, R.; Tenhaken, R. Raffinose Family Oligosaccharides Act As Galactose Stores in Seeds and Are Required for Rapid Germination of Arabidopsisin the Dark. Front. Plant Sci. 2016, 7, 1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.-C.; Bulgakov, V.P.; Jinn, T.-L. Pectin Methylesterases: Cell Wall Remodeling Proteins Are Required for Plant Response to Heat Stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]
- Vogel, J.P.; Raab, T.K.; Schiff, C.; Somerville, S.C. PMR6, a Pectate Lyase–Like Gene Required for Powdery Mildew Susceptibility in Arabidopsis. Plant Cell 2002, 14, 2095–2106. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Schumaker, K.S.; Guo, Y. Sumoylation of transcription factor MYB30 by the small ubiquitin-like modifier E3 ligase SIZ1 mediates abscisic acid response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2012, 109, 12822–12827. [Google Scholar] [CrossRef] [Green Version]
- Angeles-Nunez, J.G.; Tiessen, A. Regulation of AtSUS2 and AtSUS3 by glucose and the transcription factor LEC2 in different tissues and at different stages of Arabidopsis seed development. Plant Mol. Biol. 2012, 78, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.-M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 Encodes a Laccase-Like Enzyme Involved in Oxidative Polymerization of Flavonoids in Arabidopsis Seed Coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, T.; Takahashi, S.; Waki, T. Formation of Flavonoid Metabolons: Functional Significance of Protein-Protein Inter-actions and Impact on Flavonoid Chemodiversity. Front. Plant Sci. 2019, 10, 821. [Google Scholar] [CrossRef]
- Henrissat, B.; Callebaut, I.; Fabrega, S.; Lehn, P.; Mornon, J.P.; Davies, G. Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proc. Natl. Acad. Sci. USA 1996, 93, 5674. [Google Scholar]
- Tobimatsu, Y.; Schuetz, M. Lignin polymerization: How do plants manage the chemistry so well? Curr. Opin. Biotechnol. 2019, 56, 75–81. [Google Scholar] [CrossRef]
- Berrocal-Lobo, M.; Stone, S.; Yang, X.; Antico, J.; Callis, J.; Ramonell, K.M.; Somerville, S. ATL9, a RING Zinc Finger Protein with E3 Ubiquitin Ligase Activity Implicated in Chitin- and NADPH Oxidase-Mediated Defense Responses. PLoS ONE 2010, 5, e14426. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.-S.; Huang, X.-X.; Li, Q.; Cao, Y.P.; Bao, Y.; Meng, X.-F.; Li, Y.-J.; Fu, C.X.; Hou, B.-K. UDP-glycosyltransferase 72B1 catalyzes the glucose conjugation of monolignols and is essential for the normal cell wall lignification in Arabidopsis thaliana. Plant J. 2016, 88, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.; Batley, J.; Snowdon, R.J. Accessing complex crop genomes with next-generation sequencing. Theor. Appl. Genet. 2013, 126, 1–11. [Google Scholar] [CrossRef]
- Gacek, K.; Bayer, P.E.; Bartkowiak-Broda, I.; Szała, L.; Bocianowski, J.; Edwards, D.; Batley, J. Genome-Wide Association Study of Genetic Control of Seed Fatty Acid Biosynthesis in Brassica napus. Front. Plant Sci. 2017, 7, 2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolko, J.; Dobrzycka, A.; Bocianowski, J.; Szala, L.; Cegielska-Taras, T.; Bartkowiak-Broda, I.; Gacek, K. Genetic variation of traits affecting meal quality in black × yellow seeded doubled haploid population of winter oilseed rape. Agron. Res. 2020, 18, 2259–2270. [Google Scholar]
- Browning, S.R.; Browning, B.L. High-Resolution Detection of Identity by Descent in Unrelated Individuals. Am. J. Hum. Genet. 2010, 86, 526–539. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.M. Factors affecting the nutritional-value of canola-meal—A Review. Can. J. Anim. Sci. 1993, 73, 679–697. [Google Scholar] [CrossRef]
- Farrell, D.J.; Perez-Maldonado, R.A. Tannins in feedstuffs used in the diets of pigs and poultry in Australia. In Proceedings of the International Workshop on Tannins in Livestock and Human Nutrition, Adelaide, Australia, 31 May–2 June 1999; pp. 24–29. [Google Scholar]
- Simbaya, J.; Slominski, B.A.; Rakow, G.; Campbell, L.D.; Downey, R.K.; Bell, J.M. Quality characteristics of yellow-seeded brassica seed meals—protein, carbohydrates, and dietary fiber components. J. Agric. Food Chem. 1995, 43, 2062–2066. [Google Scholar] [CrossRef]
- Slominski, B.A.; Simbaya, J.; Campbell, L.D.; Rakow, G.; Guenter, W. Nutritive value for broilers of meals derived from newly developed varieties of yellow-seeded canola. Anim. Feed. Sci. Technol. 1999, 78, 249–262. [Google Scholar] [CrossRef]
- Hannoufa, A.; Pillai, B.V.S.; Chellamma, S. Genetic enhancement of Brassica napus seed quality. Transgenic Res. 2014, 23, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Stombaugh, S.K.; Jung, H.G.; Orf, J.H.; Somers, D.A. Genotypic and Environmental Variation in Soybean Seed Cell Wall Polysaccharides. Crop Sci. 2000, 40, 408–412. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Chen, W.; Yi, B.; Wen, J.; Shen, J.; Ma, C.; Chen, B.; Tu, J.; Fu, T. Identification of two major QTL for yellow seed color in two crosses of resynthesized Brassica napus line No. 2127-17. Mol. Breed. 2011, 28, 335–342. [Google Scholar] [CrossRef]
- Yu, C.-Y. Molecular mechanism of manipulating seed coat coloration in oilseed Brassica species. J. Appl. Genet. 2013, 54, 135–145. [Google Scholar] [CrossRef]
- del Campillo, E. Multiple Endo-1,4-beta-D-glucanase (cellulase) genes in Arabidopsis. Curr. Top. Dev. Biol. 1999, 46, 39–56, 56A, 57–61. [Google Scholar] [PubMed]
- Francoz, E.; Ranocha, P.; Nguyen-Kim, H.; Jamet, E.; Burlat, V.; Dunand, C. Roles of cell wall peroxidases in plant develop-ment. Phytochemistry 2015, 112, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.-R.; Lei, B.; Huang, H.-L.; Li, J.-N.; In, J.-M.; Tang, Z.-L.; Wang, R.; Chen, L. TRANSPARENTTESTA12 genes from Brassica napus and parental species: Cloning, evolution, and differential involvement in yellow seed trait. Mol. Genet. Genom. 2009, 281, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Krzymański, J.; Bartkowiak-Broda, I.; Krótka, K. Characteristic of yellow-seeded lines of winter oilseed rape. In Proceedings of the Rapeseed Congress, Copenhagen, Denmark, 6–10 July 2003. [Google Scholar]
- Michalski, K. Seed color assessment in rapeseed seeds using Color and Near Infrared Reflectance Spectrometers. Oilseed Crop 2009, XXI, 801–806. [Google Scholar]
- Li, R.; Yu, C.; Li, Y.; Lam, T.-W.; Yiu, S.-M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Lorenc, M.T.; Hayashi, S.; Stiller, J.; Lee, H.; Manoli, S.; Ruperao, P.; Visendi, P.; Berkman, P.J.; Lai, K.; Batley, J.; et al. Discovery of Single Nucleotide Polymorphisms in Complex Genomes Using SGSautoSNP. Biology 2012, 1, 370–382. [Google Scholar] [CrossRef] [Green Version]
- Bayer, P.E.; Ruperao, P.; Mason, A.S.; Stiller, J.; Chan, C.-K.K.; Hayashi, S.; Long, Y.; Meng, J.; Sutton, T.; Visendi, P.; et al. High-resolution skim genotyping by sequencing reveals the distribution of crossovers and gene con-versions in Cicer arietinum and Brassica napus. Theor. Appl. Genet. 2015, 128, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Trait | Z114 Mean | M305 Mean | DH Mean | DH Range | E | G | G×E | |
|---|---|---|---|---|---|---|---|---|
| d.f. | 1 | 79 | 79 | |||||
| SOC | 41.6 ± 3.1 | 43.4 ± 2.06 | 41.5 ± 3 | 35.45–49.46 | *** | *** | *** | |
| SPC | 24.21 ± 1.95 | 24.3 ± 1.3 | 25.05 ± 1.4 | 19.33–29.36 | *** | *** | *** | |
| SCC | 4.70 ± 0.6 | 0.47 ± 0.65 | 1.54 ± 0.75 | 0.03–5.1 | *** | *** | *** | |
| NDF | 17.13 ± 0.85 | 23.05 ± 0.9 | 21.36 ± 1.9 | 16.47–26.86 | ns | *** | *** | |
| ADF | 10.05 ± 0.7 | 16.9 ± 0.8 | 14.88 ± 2.4 | 6.79–22.15 | *** | *** | *** | |
| 4OH-gbsGLS | 4.96 ± 0.45 | 4.52 ± 0.6 | 4.98 ± 0.7 | 2.1–6.82 | *** | *** | *** | |
| gbsGLS | 0.36 ± 0.03 | 0.21 ± 0.01 | 0.23 ± 0.04 | 0.12–0.4 | *** | *** | *** | |
| gbnGLS | 0.33 ± 0.15 | 0.47 ± 0.1 | 0.57 ± 0.2 | 0.03–2.15 | *** | *** | *** | |
| gnaGLS | 1.23 ± 0.5 | 3.88 ± 0.6 | 2.59 ± 0.9 | 0.54–9.04 | *** | *** | *** | |
| napGLS | 0.08 ± 0.01 | 0.01 ± 0.01 | 0.10 ± 0.02 | 0.04–0.25 | ** | *** | ** | |
| proGLS | 1.91 ± 1.19 | 2.73 ± 1.55 | 4.18 ± 2.2 | 0.1–20.9 | *** | *** | *** | |
| sum_alkGLS | 3.75 ± 2.65 | 5.19 ± 2.27 | 7.49 ± 3.46 | 0.12–30.47 | *** | *** | *** | |
| sumGLS | 9.22 ± 2.05 | 8.96 ± 2.1 | 12.13 ± 3.5 | 4.47–31.23 | *** | *** | *** |
| QTL | Trait | Chr | Position | LOD Score | R2 (%) PVE |
|---|---|---|---|---|---|
| qGLS-A02 | sumGLS, gnaGLS, napGLS, gbnGLS proGLS, sum_alkGLS | A02 | 6195793, 6195788 | 4.27, 6.61 | 21.79, 31.65 |
| qGLS-A04 | proGLS, napGLS | A04 | 2003804 | 6.55 | 31.41 |
| qSPC-A04 | SPC | A04 | 8953068 | 5.52 | 27.22 |
| qSCC/ADF/NDF/GLS-A04 | SCC, ADF, NDF, gbsGLS | A04 | 18584190, 18185527 | 4.32, 4.21 | 22.02, 21.52 |
| qSPC-A06 | SPC | A06 | 20531176 | 4.49 | 22.78 |
| qGLS-C02 | gbnGLS, proGLS | C02 | 42759878 | 6.85 | 32.59 |
| qSCC/GLS-C02r | SCC, gbsGLS | C02-random | 2378220 | 4.62 | 23.35 |
| qSCC/NDF/GLS-C06 | SCC, NDF, gbsGLS | C06 | 36138053 | 4.82 | 24.23 |
| qSCC/ADF-C07 | SCC, ADF | C07 | 9412440, 9412501 | 4.78, 5.59 | 24.05, 27.51 |
| qSCC/NDF/GLS-C08 | SCC, NDF, gbsGLS | C08 | 26100309 | 4.42 | 22.46 |
| qGLS-C08 | gbnGLS, sumGLS, sum_alkGLS | C08 | 35170742 | 4.77 | 24.01 |
| QTL | QTL Size | No. Genes | B. napus | A.thaliana | Functional Annotation | References |
|---|---|---|---|---|---|---|
| qGLS-A02 | 2.5 kbp | 20 | BnaA02g12160D | AT1G65610.1 | Korrigan 2 (KOR2) | [18] |
| qGLS-A04 | 4,8 kbp | 5 | BnaA04g03060D | AT3G57240.1 | β-1,3-glucanase 3 (BG3), glycoside hydrolase, cell wall degradation, | [19] |
| qSPC-A04 | 70.3 kbp | 20 | BnaA04g10260D | AT5G40390 | Raffinose synthase 5 (RS5), Seed imbibition 1-like (SIP1) | [20] |
| qSCC/ADF/NDF/GLS-A04 | 405.8 kbp | 8 | BnaA04g27070D BnaA04g25420D | AT2G47550.1 AT2G43860.1 | Pectin methylesterase, (PE) cell wall modification Pectin lyase-like protein, (PLL), carbohydrate metabolic process | [21,22] |
| qSPC-A06 | 1.9 kbp | 19 | BnaA06g29670D | AT5G49190 | Sucrose synthase 2 (SUS2) seed maturation, starch, sucrose metabolic process, plant cell wall | [23,24] |
| qGLS-C02 | 111 bp | 8 | BnaC02g38340D BnaC02g38710D | AT5G48100.1 AT5G13930.1 | Transparent testa 10, laccase-like 15, (TT10/LAC15) lignin and flavonoids biosynthesis Transparent testa 4, (TT4) chalcone synthase, flavonoid biosynthesis | [25,26] |
| qSCC/GLS-C02r | 1 kbp | 1 | BnaC02g47290D | AT5G46040.1 | Major facilitator superfamily protein | |
| qSCC/NDF/GLS-C06 | 575 bp | 5 | BnaC06g38540D | AT1G78060.1 | Glycosyl hydrolase, (GH) carbohydrate metabolic process, plant-type cell wall | [27] |
| qSCC/ADF-C07 | 240 bp | 2 | BnaC07g05770D BnaC07g05860D | AT5G42180.1 AT5G01830.1 | Peroxidase 64, (PRX64), lignin biosynthesis, plant cell wall RING-type E3 ubiquitin transferase; (CMPG1), response to chitin | [28,29] |
| qSCC/NDF/GLS-C08 | 380 bp | 4 | BnaC08g24250D | AT3G53160.1 | UDP-glucosyl transferase 73C7 (UGT73C7) | [30] |
| qGLS-C08 | 308 bp | 7 | BnaC08g40570D | AT1G13130.1 | Cellulase, glycosyl hydrolase family 5, (GH5), carbohydrate metabolic process | [27] |
| QTL | B. napus | SNP Position | Alleles | Effect of SNP |
|---|---|---|---|---|
| qGLS-A02 | BnaA02g12160D | 6406888 6407629 | G>T A>G | missense Val>Phe missense Arg>Gly |
| qGLS-A04 | BnaA04g03060D | 2014574 2014706 2014718 | C>A A>G T>G | missense Pro>Gln missense Glu>Gly missense Leu>Arg |
| qSPC-A04 | BnaA04g10260D | 9043708 9045761 9046164 9046782 | T>C C>T A>T C>A | missense Tyr>His missense Thr>Ile missense Lys>Asn missense Phe>Leu |
| qSCC/ADF/NDF/GLS-A04 | BnaA04g27070D BnaA04g25420D | 19096076 18283611 | A>C T>C | missense Asn>Thr missense Val>Ala |
| qSPC-A06 | BnaA06g29670D | 20209248 20209490 20209679 20209685 20210405 20210426 20210576 20210746 20212314 20212875 | C>T A>C C>T C>T A>G C>T G>A A>C A>C G>A | missense Ser>Leu missense Ile>Leu missense Arg>Cys missense Pro>Ser missense Arg>Gly missense Pro>Ser missense Glu>Lys missense Arg>Ser stop missense Gly>Asp |
| qGLS-C02 | BnaC02g38340D BnaC02g38710D | 41321282 41652670 41652738 | T>A G>C C>G | missense Phe>Ile missense Trp>Ser missense His>Asp |
| qSCC/GLS-C02r | BnaC02g47290D | 3064584 3064753 3064912 3066441 | T>A T>A G>C T>C | missense Ser>Thr missense Leu>Gln stop missense Phe>Leu |
| qSCC/NDF/GLS-C06 | BnaC06g38540D | 36074371 36074734 36075755 36075812 36076103 36078104 | A>G C>T A>G C>A C>G C>T | missense Gln>Arg missense Thr>Ile missense Thr>Ala missense Leu>Ile missense His>Asp missense Arg>Trp |
| qSCC/ADF-C07 | BnaC07g05770D BnaC07g05860D | 9237994 9238393 9428459 9428556 | A>T G>A C>T G>A | missense Gln>Leu missense Arg>His missense Ala>Val missense Val>Ile |
| qSCC/NDF/GLS-C08 | BnaC08g24250D | 26289421 | G>A | missense Ser>Asn |
| qGLS-C08 | BnaC08g40570D | 35735275 35735343 35735471 35735528 35735805 35735820 35736579 35736883 35737048 35737117 | T>A G>C A>G C>T C>T A>T T>C T>G C>G T>G | missense Ser>Arg missense Glu>Gln missense Asp>Gly missense Ala>Val missense Pro>Ser missense Ile>Leu missense Tyr>His missense Ile>Met missense Ile>Met missense Asn>Lys |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gacek, K.; Bayer, P.E.; Anderson, R.; Severn-Ellis, A.A.; Wolko, J.; Łopatyńska, A.; Matuszczak, M.; Bocianowski, J.; Edwards, D.; Batley, J. QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica Napus L.). Genes 2021, 12, 1235. https://doi.org/10.3390/genes12081235
Gacek K, Bayer PE, Anderson R, Severn-Ellis AA, Wolko J, Łopatyńska A, Matuszczak M, Bocianowski J, Edwards D, Batley J. QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica Napus L.). Genes. 2021; 12(8):1235. https://doi.org/10.3390/genes12081235
Chicago/Turabian StyleGacek, Katarzyna, Philipp E. Bayer, Robyn Anderson, Anita A. Severn-Ellis, Joanna Wolko, Agnieszka Łopatyńska, Marcin Matuszczak, Jan Bocianowski, David Edwards, and Jacqueline Batley. 2021. "QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica Napus L.)" Genes 12, no. 8: 1235. https://doi.org/10.3390/genes12081235
APA StyleGacek, K., Bayer, P. E., Anderson, R., Severn-Ellis, A. A., Wolko, J., Łopatyńska, A., Matuszczak, M., Bocianowski, J., Edwards, D., & Batley, J. (2021). QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica Napus L.). Genes, 12(8), 1235. https://doi.org/10.3390/genes12081235