
Epigenetic Silencing of Tumor Suppressor lncRNA NKILA: Implication on NF-κB Signaling in Non-Hodgkin’s Lymphoma


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Cell Culture
2.3. DNA Demethylation Treatment
2.4. DNA Extraction
2.5. Methylation-Specific Polymerase Chain Reaction (MSP)
2.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.7. Quantitative Bisulfite Pyrosequencing
2.8. Knockdown of NKILA
2.9. Cell Proliferation and Cell Death
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. NKILA Was Methylated in a Tumor-Specific Manner in NHL Cells
3.2. The Expression of NKILA Was Inversely Correlated with Promoter DNA Methylation
3.3. NKILA Was Differentially Methylated in NHL Primary Samples
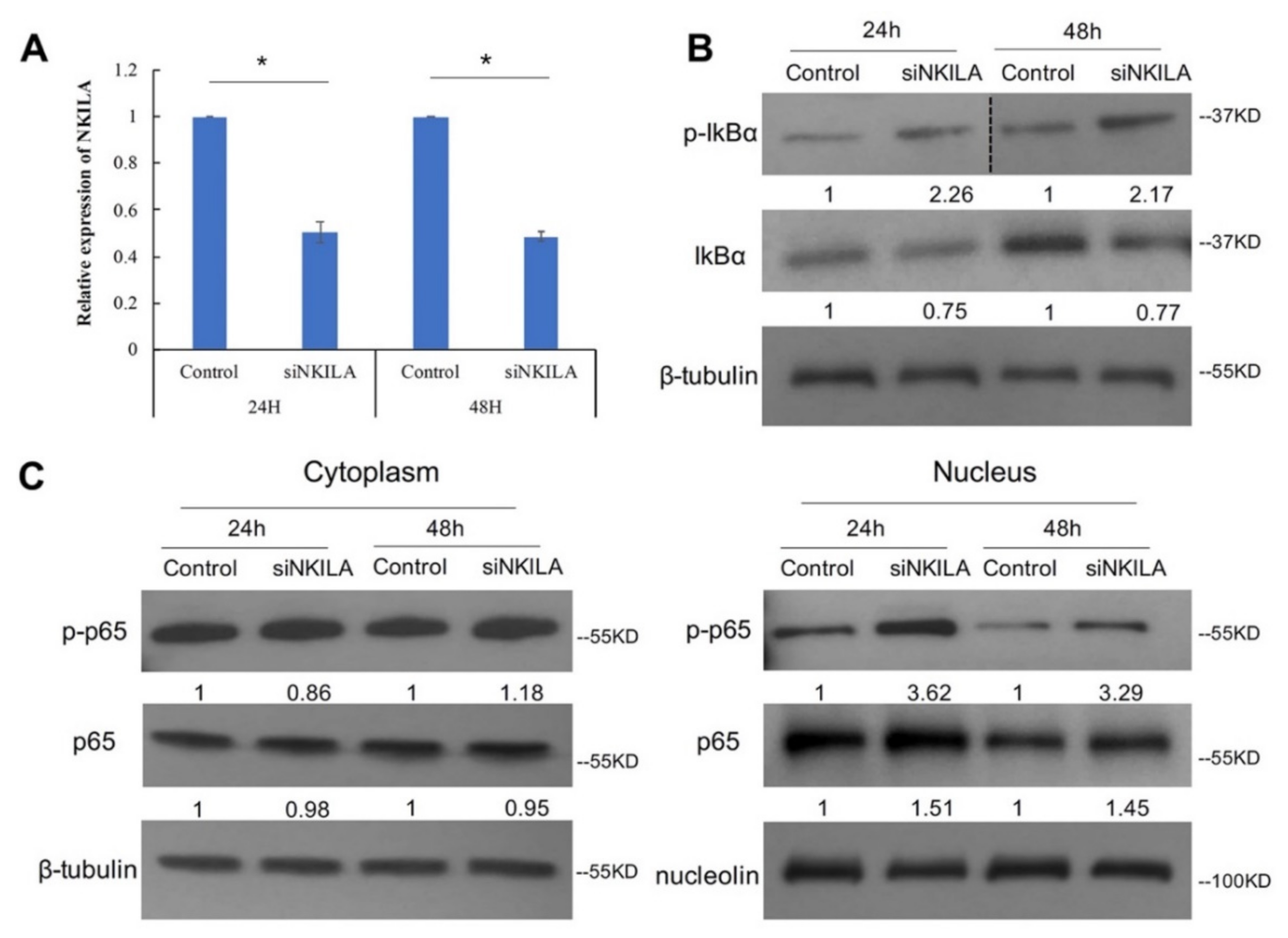
3.4. NKILA Inhibited IkBα Phosphorylation and NF-κB Activation
3.5. NKILA Was a Tumor Suppressor lncRNA in SU-DHL-1 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [Green Version]
- Au, W.Y.; Ma, S.Y.; Chim, C.S.; Choy, C.; Loong, F.; Lie, A.K.; Lam, C.C.; Leung, A.Y.; Tse, E.; Yau, C.C.; et al. Clinicopathologic features and treatment outcome of mature T-cell and natural killer-cell lymphomas diagnosed according to the World Health Organization classification scheme: A single center experience of 10 years. Ann. Oncol. 2005, 16, 206–214. [Google Scholar] [CrossRef]
- Bowzyk Al-Naeeb, A.; Ajithkumar, T.; Behan, S.; Hodson, D.J. Non-Hodgkin lymphoma. BMJ 2018, 362, k3204. [Google Scholar] [CrossRef]
- Chim, C.S.; Ma, S.Y.; Au, W.Y.; Choy, C.; Lie, A.K.; Liang, R.; Yau, C.C.; Kwong, Y.L. Primary nasal natural killer cell lymphoma: Long-term treatment outcome and relationship with the International Prognostic Index. Blood 2004, 103, 216–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Shankland, K.R.; Armitage, J.O.; Hancock, B.W. Non-Hodgkin lymphoma. Lancet 2012, 380, 848–857. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef]
- Klutstein, M.; Nejman, D.; Greenfield, R.; Cedar, H. DNA Methylation in Cancer and Aging. Cancer Res. 2016, 76, 3446–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, J.; Li, Z.; Zhu, M.; Zhang, Z.; Wang, Y.; Jing, H. Promoter hypermethylation of PTPL1, PTPN6, DAPK, p16 and 5-azacitidine inhibits growth in DLBCL. Oncol. Rep. 2016, 35, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Wong, K.Y.; Loong, F.; Srivastava, G. SOCS1 and SHP1 hypermethylation in mantle cell lymphoma and follicular lymphoma: Implications for epigenetic activation of the Jak/STAT pathway. Leukemia 2004, 18, 356–358. [Google Scholar] [CrossRef] [Green Version]
- Molavi, O.; Wang, P.; Zak, Z.; Gelebart, P.; Belch, A.; Lai, R. Gene methylation and silencing of SOCS3 in mantle cell lymphoma. Br. J. Haematol. 2013, 161, 348–356. [Google Scholar] [CrossRef]
- Yim, R.L.; Wong, K.Y.; Kwong, Y.L.; Loong, F.; Leung, C.Y.; Chu, R.; Lam, W.W.; Hui, P.K.; Lai, R.; Chim, C.S. Methylation of miR-155-3p in mantle cell lymphoma and other non-Hodgkin’s lymphomas. Oncotarget 2014, 5, 9770–9782. [Google Scholar] [CrossRef] [Green Version]
- Paik, J.H.; Jang, J.Y.; Jeon, Y.K.; Kim, W.Y.; Kim, T.M.; Heo, D.S.; Kim, C.W. MicroRNA-146a downregulates NFkappaB activity via targeting TRAF6 and functions as a tumor suppressor having strong prognostic implications in NK/T cell lymphoma. Clin. Cancer Res. 2011, 17, 4761–4771. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Spizzo, R.; Almeida, M.I.; Colombatti, A.; Calin, G.A. Long non-coding RNAs and cancer: A new frontier of translational research? Oncogene 2012, 31, 4577–4587. [Google Scholar] [CrossRef] [Green Version]
- Grote, P.; Herrmann, B.G. Long noncoding RNAs in organogenesis: Making the difference. Trends Genet. 2015, 31, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, L.; Liu, Q.; Gong, C.; Yao, Y.; Lv, X.; Lin, L.; Yao, H.; Su, F.; Li, D.; et al. A cytoplasmic NF-kappaB interacting long noncoding RNA blocks IkappaB phosphorylation and suppresses breast cancer metastasis. Cancer Cell 2015, 27, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; International Agency for Research on Cancer: Lyon, France, 2008. [Google Scholar]
- Chim, C.S.; Wong, K.Y.; Qi, Y.; Loong, F.; Lam, W.L.; Wong, L.G.; Jin, D.Y.; Costello, J.F.; Liang, R. Epigenetic inactivation of the miR-34a in hematological malignancies. Carcinogenesis 2010, 31, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Li, Y.; Wang, J.; Che, Y.; Sun, S.; Huang, J.; Chen, Z.; He, J. Long non-coding RNA NKILA inhibits migration and invasion of non-small cell lung cancer via NF-kappaB/Snail pathway. J. Exp. Clin. Cancer Res. 2017, 36, 54. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, Q.; Liu, G.; Zheng, F.; Chen, J.; Huang, D.; Ding, L.; Yang, X.; Song, E.; Xiang, Y.; et al. NKILA represses nasopharyngeal carcinoma carcinogenesis and metastasis by NF-kappaB pathway inhibition. PLoS Genet. 2019, 15, e1008325. [Google Scholar] [CrossRef]
- Chim, C.S.; Wong, K.Y.; Loong, F.; Lam, W.W.; Srivastava, G. Frequent epigenetic inactivation of Rb1 in addition to p15 and p16 in mantle cell and follicular lymphoma. Hum. Pathol. 2007, 38, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Calin, G.A.; Yuen, K.S.; Jin, D.Y.; Chim, C.S. Epigenetic silencing of miR-342-3p in B cell lymphoma and its impact on autophagy. Clin. Epigenet. 2020, 12, 150. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Wang, L.Q.; Chim, C.S. miR-1250-5p is a novel tumor suppressive intronic miRNA hypermethylated in non-Hodgkin’s lymphoma: Novel targets with impact on ERK signaling and cell migration. Cell Commun. Signal. 2021, 19, 62. [Google Scholar] [CrossRef] [PubMed]
- Lujambio, A.; Ropero, S.; Ballestar, E.; Fraga, M.F.; Cerrato, C.; Setien, F.; Casado, S.; Suarez-Gauthier, A.; Sanchez-Cespedes, M.; Git, A.; et al. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res. 2007, 67, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Liang, G.; Egger, G.; Friedman, J.M.; Chuang, J.C.; Coetzee, G.A.; Jones, P.A. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 2006, 9, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Subero, J.I.; Ammerpohl, O.; Bibikova, M.; Wickham-Garcia, E.; Agirre, X.; Alvarez, S.; Bruggemann, M.; Bug, S.; Calasanz, M.J.; Deckert, M.; et al. A comprehensive microarray-based DNA methylation study of 367 hematological neoplasms. PLoS ONE 2009, 4, e6986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Chen, J.; Yang, L.; Ouyang, Q.; Li, J.; Lao, L.; Zhao, J.; Liu, J.; Lu, Y.; Xing, Y.; et al. NKILA lncRNA promotes tumor immune evasion by sensitizing T cells to activation-induced cell death. Nat. Immunol. 2018, 19, 1112–1125. [Google Scholar] [CrossRef]
- Nagel, D.; Vincendeau, M.; Eitelhuber, A.C.; Krappmann, D. Mechanisms and consequences of constitutive NF-kappaB activation in B-cell lymphoid malignancies. Oncogene 2014, 33, 5655–5665. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.F.; Williams, S.A.; Mu, Y.; Nakano, H.; Duerr, J.M.; Buckbinder, L.; Greene, W.C. NF-kappaB RelA phosphorylation regulates RelA acetylation. Mol. Cell Biol. 2005, 25, 7966–7975. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Choi, G.E.; Ryu, S.; Kwon, S.J.; Kim, S.C.; Booth, C.; Nichols, K.E.; Kim, H.S. Stepwise phosphorylation of p65 promotes NF-kappaB activation and NK cell responses during target cell recognition. Nat. Commun. 2016, 7, 11686. [Google Scholar] [CrossRef]
- Sakurai, H.; Chiba, H.; Miyoshi, H.; Sugita, T.; Toriumi, W. IkappaB kinases phosphorylate NF-kappaB p65 subunit on serine 536 in the transactivation domain. J. Biol. Chem. 1999, 274, 30353–30356. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Greten, F.R. NF-kappaB: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005, 5, 749–759. [Google Scholar] [CrossRef]
- Bian, D.; Gao, C.; Bao, K.; Song, G. The long non-coding RNA NKILA inhibits the invasion-metastasis cascade of malignant melanoma via the regulation of NF-kB. Am. J. Cancer Res. 2017, 7, 28–40. [Google Scholar]
- Tao, F.; Xu, Y.; Yang, D.; Tian, B.; Jia, Y.; Hou, J.; Dong, M. LncRNA NKILA correlates with the malignant status and serves as a tumor-suppressive role in rectal cancer. J. Cell Biochem. 2018, 119, 9809–9816. [Google Scholar] [CrossRef]
- Yang, T.; Li, S.; Liu, J.; Yin, D.; Yang, X.; Tang, Q. lncRNA-NKILA/NF-kappaB feedback loop modulates laryngeal cancer cell proliferation, invasion, and radioresistance. Cancer Med. 2018, 7, 2048–2063. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Tang, W.; Yang, Y.; Tang, L.; Dai, R.; Pu, B.; Feng, C.; Xia, J. Long noncoding RNA NKILA enhances the anti-cancer effects of baicalein in hepatocellular carcinoma via the regulation of NF-kappaB signaling. Chem. Biol. Interact. 2018, 285, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cui, X.; Chen, J.; Feng, Y.; Song, E.; Li, J.; Liu, Y. Long non-coding RNA NKILA inhibits migration and invasion of tongue squamous cell carcinoma cells via suppressing epithelial-mesenchymal transition. Oncotarget 2016, 7, 62520–62532. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | Tm/Cycles/ 2 | Reference | |
|---|---|---|---|---|
| Methylation-Specific PCR (MSP) | ||||
| M-MSP | TAG GTA GAC GGT TTG ACG TTA GC | GAA AAA ACC TCG ACG AAA ATT AAC G | 57 °C/35 x/2 mM | |
| U-MSP | GGT AGG TAG ATG GTT TGA TGT TAG T | ACA AAA AAA CCT CAA CAA AAA TTA ACA | 55 °C/37 x/1.5 mM | |
| Semi-Quantitative RT-PCR/Quantitative Real-Time RT-PCR | ||||
| NKILA | AAC CAA ACC TAC CCA CAA CG | ACC ACT AAG TCA ATC CCA GGT G | 55 °C/40 x/2 mM(Semi-Quantitative RT-PCR) | [19] |
| GAPDH | ACC ACA GTC CAT GCC ATC ACT | TCC ACC ACC CTG TTG CTG TA | 60 °C/24 x/1.5 mM(Semi-Quantitative RT-PCR) | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.-Y.; Calin, G.; Deng, M.-D.; Au-Yeung, R.K.H.; Wang, L.-Q.; Chim, C.-S. Epigenetic Silencing of Tumor Suppressor lncRNA NKILA: Implication on NF-κB Signaling in Non-Hodgkin’s Lymphoma. Genes 2022, 13, 128. https://doi.org/10.3390/genes13010128
Zhang M-Y, Calin G, Deng M-D, Au-Yeung RKH, Wang L-Q, Chim C-S. Epigenetic Silencing of Tumor Suppressor lncRNA NKILA: Implication on NF-κB Signaling in Non-Hodgkin’s Lymphoma. Genes. 2022; 13(1):128. https://doi.org/10.3390/genes13010128
Chicago/Turabian StyleZhang, Min-Yue, George Calin, Ming-Dan Deng, Rex K. H. Au-Yeung, Lu-Qian Wang, and Chor-Sang Chim. 2022. "Epigenetic Silencing of Tumor Suppressor lncRNA NKILA: Implication on NF-κB Signaling in Non-Hodgkin’s Lymphoma" Genes 13, no. 1: 128. https://doi.org/10.3390/genes13010128
APA StyleZhang, M. -Y., Calin, G., Deng, M. -D., Au-Yeung, R. K. H., Wang, L. -Q., & Chim, C. -S. (2022). Epigenetic Silencing of Tumor Suppressor lncRNA NKILA: Implication on NF-κB Signaling in Non-Hodgkin’s Lymphoma. Genes, 13(1), 128. https://doi.org/10.3390/genes13010128