
Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Immunofluorescence
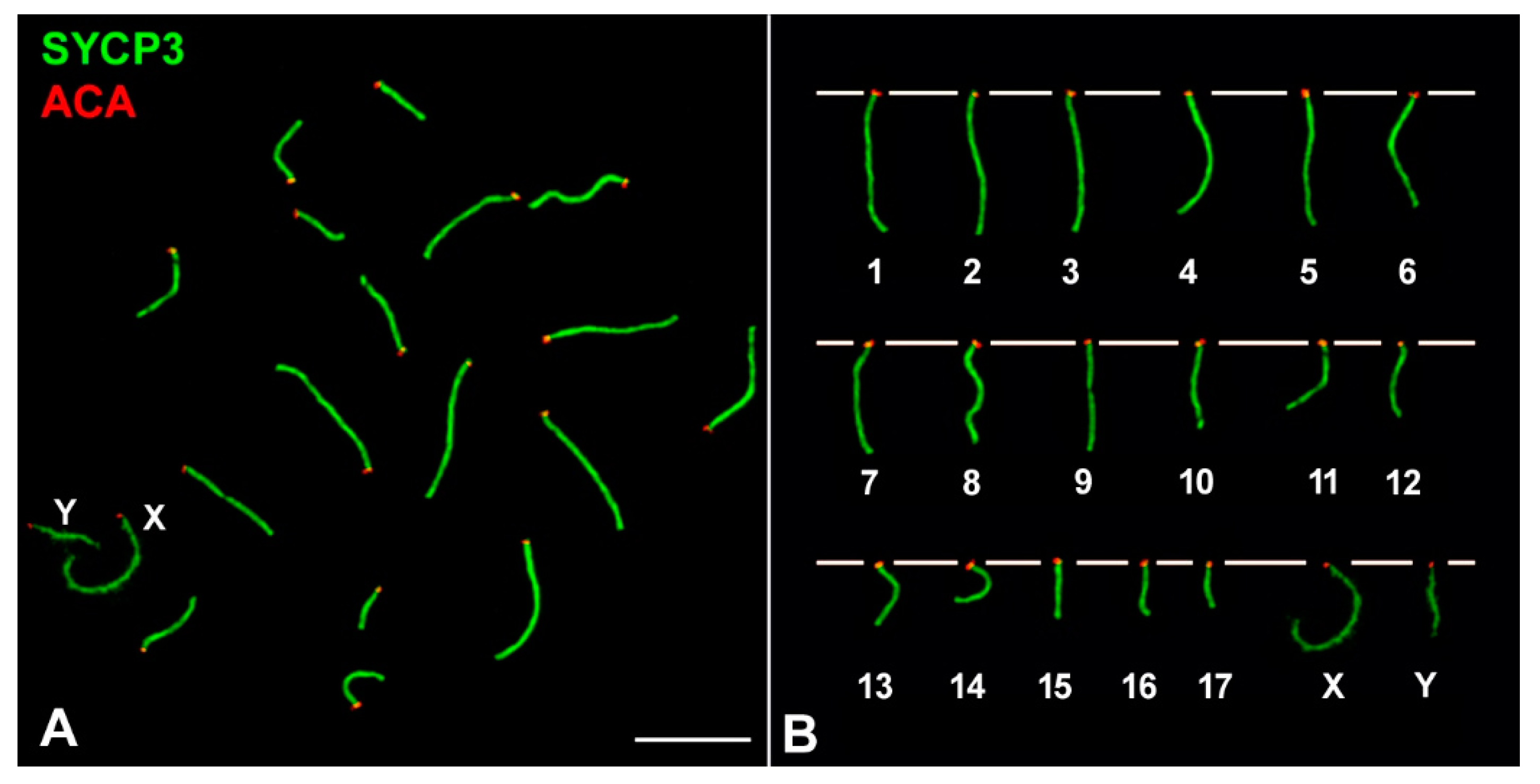
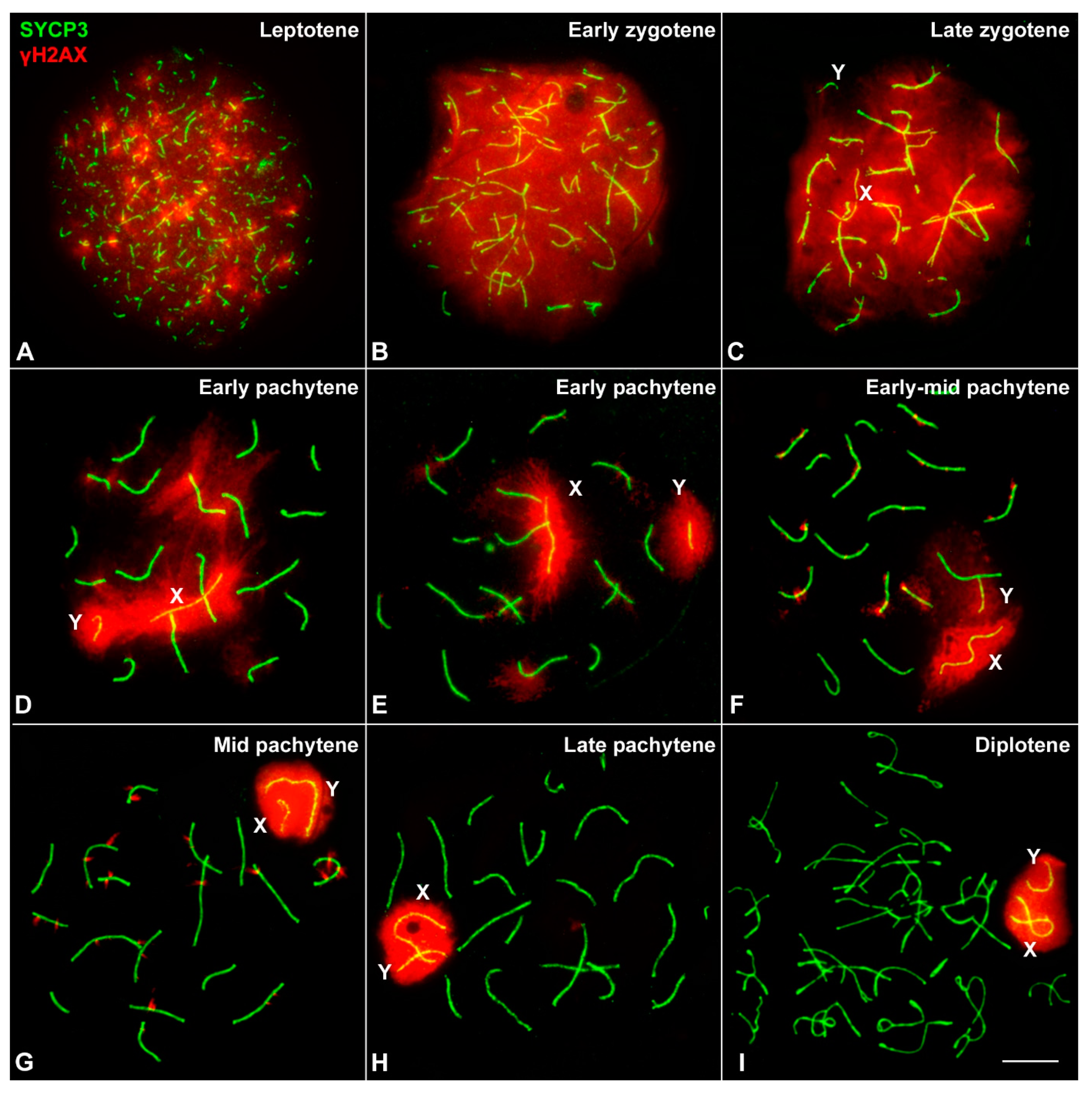
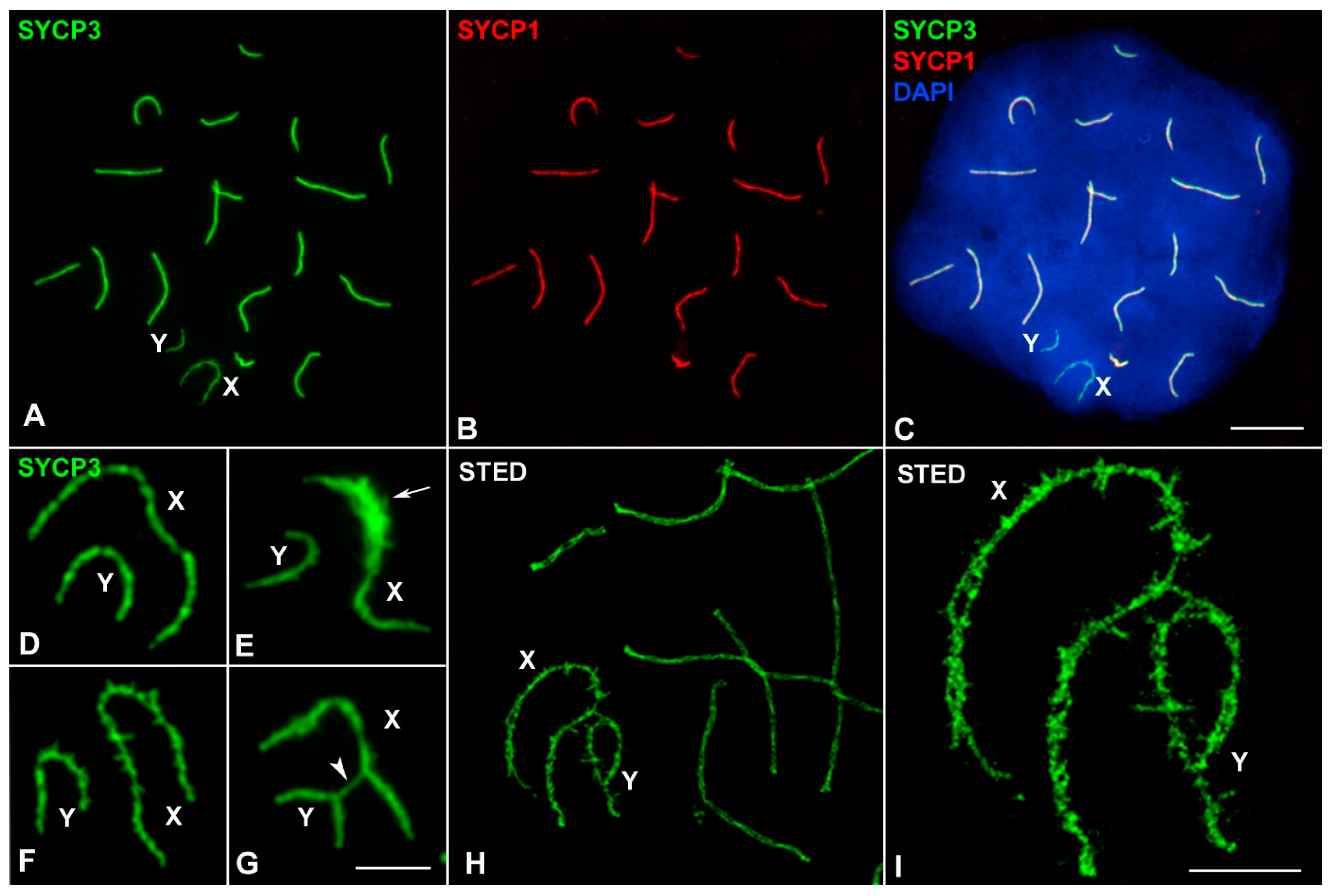
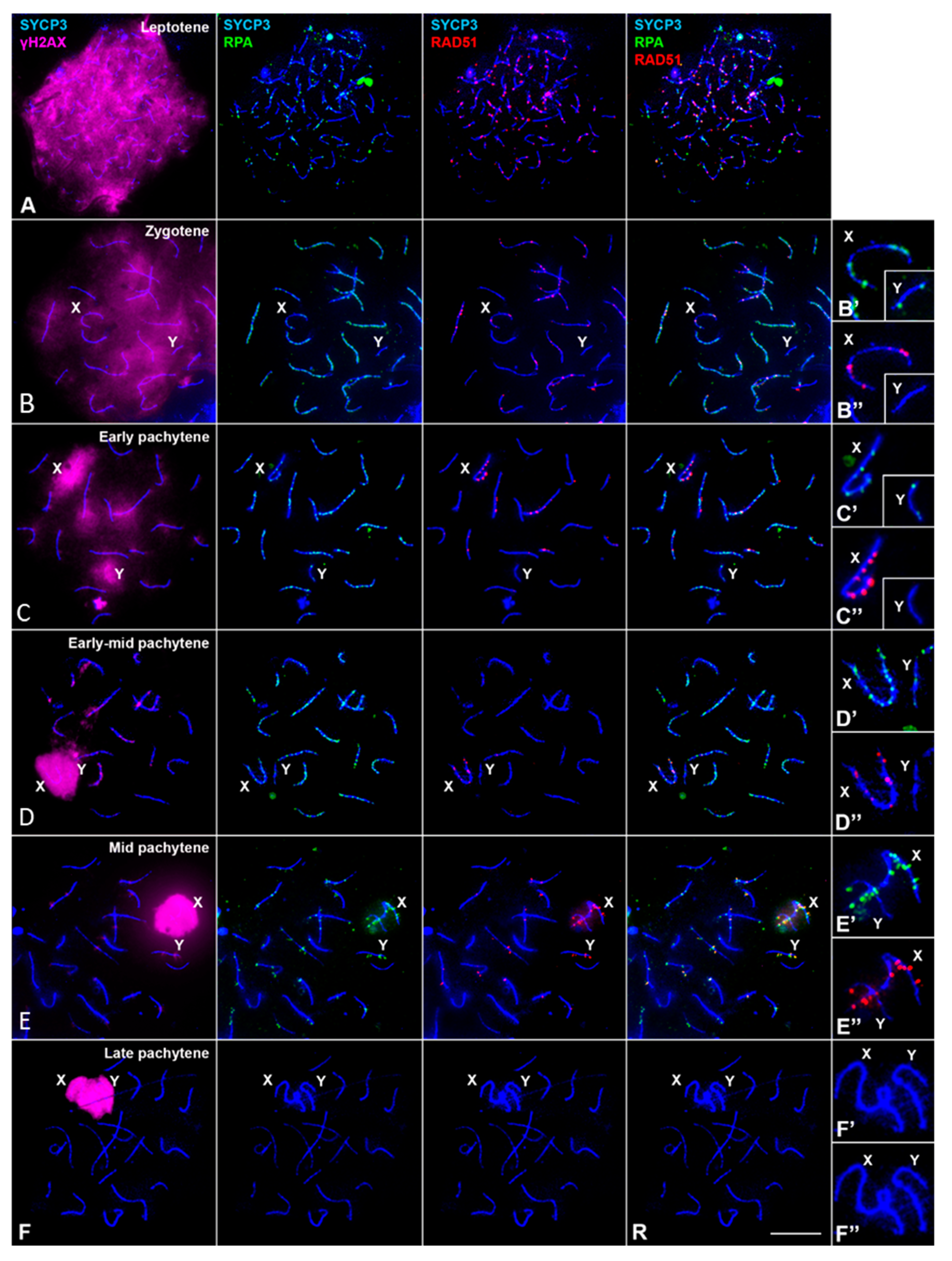
3. Results
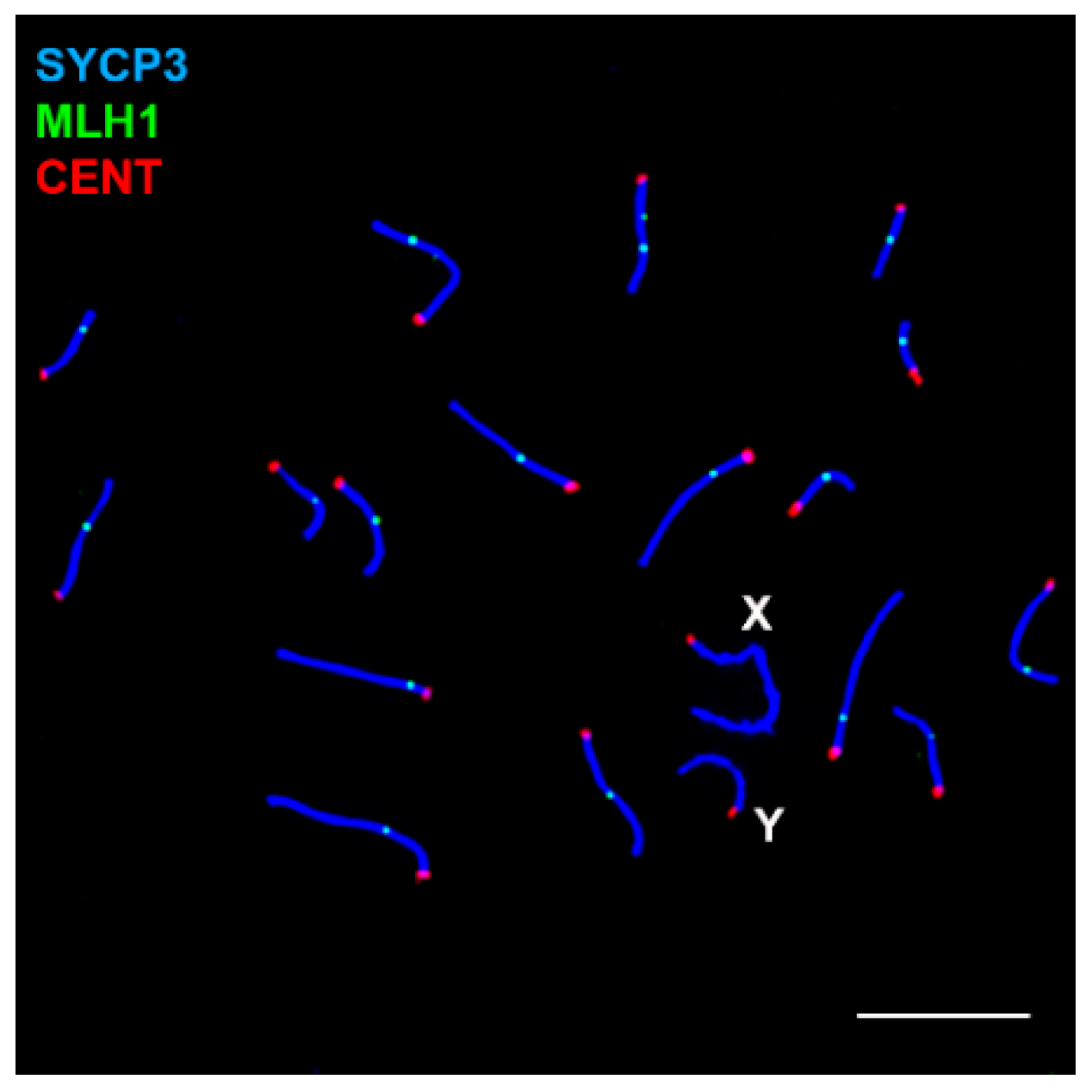
3.1. Sex Chromosome Pairing
3.2. DNA Repair/Recombination Dynamics in M. mattheyi
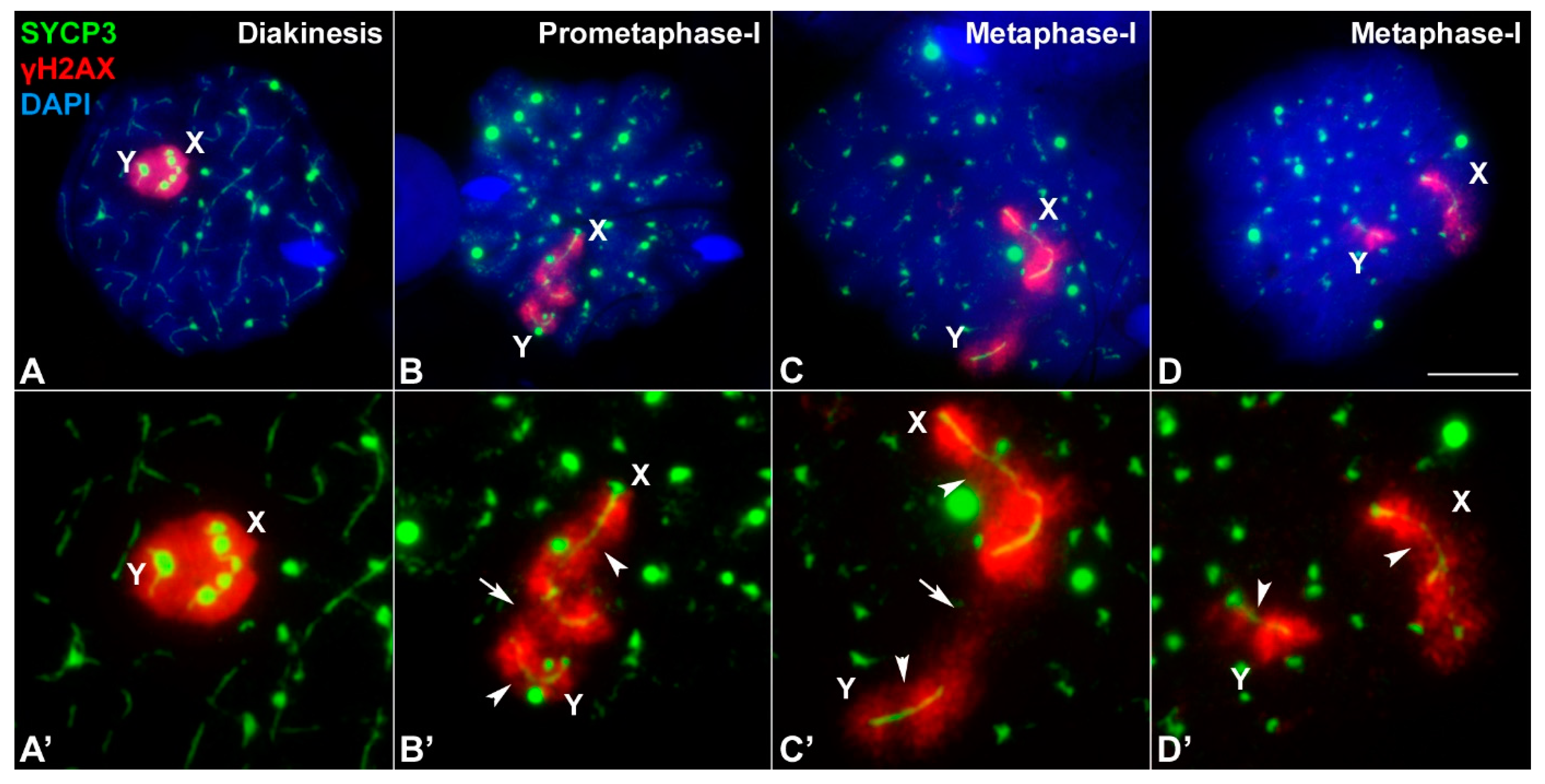
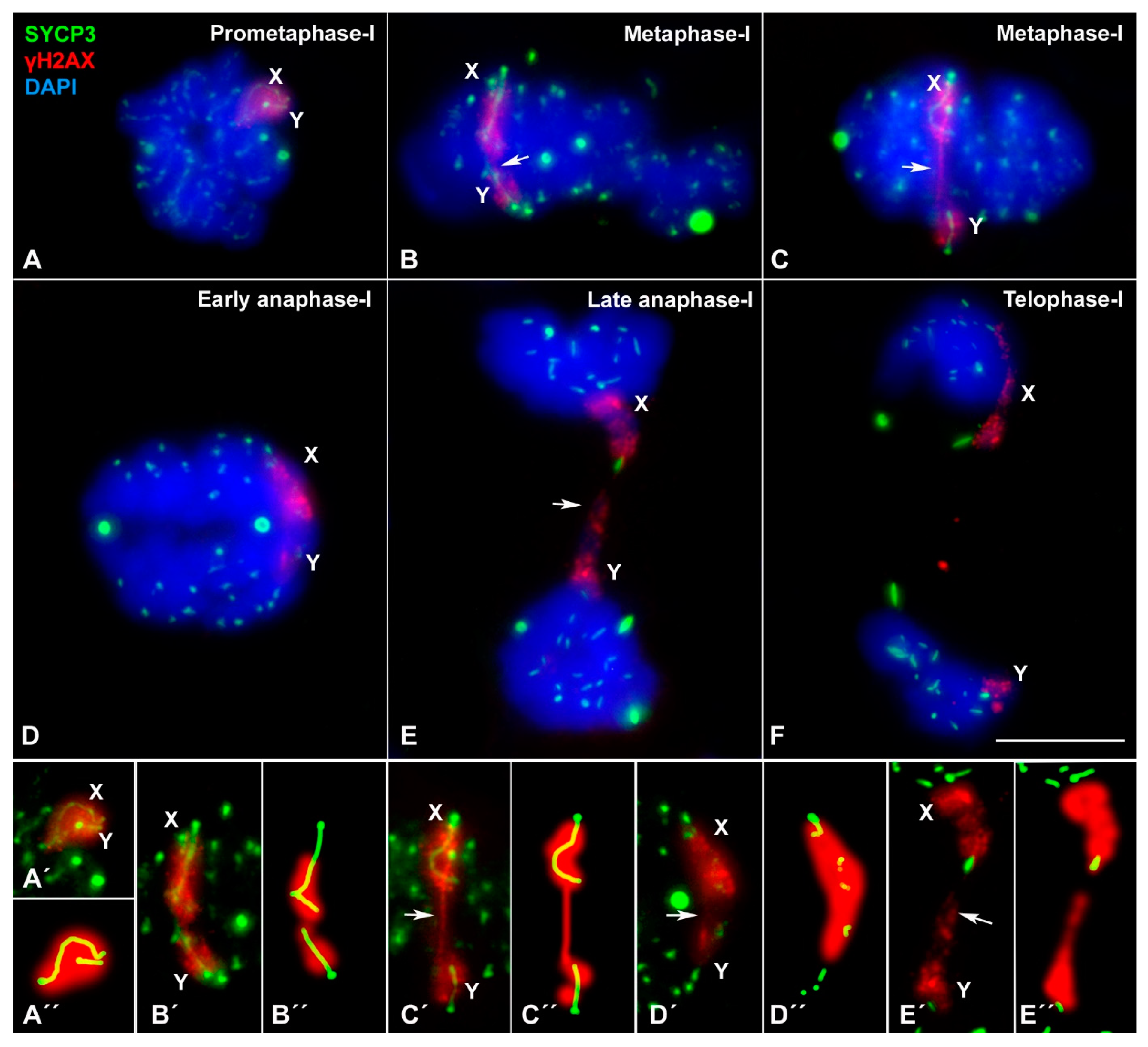
3.3. Sex Chromosome Segregation
4. Discussion
4.1. Origin of the Asynaptic Condition of Sex Chromosomes in M. mattheyi
4.2. DNA Damage and Repair in the Absence of Synapsis
4.3. Sex Chromosome Pairing in the Absence of Homologous Recombination
4.4. An Unreported Mechanism of Sex Chromosome Association at Metaphase-I
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- John, B. Meiosis; Cambridge University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Page, S.L.; Hawley, R.S. The genetics and molecular biology of the synaptonemal complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef]
- von Wettstein, D.; Rasmussen, S.W.; Holm, P.B. The synaptonemal complex in genetic segregation. Annu. Rev. Genet. 1984, 18, 331–413. [Google Scholar] [CrossRef]
- Keeney, S.; Neale, M.J. Initiation of meiotic recombination by formation of DNA double-strand breaks: Mechanism and regulation. Biochem. Soc. Trans. 2006, 34, 523–525. [Google Scholar] [CrossRef] [Green Version]
- Grey, C.; de Massy, B. Chromosome Organization in Early Meiotic Prophase. Front. Cell Dev. Biol. 2021, 9, 688878. [Google Scholar] [CrossRef] [PubMed]
- Zickler, D.; Kleckner, N. Meiotic chromosomes: Integrating structure and function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef] [PubMed]
- Zickler, D.; Kleckner, N. Recombination, pairing, and pynapsis of homologs during meiosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a016626. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.A.; Watson, J.M. Mammalian sex chromosomes: Evolution of organization and function. Chromosoma 1991, 101, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Sex Chromosome Specialization and Degeneration in Mammals. Cell 2006, 124, 901–914. [Google Scholar] [CrossRef] [Green Version]
- Burgoyne, P.S. Genetic homology and crossing over in the X and Y chromosomes of Mammals. Hum. Genet. 1982, 61, 85–90. [Google Scholar] [CrossRef]
- Goetz, P.; Chandley, A.C.; Speed, R.M. Morphological and temporal sequence of meiotic prophase development at puberty in the male mouse. J. Cell Sci. 1984, 65, 249–263. [Google Scholar] [CrossRef]
- Solari, A.J. The spatial relationship of the X and Y chromosomes during meiotic prophase in mouse spermatocytes. Chromosoma 1970, 29, 217–236. [Google Scholar] [CrossRef]
- Solari, A.J. The behavior of the XY pair in mammals. Int. Rev. Cytol. 1974, 38, 273–317. [Google Scholar]
- Page, J.; de la Fuente, R.; Manterola, M.; Parra, M.T.; Viera, A.; Berrios, S.; Fernandez-Donoso, R.; Rufas, J.S. Inactivation or non-reactivation: What accounts better for the silence of sex chromosomes during mammalian male meiosis? Chromosoma 2012, 121, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Page, J.; de la Fuente, R.; Gomez, R.; Calvente, A.; Viera, A.; Parra, M.T.; Santos, J.L.; Berrios, S.; Fernandez-Donoso, R.; Suja, J.A.; et al. Sex chromosomes, synapsis, and cohesins: A complex affair. Chromosoma 2006, 115, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Solari, A.J. Sex Chromosomes and Sex Determination in Vertebrates; CRC Press: Boca Raton, FL, USA, 1993; p. 308. [Google Scholar]
- Handel, M.A. The XY body: A specialized meiotic chromatin domain. Exp. Cell Res. 2004, 296, 57–63. [Google Scholar] [CrossRef]
- McKee, B.D.; Handel, M.A. Sex chromosomes, recombination, and chromatin conformation. Chromosoma 1993, 102, 71–80. [Google Scholar] [CrossRef]
- Turner, J.M. Meiotic sex chromosome inactivation. Development 2007, 134, 1823–1831. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.M.; Aprelikova, O.; Xu, X.; Wang, R.; Kim, S.; Chandramouli, G.V.; Barrett, J.C.; Burgoyne, P.S.; Deng, C.X. BRCA1, histone H2AX phosphorylation, and male meiotic sex chromosome inactivation. Curr. Biol. 2004, 14, 2135–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauppi, L.; Barchi, M.; Baudat, F.; Romanienko, P.J.; Keeney, S.; Jasin, M. Distinct properties of the XY pseudoautosomal region crucial for male meiosis. Science 2011, 331, 916–920. [Google Scholar] [CrossRef] [Green Version]
- Kauppi, L.; Jasin, M.; Keeney, S. The tricky path to recombining X and Y chromosomes in meiosis. Ann. N. Y. Acad. Sci. 2012, 1267, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acquaviva, L.; Boekhout, M.; Karasu, M.E.; Brick, K.; Pratto, F.; Li, T.; van Overbeek, M.; Kauppi, L.; Camerini-Otero, R.D.; Jasin, M.; et al. Ensuring meiotic DNA break formation in the mouse pseudoautosomal region. Nature 2020, 582, 426–431. [Google Scholar] [CrossRef]
- Boekhout, M.; Karasu, M.E.; Wang, J.; Acquaviva, L.; Pratto, F.; Brick, K.; Eng, D.Y.; Xu, J.; Camerini-Otero, R.D.; Patel, D.J.; et al. REC114 partner ANKRD31 controls number, timing, and location of meiotic DNA breaks. Mol. Cell 2019, 74, 1053–1068. [Google Scholar] [CrossRef]
- Papanikos, F.; Clement, J.A.J.; Testa, E.; Ravindranathan, R.; Grey, C.; Dereli, I.; Bondarieva, A.; Valerio-Cabrera, S.; Stanzione, M.; Schleiffer, A.; et al. Mouse ANKRD31 regulates spatiotemporal patterning of meiotic recombination initiation and ensures recombination between X and Y sex chromosomes. Mol. Cell 2019, 74, 1069–1085. [Google Scholar] [CrossRef] [PubMed]
- Ashley, T.; Plug, A.W.; Xu, J.; Solari, A.J.; Reddy, G.; Golub, E.I.; Ward, D.C. Dynamic changes in Rad51 distribution on chromatin during meiosis in male and female vertebrates. Chromosoma 1995, 104, 19–28. [Google Scholar] [CrossRef]
- Enguita-Marruedo, A.; Martin-Ruiz, M.; Garcia, E.; Gil-Fernandez, A.; Parra, M.T.; Viera, A.; Rufas, J.S.; Page, J. Transition from a meiotic to a somatic-like DNA damage response during the pachytene stage in mouse meiosis. PLoS Genet. 2019, 15, e1007439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, A.; Schoenmakers, S.; Baarends, W.M. DNA double strand break repair, chromosome synapsis and transcriptional silencing in meiosis. Epigenetics 2010, 5, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Ashley, T.; Moses, M.J. End association and segregation of the achiasmatic X and Y chromosomes of the sand rat, Psammomys obesus. Chromosoma 1980, 78, 203–210. [Google Scholar] [CrossRef]
- Borodin, P.M.; Basheva, E.A.; Torgasheva, A.A.; Dashkevich, O.A.; Golenishchev, F.N.; Kartavtseva, I.V.; Mekada, K.; Dumont, B.L. Multiple independent evolutionary losses of XY pairing at meiosis in the grey voles. Chromosome Res. 2012, 20, 259–268. [Google Scholar] [CrossRef]
- Borodin, P.M.; Sablina, O.V.; Rodionova, M.I. Pattern of X-Y chromosome pairing in microtine rodents. Hereditas 1995, 123, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Carnero, A.; Jimenez, R.; Burgos, M.; Sanchez, A.; Diaz de la Guardia, R. Achiasmatic sex chromosomes in Pitymys duodecimcostatus: Mechanisms of association and segregation. Cytogenet. Cell Genet. 1991, 56, 78–81. [Google Scholar] [CrossRef]
- Solari, A.J.; Ashley, T. Ultrastructure and behavior of the achiasmatic, telosynaptic XY pair of the sand rat (Psammomys obesus). Chromosoma 1977, 62, 319–336. [Google Scholar] [CrossRef]
- Wolf, K.W. How meiotic cells deal with non-exchange chromosomes. Bioessays 1994, 16, 107–114. [Google Scholar] [CrossRef]
- Jimenez, R.; Carnero, A.; Burgos, M.; Sanchez, A.; Diaz de la Guardia, R. Achiasmatic giant sex chromosomes in the vole Microtus cabrerae (Rodentia, Microtidae). Cytogenet. Cell Genet. 1991, 57, 56–58. [Google Scholar]
- Wolf, K.W.; Baumgart, K.; Winking, H. Meiotic association and segregation of the giant sex chromosomes in male field vole (Microtus agrestis). Chromosoma 1988, 97, 124–133. [Google Scholar] [CrossRef]
- Page, J.; Berrios, S.; Parra, M.T.; Viera, A.; Suja, J.A.; Prieto, I.; Barbero, J.L.; Rufas, J.S.; Fernandez-Donoso, R. The program of sex chromosome pairing in meiosis is highly conserved across marsupial species: Implications for sex chromosome evolution. Genetics 2005, 170, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Page, J.; Berrios, S.; Rufas, J.S.; Parra, M.T.; Suja, J.A.; Heyting, C.; Fernandez-Donoso, R. The pairing of X and Y chromosomes during meiotic prophase in the marsupial species Thylamys elegans is maintained by a dense plate developed from their axial elements. J. Cell Sci. 2003, 116, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Roche, L.; Seluja, G.; Wettstein, R. The meiotic behaviour of the XY pair in Lutreolina crassicaudata (Marsupialia: Didelphoidea). Genetica 1986, 71, 213–224. [Google Scholar] [CrossRef]
- Seluja, G.; Roche, L.; Solari, A.J. Male meiotic prophase in Didelphis albiventris: A comparative cytological and electron microscopical study. J. Hered. 1987, 78, 218–222. [Google Scholar] [CrossRef]
- Sharp, P. Sex chromosome pairing during male meiosis in marsupials. Chromosoma 1982, 86, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Solari, A.J.; Bianchi, N.O. The synaptic behaviour of the X and Y chromosomes in the marsupial Monodelphis dimidiata. Chromosoma 1975, 52, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Page, J.; Viera, A.; Parra, M.T.; de la Fuente, R.; Suja, J.A.; Prieto, I.; Barbero, J.L.; Rufas, J.S.; Berrios, S.; Fernandez-Donoso, R. Involvement of synaptonemal complex proteins in sex chromosome segregation during marsupial male meiosis. PLoS Genet. 2006, 2, e136. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Donoso, R.; Berríos, S.; Rufas, J.S.; Page, J. Marsupial sex chromosome behaviour during male meiosis. In Marsupial Genetics and Genomics; Deakin, J.A., Waters, P.D., Marshall Graves, J.A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 187–206. [Google Scholar]
- de la Fuente, R.; Parra, M.T.; Viera, A.; Calvente, A.; Gomez, R.; Suja, J.A.; Rufas, J.S.; Page, J. Meiotic pairing and segregation of achiasmate sex chromosomes in eutherian mammals: The role of SYCP3 protein. PLoS Genet. 2007, 3, e198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente, R.; Sanchez, A.; Marchal, J.A.; Viera, A.; Parra, M.T.; Rufas, J.S.; Page, J. A synaptonemal complex-derived mechanism for meiotic segregation precedes the evolutionary loss of homology between sex chromosomes in arvicolid mammals. Chromosoma 2012, 121, 433–446. [Google Scholar] [CrossRef]
- Macholán, M. Multivariate Analysis of Morphometric Variation in Asian Mus and Sub-Saharan Nannomys (Rodentia: Muridae). Zool. Anz. 2001, 240, 7–14. [Google Scholar] [CrossRef]
- Catzeflis, F.M.; Denys, C. The african Nannomys (Muridae): An early offshoot from the Mus lineage-Evidence from scnDNA hybridization experiments and compared morphology. Isr. J. Zool. 1992, 38, 219–231. [Google Scholar] [CrossRef]
- Happold, D.C.; Kingdon, J.; Happold, M. Mammals of Africa/Volume III, Rodents, Hares and Rabbits; Happold, D.C., Ed.; Bloomsbury: London, UK, 2013. [Google Scholar]
- Colomina, V.; Catalan, J.; Britton-Davidian, J.; Veyrunes, F. Extensive amplification of telomeric repeats in the karyotypically highly diverse African pygmy mice. Cytogenet. Genome Res. 2017, 152, 55–64. [Google Scholar] [CrossRef]
- Petter, F. Une souris nouvelle d’Afrique occidentale Mus mattheyi sp. nov. Mammalia 1969, 33, 118–123. [Google Scholar] [CrossRef]
- Veyrunes, F.; Catalan, J.; Sicard, B.; Robinson, T.J.; Duplantier, J.-M.; Granjon, L.; Dobigny, G.; Britton-Davidian, J. Autosome and sex chromosome diversity among the African pygmy mice, subgenus Nannomys (Murinae; Mus). Chromosome Res. 2004, 12, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Matthey, R. Nouvelles contributions à la cytogénétique des Mus Africains du sous-genre Leggada. Experientia 1966, 22, 400–401. [Google Scholar] [CrossRef]
- Britton-Davidian, J.; Robinson, T.J.; Veyrunes, F. Systematics and evolution of the African pygmy mice, subgenus Nannomys: A review. Acta Oecol. 2012, 42, 41–49. [Google Scholar] [CrossRef]
- Gil-Fernandez, A.; Saunders, P.A.; Martin-Ruiz, M.; Ribagorda, M.; Lopez-Jimenez, P.; Jeffries, D.L.; Parra, M.T.; Viera, A.; Rufas, J.S.; Perrin, N.; et al. Meiosis reveals the early steps in the evolution of a neo-XY sex chromosome pair in the African pygmy mouse Mus minutoides. PLoS Genet. 2020, 16, e1008959. [Google Scholar] [CrossRef]
- Jotterand-Bellomo, M. Le caryotype et la spermatogénèse de Mus setulosus (bandes Q, C, G et coloration argentique). Genetica 2004, 56, 217–227. [Google Scholar] [CrossRef]
- Baudat, F.; de Massy, B.; Veyrunes, F. Sex chromosome quadrivalents in oocytes of the African pygmy mouse Mus minutoides that harbors non-conventional sex chromosomes. Chromosoma 2019, 128, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Saunders, P.A.; Perez, J.; Rahmoun, M.; Ronce, O.; Crochet, P.-A.; Veyrunes, F. XY females do better than the XX in the African pygmy mouse Mus minutoides. Evolution 2014, 68, 2119–2127. [Google Scholar] [CrossRef]
- Peters, A.H.; Plug, A.W.; van Vugt, M.J.; de Boer, P. A drying-down technique for the spreading of mammalian meiocytes from the male and female germline. Chromosome Res. 1997, 5, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Page, J.; Suja, J.A.; Santos, J.L.; Rufas, J.S. Squash procedure for protein immunolocalization in meiotic cells. Chromosome Res. 1998, 6, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Plug, A.W.; Peters, A.H.; Keegan, K.S.; Hoekstra, M.F.; de Boer, P.; Ashley, T. Changes in protein composition of meiotic nodules during mammalian meiosis. J. Cell Sci. 1998, 111, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Xue, J.; Yin, H.; Guo, R.; Luo, M.; Ye, L.; Shi, Q.; Huang, X.; Liu, M.; Sha, J.; et al. Dual functions for the ssDNA-binding protein RPA in meiotic recombination. PLoS Genet. 2019, 15, e1007952. [Google Scholar] [CrossRef] [Green Version]
- Moens, P.B.; Kolas, N.K.; Tarsounas, M.; Marcon, E.; Cohen, P.E.; Spyropoulos, B. The time course and chromosomal localization of recombination-related proteins at meiosis in the mouse are compatible with models that can resolve the early DNA-DNA interactions without reciprocal recombination. J. Cell Sci. 2002, 115, 1611–1622. [Google Scholar] [CrossRef]
- Oliver-Bonet, M.; Turek, P.J.; Sun, F.; Ko, E.; Martin, R.H. Temporal progression of recombination in human males. Mol. Hum. Reprod. 2005, 11, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra, M.T.; Viera, A.; Gomez, R.; Page, J.; Benavente, R.; Santos, J.L.; Rufas, J.S.; Suja, J.A. Involvement of the cohesin Rad21 and SCP3 in monopolar attachment of sister kinetochores during mouse meiosis I. J. Cell Sci. 2004, 117, 1221–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grutzner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef]
- Bellott, D.W.; Hughes, J.F.; Skaletsky, H.; Brown, L.G.; Pyntikova, T.; Cho, T.J.; Koutseva, N.; Zaghlul, S.; Graves, T.; Rock, S.; et al. Mammalian Y chromosomes retain widely expressed dosage-sensitive regulators. Nature 2014, 508, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Gil-Fernández, A.; Matveevsky, S.; Martín-Ruiz, M.; Ribagorda, M.; Parra, M.T.; Viera, A.; Rufas, J.S.; Kolomiets, O.; Bakloushinskaya, I.; Page, J. Sex differences in the meiotic behavior of an XX sex chromosome pair in males and females of the mole vole Ellobius tancrei: Turning an X into a Y chromosome? Chromosoma 2021, 113–131. [Google Scholar] [CrossRef] [PubMed]
- Torgasheva, A.; Malinovskaya, L.; Zadesenets, K.S.; Slobodchikova, A.; Shnaider, E.; Rubtsov, N.; Borodin, P. Highly conservative pattern of sex chromosome synapsis and recombination in neognathae birds. Genes 2021, 12, 1358. [Google Scholar] [CrossRef]
- Pajpach, F.; Wu, T.; Shearwin-Whyatt, L.; Jones, K.; Grützner, F. Flavors of non-random meiotic segregation of autosomes and sex chromosomes. Genes 2021, 12, 1338. [Google Scholar] [CrossRef]
- Waters, P.D.; Ruiz-Herrera, A. Meiotic executioner genes protect the Y from extinction. Trends Genet. 2020, 36, 728–738. [Google Scholar] [CrossRef]
- Solari, A.J. The evolution of the ultrastructure of the sex chromosomes (sex vesicle) during meiotic prophase in mouse spermatocytes. J. Ultrastruct. Res. 1969, 27, 289–305. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Sutcliffe, M.J.; Palmer, S.J. Fertility in mice requires X-Y pairing and a Y-chromosomal “spermiogenesis” gene mapping to the long arm. Cell 1992, 71, 391–398. [Google Scholar] [CrossRef]
- De Vries, F.A.; de Boer, E.; van den Bosch, M.; Baarends, W.M.; Ooms, M.; Yuan, L.; Liu, J.G.; van Zeeland, A.A.; Heyting, C.; Pastink, A. Mouse Sycp1 functions in synaptonemal complex assembly, meiotic recombination, and XY body formation. Genes. Dev. 2005, 19, 1376–1389. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Capetillo, O.; Mahadevaiah, S.K.; Celeste, A.; Romanienko, P.J.; Camerini-Otero, R.D.; Bonner, W.M.; Manova, K.; Burgoyne, P.; Nussenzweig, A. H2AX is required for chromatin remodeling and inactivation of sex chromosomes in male mouse meiosis. Dev. Cell 2003, 4, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Handel, M.A. The XY body: An attractive chromatin domain. Biol. Reprod. 2020, 102, 985–987. [Google Scholar] [CrossRef]
- Turner, J.M.A. Meiotic Silencing, Infertility and X Chromosome Evolution. In Epigenomics; Ferguson-Smith, A.C., Greally, J.M., Martienssen, R.A., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 301–318. [Google Scholar]
- Ashley, T.; Jaarola, M.; Fredga, K. Absence of synapsis during pachynema of the normal sized sex chromosomes of Microtus arvalis. Hereditas 1989, 111, 295–304. [Google Scholar] [CrossRef]
- Graves, J.A. Mammals that break the rules: Genetics of marsupials and monotremes. Annu. Rev. Genet. 1996, 30, 233–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayman, D.L. Marsupial cytogenetics. Aust. J. Zool. 1990, 37, 331–349. [Google Scholar] [CrossRef]
- Charlesworth, B.; Charlesworth, D. The degeneration of Y chromosomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1563–1572. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, W.R. Evolution of the Y Sex Chromosome in Animals: Y chromosomes evolve through the degeneration of autosomes. BioScience 1996, 46, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.A.; Wakefield, M.J.; Toder, R. The origin and evolution of the pseudoautosomal regions of human sex chromosomes. Hum. Mol. Genet. 1998, 7, 1991–1996. [Google Scholar] [CrossRef] [Green Version]
- Dumont, B.L.; Williams, C.L.; Ng, B.L.; Horncastle, V.; Chambers, C.L.; McGraw, L.A.; Adams, D.; Mackay, T.F.C.; Breen, M. Relationship Between Sequence Homology, Genome Architecture, and Meiotic Behavior of the Sex Chromosomes in North American Voles. Genetics 2018, 210, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Veyrunes, F.; Britton-Davidian, J.; Robinson, T.J.; Calvet, E.; Denys, C.; Chevret, P. Molecular phylogeny of the African pygmy mice, subgenus Nannomys (Rodentia, Murinae, Mus): Implications for chromosomal evolution. Mol. Phylogenet. Evol. 2005, 36, 358–369. [Google Scholar] [CrossRef]
- Viegas-Péquignot, E.; Benazzou, T.; Dutrillaux, B.; Petter, F. Complex evolution of sex chromosomes in Gerbillidae (Rodentia). Cytogenet. Cell Genet. 1982, 34, 158–167. [Google Scholar] [CrossRef]
- Deakin, J.E. Chromosome evolution in marsupials. Genes 2018, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladkikh, O.L.; Romanenko, S.A.; Lemskaya, N.A.; Serdyukova, N.A.; O’Brien, P.C.; Kovalskaya, J.M.; Smorkatcheva, A.V.; Golenishchev, F.N.; Perelman, P.L.; Trifonov, V.A.; et al. Rapid karyotype evolution in Lasiopodomys involved at least two autosome-sex chromosome translocations. PLoS ONE 2016, 11, e0167653. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Turner, J.M. The consequences of asynapsis for mammalian meiosis. Nat. Rev. Genet. 2009, 10, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Faisal, I.; Kauppi, L. Sex chromosome recombination failure, apoptosis, and fertility in male mice. Chromosoma 2016, 125, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Checchi, P.M.; Lawrence, K.S.; Van, M.V.; Larson, B.J.; Engebrecht, J. Pseudosynapsis and decreased stringency of meiotic repair pathway choice on the hemizygous sex chromosome of Caenorhabditis elegans males. Genetics 2014, 197, 543–560. [Google Scholar] [CrossRef] [Green Version]
- Enguita-Marruedo, A.; Van Cappellen, W.A.; Hoogerbrugge, J.W.; Carofiglio, F.; Wassenaar, E.; Slotman, J.A.; Houtsmuller, A.; Baarends, W.M. Live cell analyses of synaptonemal complex dynamics and chromosome movements in cultured mouse testis tubules and embryonic ovaries. Chromosoma 2018, 127, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Scherthan, H.; Weich, S.; Schwegler, H.; Heyting, C.; Harle, M.; Cremer, T. Centromere and telomere movements during early meiotic prophase of mouse and man are associated with the onset of chromosome pairing. J. Cell Biol. 1996, 134, 1109–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer-Fender, S. Molecular aspects of XY body formation. Cytogenet. Genome Res. 2003, 103, 245–255. [Google Scholar]
- van der Heijden, G.W.; Derijck, A.A.; Posfai, E.; Giele, M.; Pelczar, P.; Ramos, L.; Wansink, D.G.; van der Vlag, J.; Peters, A.H.; de Boer, P. Chromosome-wide nucleosome replacement and H3.3 incorporation during mammalian meiotic sex chromosome inactivation. Nat. Genet. 2007, 39, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Faisal, I.; Kauppi, L. Reduced MAD2 levels dampen the apoptotic response to non-exchange sex chromosomes and lead to sperm aneuploidy. Development 2017, 144, 1988–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veyrunes, F.; Perez, J.; Borremans, B.; Gryseels, S.; Richards, L.R.; Duran, A.; Chevret, P.; Robinson, T.J.; Britton-Davidian, J. A new cytotype of the African pygmy mouse Mus minutoides in Eastern Africa. Implications for the evolution of sex-autosome translocations. Chromosome Res. 2014, 22, 533–543. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Fernández, A.; Ribagorda, M.; Martín-Ruiz, M.; López-Jiménez, P.; Laguna, T.; Gómez, R.; Parra, M.T.; Viera, A.; Veyrunes, F.; Page, J. Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals. Genes 2021, 12, 1434. https://doi.org/10.3390/genes12091434
Gil-Fernández A, Ribagorda M, Martín-Ruiz M, López-Jiménez P, Laguna T, Gómez R, Parra MT, Viera A, Veyrunes F, Page J. Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals. Genes. 2021; 12(9):1434. https://doi.org/10.3390/genes12091434
Chicago/Turabian StyleGil-Fernández, Ana, Marta Ribagorda, Marta Martín-Ruiz, Pablo López-Jiménez, Tamara Laguna, Rocío Gómez, María Teresa Parra, Alberto Viera, Frederic Veyrunes, and Jesús Page. 2021. "Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals" Genes 12, no. 9: 1434. https://doi.org/10.3390/genes12091434
APA StyleGil-Fernández, A., Ribagorda, M., Martín-Ruiz, M., López-Jiménez, P., Laguna, T., Gómez, R., Parra, M. T., Viera, A., Veyrunes, F., & Page, J. (2021). Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals. Genes, 12(9), 1434. https://doi.org/10.3390/genes12091434