

Confounding Factors Impacting microRNA Expression in Human Saliva: Methodological and Biological Considerations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Data and Sample Collection
2.3. Sample Handling and RNA Processing
2.4. Statistical Analysis
3. Results
3.1. Patient Demographics and Sample Attributes
3.2. Salivary miRNA Expression Results
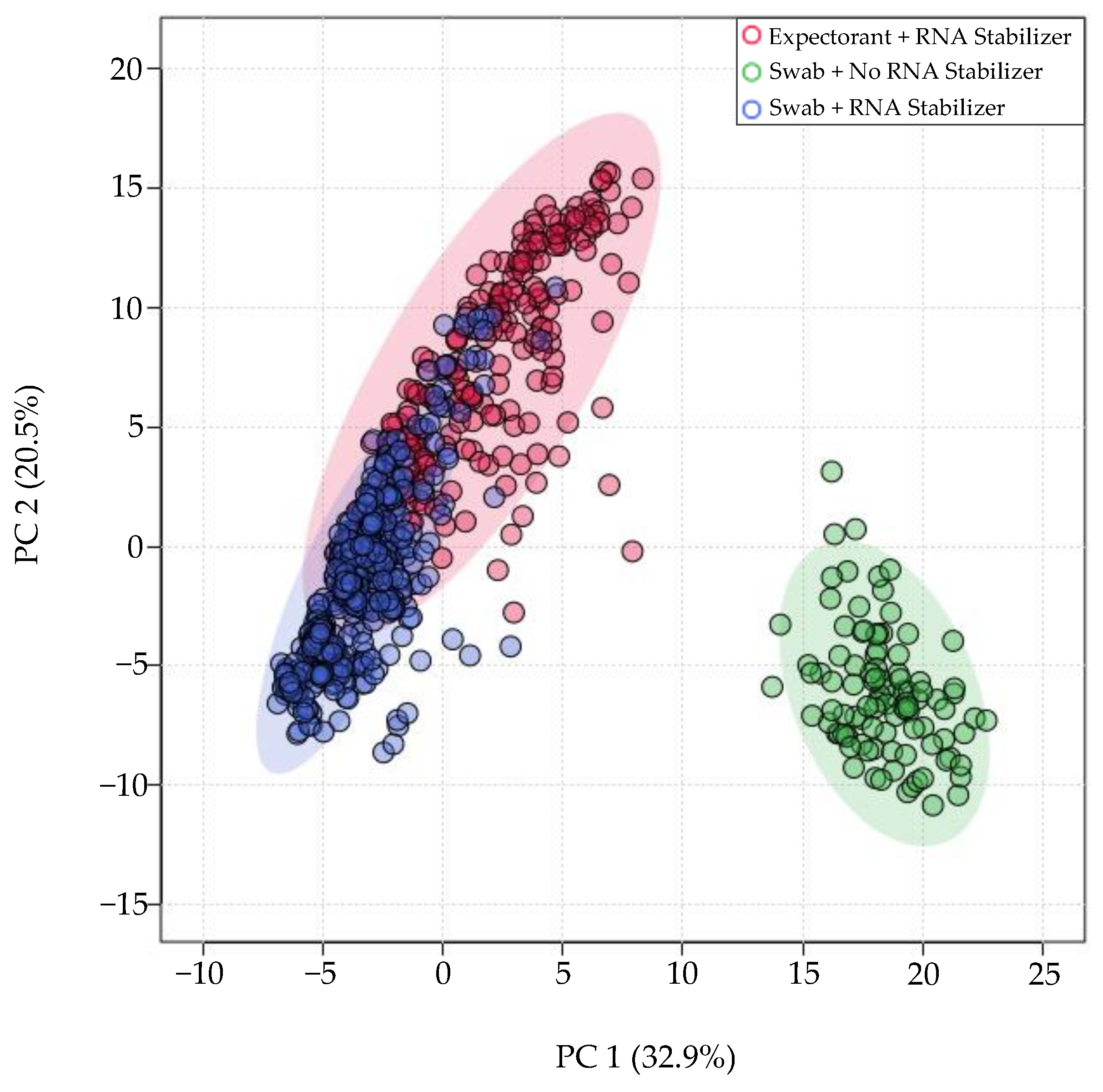
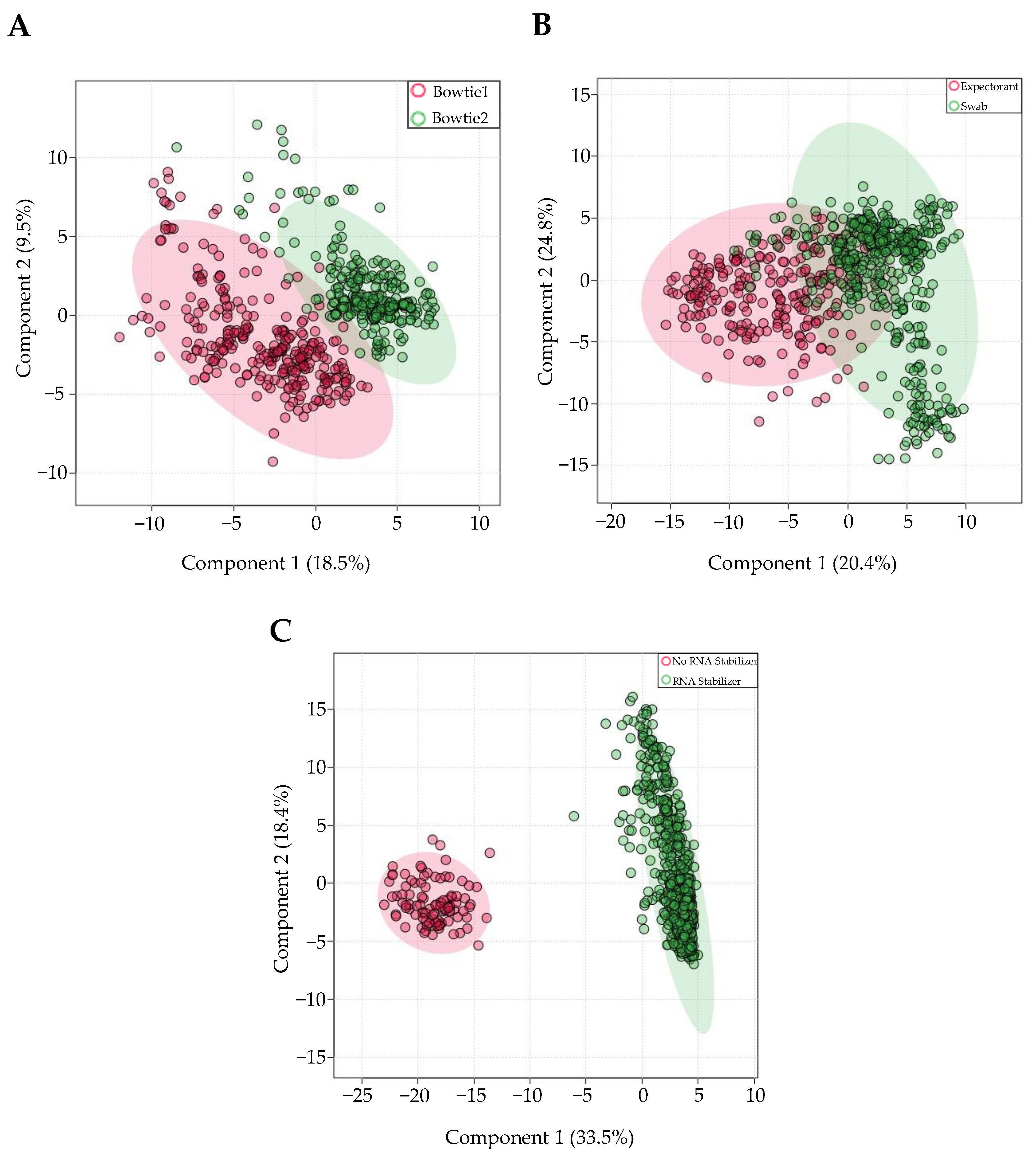
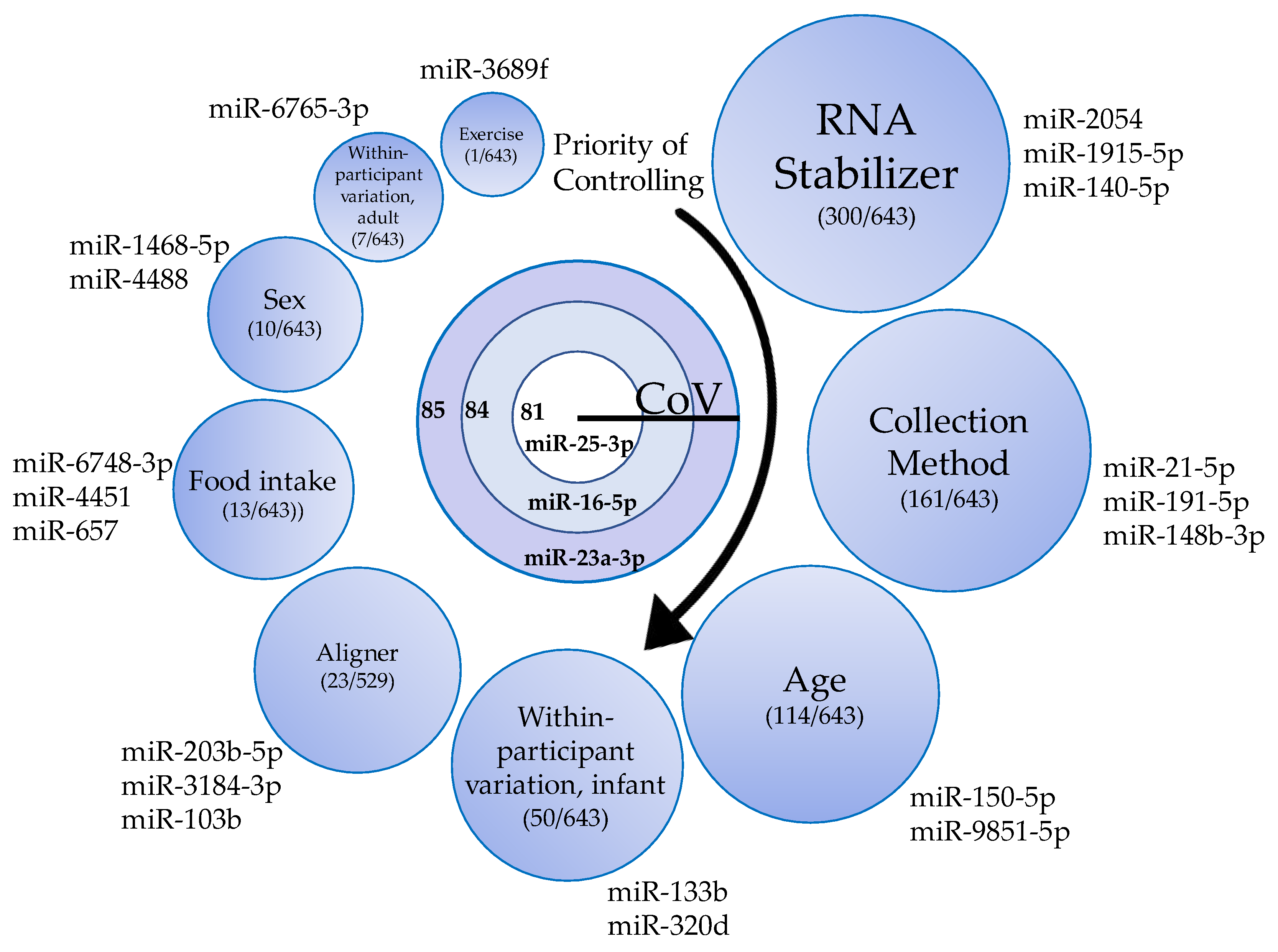
3.3. miRNA Expression Variation as a Result of Technical Factors: Overview
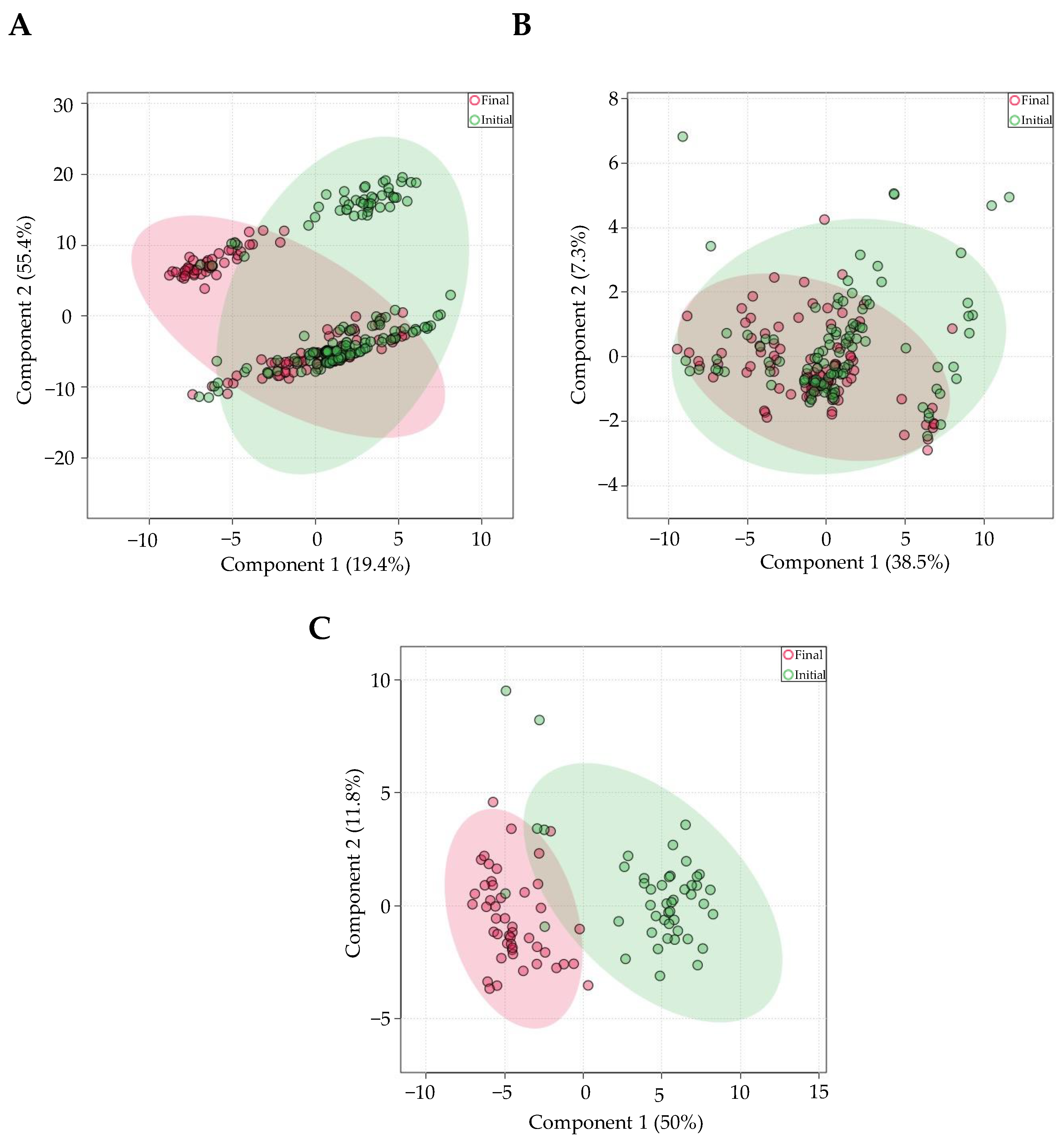
3.4. Individual Effects of Aligner Tool, Saliva Collection Method, and Presence/Absence of RNA Stabilizer on miRNA Expression
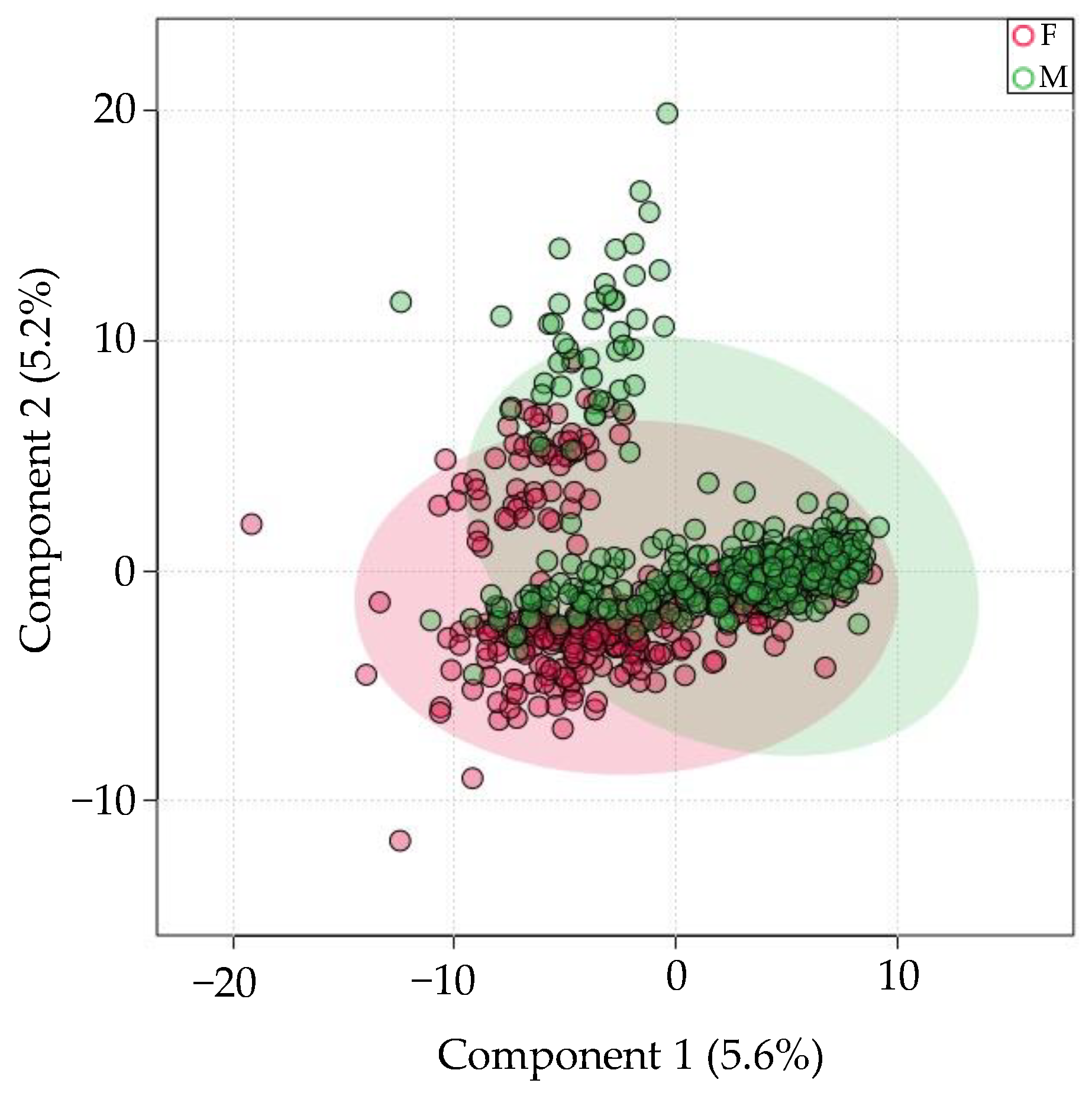
3.5. There Is Minimal Sex-Specific miRNA Expression Variation in Saliva
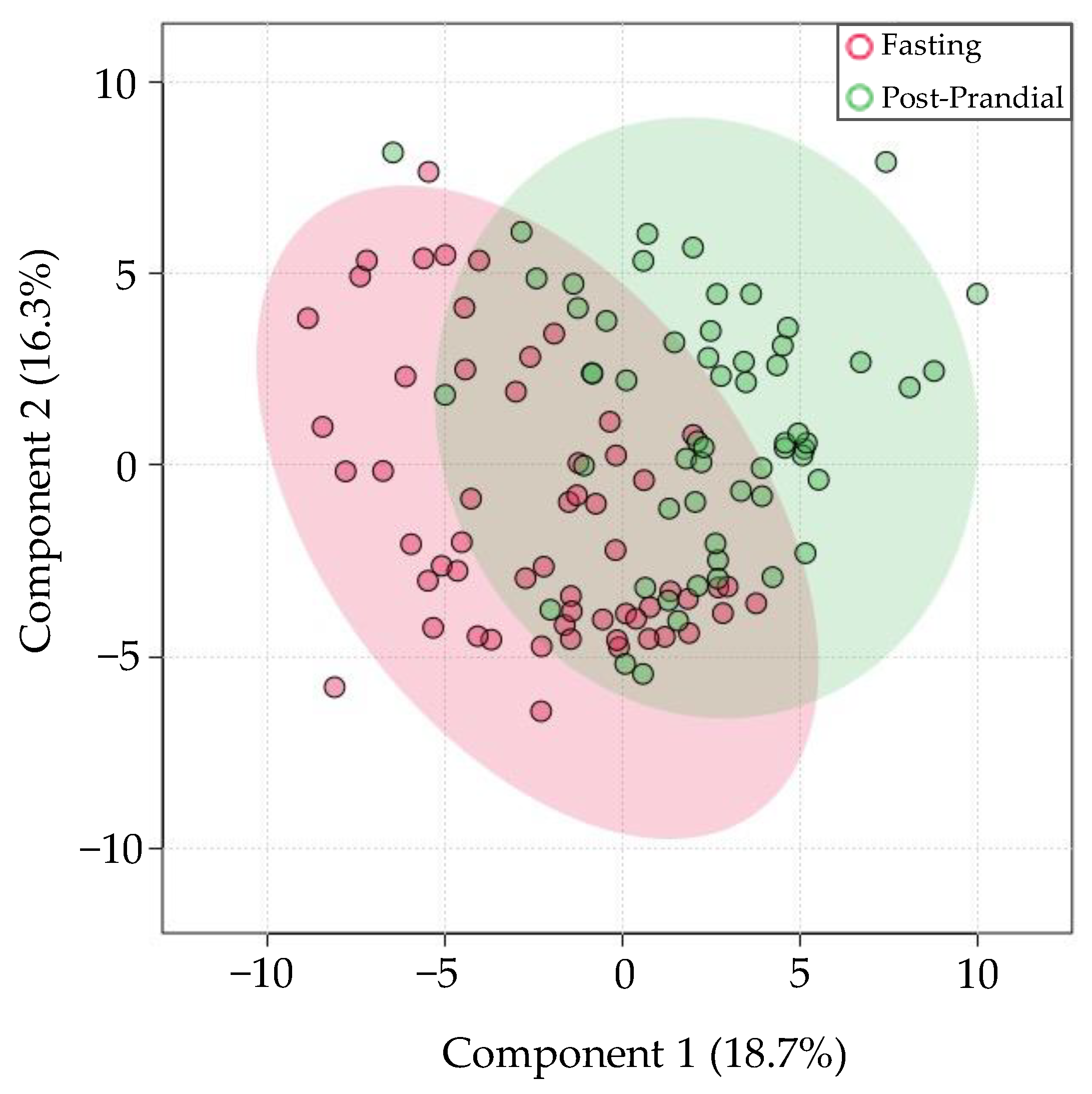
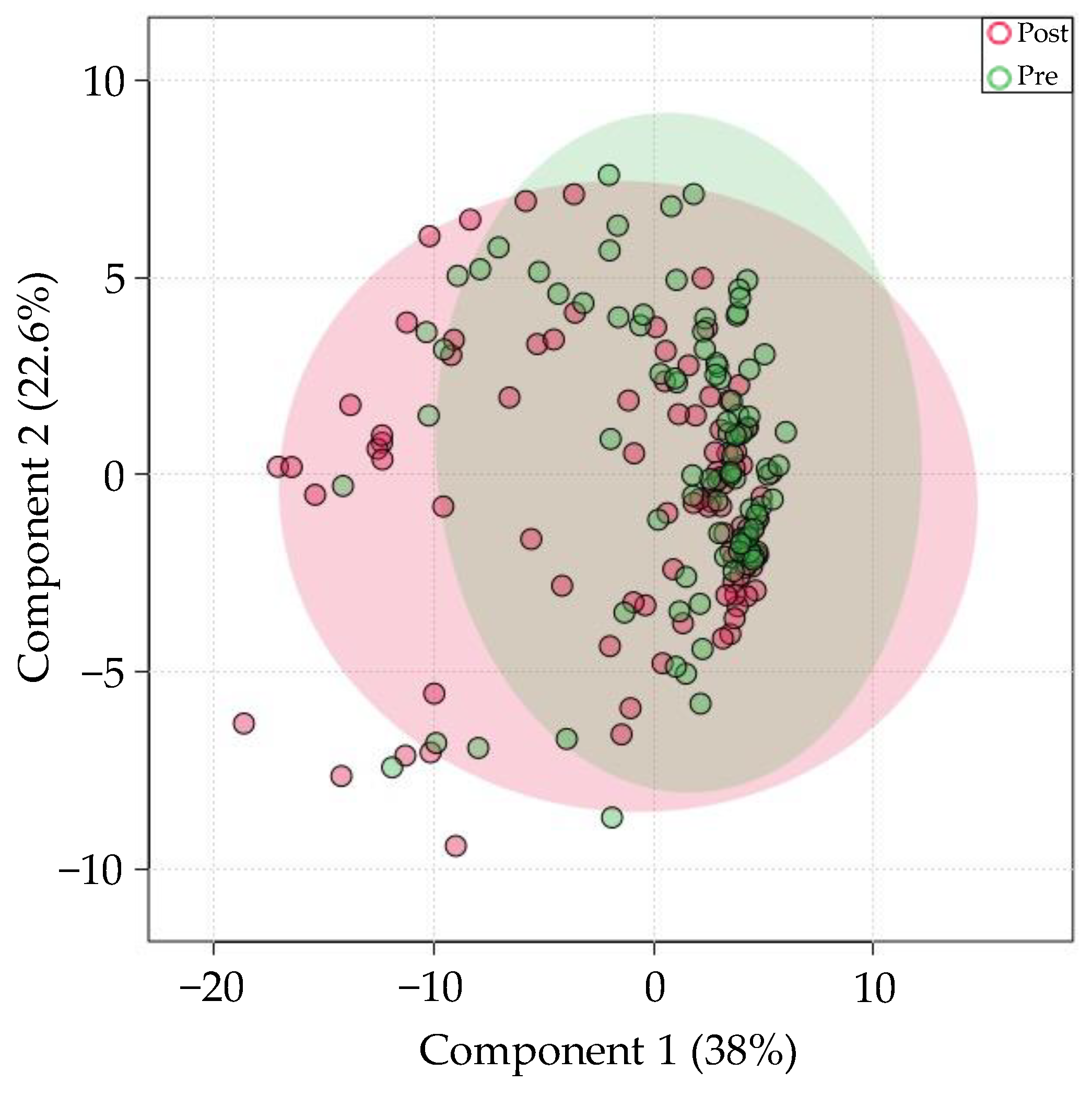
3.6. Differences in Prandial Status at the Time of Saliva Collection Account for Moderate Variation in miRNA Levels
3.7. Exercise has Minimal Impact on Saliva miRNA Profiles
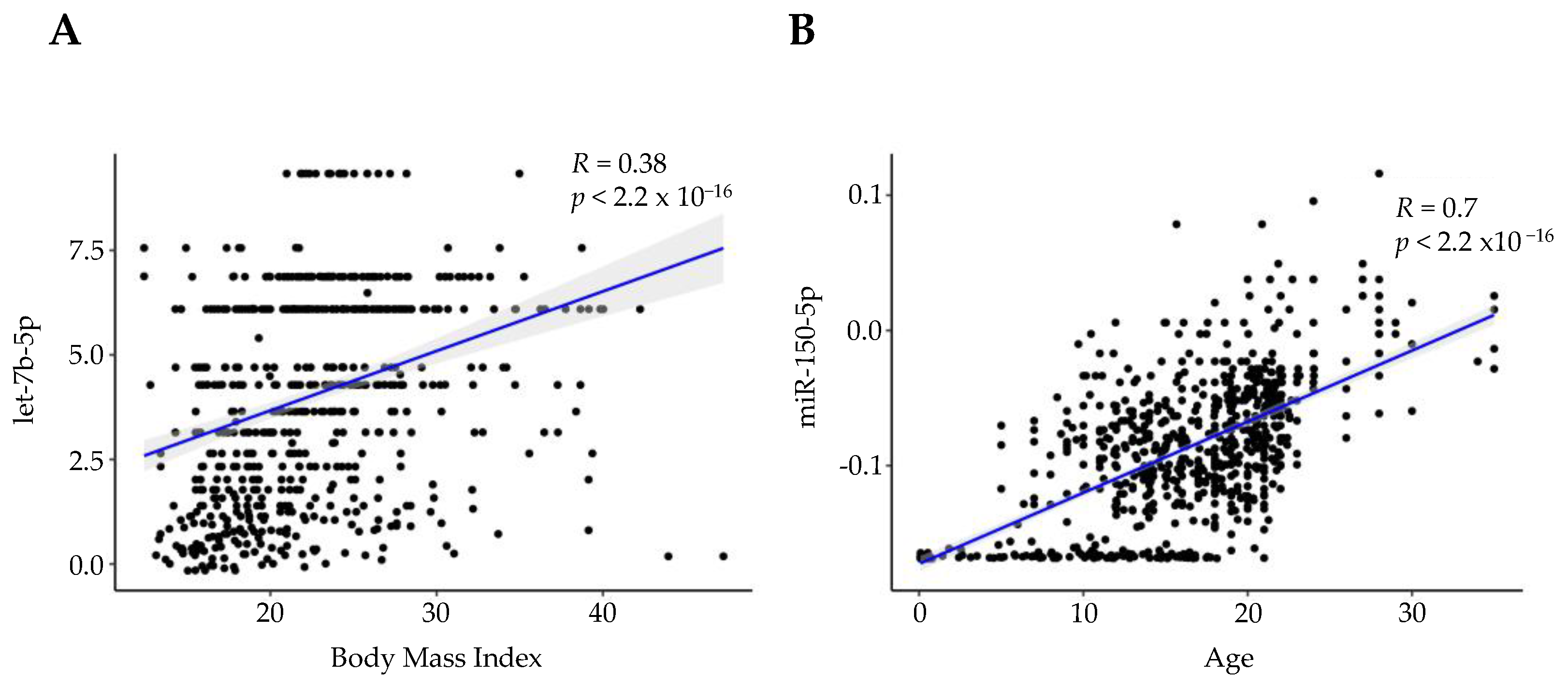
3.8. miRNA Expression Indicates over 75% of miRNAs Consistently Present in Saliva Are Associated with BMI and Age
3.9. Intra-Individual Salivary miRNA Levels Are Relatively Stable over Time for Adults, But Not Infants
3.10. miRNAs as Internal Control Genes
4. Discussion
4.1. Prevalence Analysis
4.2. Aligner
4.3. RNA Stabilizer
4.4. Method of Collection
4.5. Sex-Specific Differences on Salivary miRNA Expression
4.6. Prandial Status Differences on miRNA Expression
4.7. Exercise Differences on miRNA Expression
4.8. miRNAs That Correlate with Age and BMI
4.9. Variation of Intra-Participant Salivary miRNA Expression Is Greater in Infants Than Young Adults
4.10. Endogenous Control miRNAs
4.11. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corey-Bloom, J.; Fischer, R.S.; Kim, A.; Snell, C.; Parkin, G.M.; Granger, D.A.; Granger, S.W.; Thomas, E.A. Levels of Interleukin-6 in Saliva, but Not Plasma, Correlate with Clinical Metrics in Huntington’s Disease Patients and Healthy Control Subjects. Int. J. Mol. Sci. 2020, 21, 6363. [Google Scholar] [CrossRef]
- Han, Y.; Jia, L.; Zheng, Y.; Li, W. Salivary Exosomes: Emerging Roles in Systemic Disease. Int. J. Biol. Sci. 2018, 14, 633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.J.; Loeffert, A.C.; Stokes, J.; Olympia, R.P.; Bramley, H.; Hicks, S.D. Association of Salivary MicroRNA Changes With Prolonged Concussion Symptoms. JAMA Pediatr. 2018, 172, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Beversdorf, D.Q.; Sohl, K.; Levitskiy, D.; Tennant, P.; Goin-Kochel, R.P.; Shaffer, R.C.; Confair, A.; Middleton, F.A.; Hicks, S.D. Saliva RNA Biomarkers of Gastrointestinal Dysfunction in Children With Autism and Neurodevelopmental Disorders: Potential Implications for Precision Medicine. Front. Psychiatry 2022, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.L.; Haffajee, A.D.; Delvin, P.M.; Norris, C.M.; Posner, M.R.; Goodson, J.M. The salivary microbiota as a diagnostic indicator of oral cancer: A descriptive, non-randomized study of cancer-free and oral squamous cell carcinoma subjects. J. Transl. Med. 2005, 3, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, N.; Vining, M.; Bloch-Salisbury, E. Salivary cortisol levels as a biomarker for severity of withdrawal in opioid-exposed newborns. Pediatr. Res. 2019, 87, 1033–1038. [Google Scholar] [CrossRef]
- Hicks, S.D.; Johnson, J.; Carney, M.C.; Bramley, H.; Olympia, R.P.; Loeffert, A.C.; Thomas, N.J. Overlapping MicroRNA Expression in Saliva and Cerebrospinal Fluid Accurately Identifies Pediatric Traumatic Brain Injury. J. Neurotrauma 2018, 35, 64. [Google Scholar] [CrossRef]
- Diesch, T.; Filippi, C.; Fritschi, N.; Filippi, A.; Ritz, N. Cytokines in saliva as biomarkers of oral and systemic oncological or infectious diseases: A systematic review. Cytokine 2021, 143, 155506. [Google Scholar] [CrossRef]
- Wagner, K.E.; McCormick, J.B.; Barns, S.; Carney, M.; Middleton, F.A.; Hicks, S.D. Parent Perspectives Towards Genetic and Epigenetic Testing for Autism Spectrum Disorder. J. Autism Dev. Disord. 2019, 50, 3114–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizawa, J.M.; Schafer, C.A.; Schafer, J.J.; Farrell, J.J.; Paster, B.J.; Wong, D.T.W. Salivary Biomarkers: Toward Future Clinical and Diagnostic Utilities. Clin. Microbiol. Rev. 2013, 26, 781. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.C.; Chan, W.C.; Hu, L.Y.; Lai, C.H.; Hsu, C.N.; Lin, W. Identification of homologous microRNAs in 56 animal genomes. Genomics 2010, 96, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.H.; Shin, S.; Jung, S.R.; Kim, E.; Song, J.J.; Hohng, S. Human Argonaute 2 Has Diverse Reaction Pathways on Target RNAs. Mol. Cell 2015, 59, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicks, S.D.; Carpenter, R.L.; Wagner, K.E.; Pauley, R.; Barros, M.; Tierney-Aves, C.; Barns, S.; Greene, C.D.; Middleton, F.A. Saliva MicroRNA Differentiates Children With Autism From Peers With Typical and Atypical Development. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 296–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehovic, E.; Spahic, L.; Smajlovic-Skenderagic, L.; Pistoljevic, N.; Dzanko, E.; Hajdarpasic, A. Identification of developmental disorders including autism spectrum disorder using salivary miRNAs in children from Bosnia and Herzegovina. PLoS ONE 2020, 15, e0232351. [Google Scholar] [CrossRef]
- Hicks, S.D.; Onks, C.; Kim, R.Y.; Zhen, K.J.; Loeffert, J.; Loeffert, A.C.; Olympia, R.P.; Fedorchak, G.; DeVita, S.; Rangnekar, A.; et al. Diagnosing mild traumatic brain injury using saliva RNA compared to cognitive and balance testing. Clin. Transl. Med. 2020, 10, e197. [Google Scholar] [CrossRef]
- Miller, K.E.; MacDonald, J.P.; Sullivan, L.; Venkata, L.P.R.; Shi, J.; Yeates, K.O.; Chen, S.; Alshaikh, E.; Taylor, H.G.; Hautmann, A.; et al. Salivary miRNA Expression in Children With Persistent Post-concussive Symptoms. Front. Public Health 2022, 10, 1528. [Google Scholar] [CrossRef] [PubMed]
- Hicks, S.D.; Leddy, J.; Lichak, B.P.; Onks, C.; Dretsch, M.; Tennant, P.; Haider, M.N.; Olympia, R.P.; Zuckerman, S.L.; Loeffert, J.; et al. Defining Biological Phenotypes of Mild Traumatic Brain Injury Using Saliva MicroRNA Profiles. J. Neurotrauma 2022, 39, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, V.; O’Halloran, P.; Watson, C.N.; Begum, G.; Acharjee, A.; Yakoub, K.M.; Bentley, C.; Davies, D.J.; Iliceto, P.; Candilera, G.; et al. Unique diagnostic signatures of concussion in the saliva of male athletes: The Study of Concussion in Rugby Union through MicroRNAs (SCRUM). Br. J. Sports Med. 2021, 55, 1395. [Google Scholar] [CrossRef] [PubMed]
- Bendifallah, S.; Suisse, S.; Puchar, A.; Delbos, L.; Poilblanc, M.; Descamps, P.; Golfier, F.; Jornea, L.; Bouteiller, D.; Touboul, C.; et al. Salivary MicroRNA Signature for Diagnosis of Endometriosis. J. Clin. Med. 2022, 11, 612. [Google Scholar] [CrossRef]
- Kang, J.W.; Eun, Y.G.; Lee, Y.C. Diagnostic Value of Salivary miRNA in Head and Neck Squamous Cell Cancer: Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2021, 22, 7026. [Google Scholar] [CrossRef] [PubMed]
- Fadhil, R.S.; Wei, M.Q.; Nikolarakos, D.; Good, D.; Nair, R.G. Salivary microRNA miR-let-7a-5p and miR-3928 could be used as potential diagnostic bio-markers for head and neck squamous cell carcinoma. PLoS ONE 2020, 15, e0221779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapado-González, Ó.; Majem, B.; Muinelo-Romay, L.; álvarez-Castro, A.; Santamaría, A.; Gil-Moreno, A.; López-López, R.; Suárez-Cunqueiro, M.M. Human salivary microRNAs in Cancer. J. Cancer 2018, 9, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, E.A.; Boulghassoul-Pietrzykowska, N.; Wang, Y.; Anees, O.; Kinstlinger, N.S.; Lee, M.; Hamza, S.; Feng, Y.; Pietrzykowski, A.Z. Non-Invasive microRNA Profiling in Saliva can Serve as a Biomarker of Alcohol Exposure and Its Effects in Humans. Front. Genet. 2022, 12, 804222. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.J.; Chen, X.; Tanaka, Y.; Farrer, L.A.; Kranzler, H.R.; Nunez, Y.Z.; Henderson, D.C.; Gelernter, J.; Zhang, H. Salivary microRNAs identified by small RNA sequencing and machine learning as potential biomarkers of alcohol dependence. Epigenomics 2019, 11, 739–749. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, J.; Sun, Y.; Li, F.; Wei, L.; Sun, W.; Deng, J.; Yuan, Y.; Wang, Z. Profiling of Differentially Expressed MicroRNAs in Saliva of Parkinson’s Disease Patients. Front. Neurol. 2021, 12, 738530. [Google Scholar] [CrossRef] [PubMed]
- Cressatti, M.; Juwara, L.; Galindez, J.M.; Velly, A.M.; Nkurunziza, E.S.; Marier, S.; Canie, O.; Gornistky, M.; Schipper, H.M. Salivary microR-153 and microR-223 Levels as Potential Diagnostic Biomarkers of Idiopathic Parkinson’s Disease. Mov. Disord. 2020, 35, 468–477. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, L.; Qiao, X.; Zhang, X.; Dong, S.F.; Wu, M.T.; Zhai, K.; Shi, H.Z. Salivary microRNAs show potential as biomarkers for early diagnosis of malignant pleural effusion. Transl. Lung Cancer Res. 2020, 9, 1247. [Google Scholar] [CrossRef] [PubMed]
- Alsop, E.; Meechoovet, B.; Kitchen, R.; Sweeney, T.; Beach, T.G.; Serrano, G.E.; Hutchins, E.; Ghiran, I.; Reiman, R.; Syring, M.; et al. A Novel Tissue Atlas and Online Tool for the Interrogation of Small RNA Expression in Human Tissues and Biofluids. Front. Cell Dev. Biol. 2022, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Yeri, A.; Courtright, A.; Reiman, R.; Carlson, E.; Beecroft, T.; Janss, A.; Siniard, A.; Richholt, R.; Balak, C.; Rozowsky, J.; et al. Total Extracellular Small RNA Profiles from Plasma, Saliva, and Urine of Healthy Subjects. Sci. Rep. 2017, 7, srep44061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, R.; Heavey, S.; Graham, D.G.; Wellman, R.; Khan, S.; Thrumurthy, S.; Simpson, B.S.; Baker, T.; Jevons, S.; Ariza, J.; et al. An optimised saliva collection method to produce high-yield, high-quality RNA for translational research. PLoS ONE 2020, 15, e0229791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, J.; Woldhuis, R.R.; Boudewijn, I.M.; van den Berg, A.; Kluiver, J.; Kok, K.; Terpstra, M.M.; Guryev, V.; de Vries, M.; Vermeulen, C.J.; et al. Age-related gene and miRNA expression changes in airways of healthy individuals. Sci. Rep. 2019, 9, 3765. [Google Scholar] [CrossRef] [Green Version]
- Noren Hooten, N.; Fitzpatrick, M.; Wood, W.H.; De, S.; Ejiogu, N.; Zhang, Y.; Mattison, J.A.; Becker, K.G.; Zonderman, A.B.; Evans, M.K. Age-related changes in microRNA levels in serum. Aging (Albany NY) 2013, 5, 725. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Eghbali, M. Influence of sex differences on microRNA gene regulation in disease. Biol. Sex Differ. 2014, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Karere, G.M.; Cox, L.A.; Bishop, A.C.; South, A.M.; Shaltout, H.A.; Mercado-Deane, M.G.; Cuda, S. Sex Differences in MicroRNA Expression and Cardiometabolic Risk Factors in Hispanic Adolescents with Obesity. J. Pediatr. 2021, 235, 138–143.e5. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, Q.; Ma, X.; Wang, J.; Liang, T. miRNA and mRNA expression analysis reveals potential sex-biased miRNA expression. Sci. Rep. 2017, 7, 39812. [Google Scholar] [CrossRef] [Green Version]
- Mantilla-Escalante, D.C.; de las Hazas, M.C.L.; Gil-Zamorano, J.; del Pozo-Acebo, L.; Crespo, M.C.; Martín-Hernández, R.; del Saz, A.; Tomé-Carneiro, J.; Cardona, F.; Cornejo-Pareja, I.; et al. Postprandial Circulating miRNAs in Response to a Dietary Fat Challenge. Nutrients 2019, 11, 1326. [Google Scholar] [CrossRef] [Green Version]
- Kogure, A.; Uno, M.; Ikeda, T.; Nishida, E. The microRNA machinery regulates fasting-induced changes in gene expression and longevity in Caenorhabditis elegans. J. Biol. Chem. 2017, 292, 11300. [Google Scholar] [CrossRef]
- Scipioni, A.M.; Ornstein, R.M.; Hicks, S.D.; Zhan, X. Differential Expression Of Salivary Microrna In Anorexia Nervosa And Anxiety Disorders. J. Adolesc. Health 2019, 64, S13. [Google Scholar] [CrossRef]
- Hicks, S.D.; Jacob, P.; Perez, O.; Baffuto, M.; Gagnon, Z.; Middleton, F.A. The Transcriptional Signature of a Runner’s High. Med. Sci. Sports Exerc. 2019, 51, 970–978. [Google Scholar] [CrossRef] [PubMed]
- LaRocca, D.; Barns, S.; Hicks, S.D.; Brindle, A.; Williams, J.; Uhlig, R.; Johnson, P.; Neville, C.; Middleton, F.A. Comparison of serum and saliva miRNAs for identification and characterization of mTBI in adult mixed martial arts fighters. PLoS ONE 2019, 14, e0207785. [Google Scholar] [CrossRef] [Green Version]
- Telles, G.D.; Libardi, C.A.; Conceição, M.S.; Vechin, F.C.; Lixandrão, M.E.; De Andrade, A.L.L.; Guedes, D.N.; Ugrinowitsch, C.; Camera, D.M. Time Course of Skeletal Muscle miRNA Expression after Resistance, High-Intensity Interval, and Concurrent Exercise. Med. Sci. Sports Exerc. 2021, 53, 1708–1718. [Google Scholar] [CrossRef]
- Zhou, Q.; Shi, C.; Lv, Y.; Zhao, C.; Jiao, Z.; Wang, T. Circulating microRNAs in Response to Exercise Training in Healthy Adults. Front. Genet. 2020, 11, 256. [Google Scholar] [CrossRef] [Green Version]
- Iacomino, G.; Siani, A. Role of microRNAs in obesity and obesity-related diseases. Genes Nutr. 2017, 12, 23. [Google Scholar] [CrossRef] [Green Version]
- Shirtcliff, E.A.; Granger, D.A.; Schwartz, E.; Curran, M.J. Use of salivary biomarkers in biobehavioral research: Cotton-based sample collection methods can interfere with salivary immunoassay results. Psychoneuroendocrinology 2001, 26, 165–173. [Google Scholar] [CrossRef]
- Golatowski, C.; Gesell Salazar, M.; Dhople, V.M.; Hammer, E.; Kocher, T.; Jehmlich, N.; Völker, U. Comparative evaluation of saliva collection methods for proteome analysis. Clin. Chim. Acta 2013, 419, 42–46. [Google Scholar] [CrossRef]
- Henson, B.S.; Wong, D.T. Collection, storage, and processing of saliva samples for downstream molecular applications. Methods Mol. Biol. 2010, 666, 21–30. [Google Scholar]
- Ziemann, M.; Kaspi, A.; El-Osta, A. Evaluation of microRNA alignment techniques. RNA 2016, 22, 1120–1138. [Google Scholar] [CrossRef] [Green Version]
- Tam, S.; Tsao, M.S.; McPherson, J.D. Optimization of miRNA-seq data preprocessing. Brief. Bioinform. 2015, 16, 950. [Google Scholar] [CrossRef] [PubMed]
- Romani, C.; Salviato, E.; Paderno, A.; Zanotti, L.; Ravaggi, A.; Deganello, A.; Berretti, G.; Gualtieri, T.; Marchini, S.; D’Incalci, M.; et al. Genome-wide study of salivary miRNAs identifies miR-423-5p as promising diagnostic and prognostic biomarker in oral squamous cell carcinoma. Theranostics 2021, 11, 2987. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.E.; Buffa, F.M.; Camps, C.; Ramachandran, A.; Leek, R.; Taylor, M.; Patil, M.; Sheldon, H.; Betts, G.; Homer, J.; et al. The small-nucleolar RNAs commonly used for microRNA normalisation correlate with tumour pathology and prognosis. Br. J. Cancer 2011, 104, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masè, M.; Grasso, M.; Avogaro, L.; D’Amato, E.; Tessarolo, F.; Graffigna, A.; Denti, M.A.; Ravelli, F. Selection of reference genes is critical for miRNA expression analysis in human cardiac tissue. A focus on atrial fibrillation. Sci. Rep. 2017, 7, srep41127. [Google Scholar] [CrossRef]
- Ostheim, P.; Tichý, A.; Sirak, I.; Davidkova, M.; Stastna, M.M.; Kultova, G.; Paunesku, T.; Woloschak, G.; Majewski, M.; Port, M.; et al. Overcoming challenges in human saliva gene expression measurements. Sci. Rep. 2020, 10, 11147. [Google Scholar] [CrossRef]
- Hicks, S.D.; Khurana, N.; Williams, J.; Greene, C.D.; Uhlig, R.; Middleton, F.A. Diurnal oscillations in human salivary microRNA and microbial transcription: Implications for human health and disease. PLoS ONE 2018, 13, e0198288. [Google Scholar] [CrossRef]
- Hicks, S.D.; Onks, C.; Kim, R.Y.; Zhen, K.J.; Loeffert, J.; Loeffert, A.C.; Olympia, R.P.; Fedorchak, G.; DeVita, S.; Gagnon, Z.; et al. Refinement of saliva microRNA biomarkers for sports-related concussion. J. Sport Health Sci. 2021. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, J.; Liao, Y.; Huang, L.; Wen, C.; Lin, M.; Li, W.; Zhu, Y.; Wu, X.; Iwamoto, A.; et al. MiR-27b-3p promotes migration and invasion in colorectal cancer cells by targeting HOXA10. Biosci. Rep. 2019, 39, BSR20191087. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, G.; Zhang, B.; Liu, C.; Yu, Y.; Jin, Y. miR-27b-3p suppresses cell proliferation, migration and invasion by targeting LIMK1 in colorectal cancer. Int. J. Clin. Exp. Pathol. 2017, 10, 9251. [Google Scholar]
- Han, M.; Li, N.; Li, F.; Wang, H.; Ma, L. MiR-27b-3p exerts tumor suppressor effects in esophageal squamous cell carcinoma by targeting Nrf2. Hum. Cell 2020, 33, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Hummel, R.; Sie, C.; Watson, D.I.; Wang, T.; Ansar, A.; Michael, M.Z.; Van Der Hoek, M.; Haier, J.; Hussey, D.J. MicroRNA signatures in chemotherapy resistant esophageal cancer cell lines. World J. Gastroenterol. 2014, 20, 14904–14912. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, Z.; Zhou, J.; Liu, Z. circRNA RNF111 regulates the growth, migration and invasion of gastric cancer cells by binding to miR-27b-3p. Int. J. Mol. Med. 2020, 46, 1873. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Zhi, X.; Zhang, X.; Fu, M.; Huang, H.; Fan, Y.; Guan, W.; Zou, C. miR-27b-3p suppresses cell proliferation through targeting receptor tyrosine kinase like orphan receptor 1 in gastric cancer. J. Exp. Clin. Cancer Res. 2015, 34, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Hu, J.; Yu, T.; You, S.; Zhang, Y.; Hu, L. miR-27b-3p/MARCH7 regulates invasion and metastasis of endometrial cancer cells through Snail-mediated pathway. Acta Biochim. Biophys. Sin. (Shanghai) 2019, 51, 492–500. [Google Scholar] [CrossRef]
- Kim, M.K.; Lee, S.K.; Park, J.H.; Lee, J.H.; Yun, B.H.; Park, J.H.; Seo, S.K.; Cho, S.; Choi, Y.S. Ginsenoside Rg3 Decreases Fibrotic and Invasive Nature of Endometriosis by Modulating miRNA-27b: In Vitro and In Vivo Studies. Sci. Rep. 2017, 7, 17670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Si, W.; Shen, J.; Du, C.; Lou, W.; Bao, C.; Zheng, H.; Pan, J.; Zhong, G.; Xu, L.; et al. miR-27b-3p inhibits proliferation and potentially reverses multi-chemoresistance by targeting CBLB/GRB2 in breast cancer cells. Cell Death Dis. 2018, 9, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, X.; Ge, D.; Shen, D.; Chen, X.; Wang, X.; Zhang, L.; Jia, C.; Zeng, J.; He, Y.; Qiu, H.; et al. miR-27b Suppresses Endothelial Cell Proliferation and Migration by Targeting Smad7 in Kawasaki Disease. Cell. Physiol. Biochem. 2018, 48, 1804–1814. [Google Scholar] [CrossRef]
- Puppo, M.; Bucci, G.; Rossi, M.; Giovarelli, M.; Bordo, D.; Moshiri, A.; Gorlero, F.; Gherzi, R.; Briata, P. miRNA-Mediated KHSRP Silencing Rewires Distinct Post-transcriptional Programs during TGF-β-Induced Epithelial-to-Mesenchymal Transition. Cell Rep. 2016, 16, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Dawes, C. Estimates, from salivary analyses, of the turnover time of the oral mucosal epithelium in humans and the number of bacteria in an edentulous mouth. Arch. Oral Biol. 2003, 48, 329–336. [Google Scholar] [CrossRef]
- Ferragina, P.; Manzini, G. Opportunistic data structures with applications. In Proceedings of the 41st Annual Symposium on Foundations of Computer Science, Redondo Beach, CA, USA, 12–14 November 2000; pp. 390–398. [Google Scholar]
- Manzini, G. The Burrows-Wheeler Transform: Theory and Practice. In International Symposium on Mathematical Foundations of Computer Science; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef]
- Jadhav, K.B.; Shah, V.; Mahajan, A.; Malhotra, P.; Parmar, G.; Shah, J. Salivary and tumour tissue miR-21 for prediction of cervical lymph node metastasis in patients with oral squamous cell carcinoma: An observational study. Lancet. Oncol. 2022, 23, S26. [Google Scholar] [CrossRef]
- Khalifa, O.; Pers, Y.M.; Ferreira, R.; Sénéchal, A.; Jorgensen, C.; Apparailly, F.; Duroux-Richard, I. X-Linked miRNAs Associated with Gender Differences in Rheumatoid Arthritis. Int. J. Mol. Sci. 2016, 17, 1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Zhi, T.; Xu, W.; Xu, X.; Wu, W.; Yu, T.; Nie, E.; Zhou, X.; Bao, Z.; Jin, X.; et al. MicroRNA-1468-5p inhibits glioma cell proliferation and induces cell cycle arrest by targeting RRM1. Am. J. Cancer Res. 2017, 7, 784. [Google Scholar] [PubMed]
- McLeod, J.; Elford, H.; Ryan, J. The ribonucleotide reductase inhibitors didox and trimidox suppress IgE-stimulated mast cell cytokine secretion (HYP3P.350). J. Immunol. 2014, 192, 54. [Google Scholar]
- Guo, X.; Su, B.; Zhou, Z.; Sha, J. Rapid evolution of mammalian X-linked testis microRNAs. BMC Genom. 2009, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Bu, L.; Salto, L.M.; De Leon, K.J.; De Leon, M. Polymorphisms in fatty acid binding protein 5 show association with type 2 diabetes. Diabetes Res. Clin. Pract. 2011, 92, 82–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, K.; Guo, Y.; Zhang, Y.; Wang, K.; Jia, Y.; Sun, S. Allele-specific targeting of hsa-miR-657 to human IGF2R creates a potential mechanism underlying the association of ACAA-insertion/deletion polymorphism with type 2 diabetes. Biochem. Biophys. Res. Commun. 2008, 374, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Z.; Liu, G.; Jin, C.; Zhang, Q.; Man, S.; Wang, Z. miR-657 Promotes Macrophage Polarization toward M1 by Targeting FAM46C in Gestational Diabetes Mellitus. Mediators Inflamm. 2019, 2019, 4851214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Wang, H.; Li, C.; Zhang, X.; Xiu, X.; Teng, P.; Wang, Z. Dysregulation of microRNA-657 influences inflammatory response via targeting interleukin-37 in gestational diabetes mellitus. J. Cell. Physiol. 2019, 234, 7141–7148. [Google Scholar] [CrossRef]
- Buj-Bello, A.; Laugel, V.; Messaddeq, N.; Zahreddine, H.; Laporte, J.; Pellissier, J.F.; Mandel, J.L. The lipid phosphatase myotubularin is essential for skeletal muscle maintenance but not for myogenesis in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 15060–15065. [Google Scholar] [CrossRef] [Green Version]
- Kuang, S.Q.; Kwartler, C.S.; Byanova, K.L.; Pham, J.; Gong, L.; Prakash, S.K.; Huang, J.; Kamm, K.E.; Stull, J.T.; Sweeney, H.L.; et al. Rare, nonsynonymous variant in the smooth muscle-specific isoform of myosin heavy chain, MYH11, R247C, alters force generation in the aorta and phenotype of smooth muscle cells. Circ. Res. 2012, 110, 1411–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Tang, D.; Wang, W.; Yang, Y.; Wu, X.; Wang, L.; Wang, D. circlmtk2 acts as a sponge of miR-150-5p and promotes proliferation and metastasis in gastric cancer. Mol. Cancer 2019, 18, 162. [Google Scholar] [CrossRef]
- Liu, F.; Di Wang, X. miR-150-5p represses TP53 tumor suppressor gene to promote proliferation of colon adenocarcinoma. Sci. Rep. 2019, 9, 6740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.B.; Moolani, H.V.; Sene, A.; Sidhu, R.; Kell, P.; Lin, J.B.; Dong, Z.; Ban, N.; Ory, D.S.; Apte, R.S. Macrophage microRNA-150 promotes pathological angiogenesis as seen in age-related macular degeneration. JCI Insight 2018, 3, e120157. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Danielson, K.M.; Benton, M.C.; Ziegler, O.; Shah, R.; Stubbs, R.S.; Das, S.; Macartney-Coxson, D. miRNA signatures of insulin resistance in obesity. Obesity (Silver Spring) 2017, 25, 1734. [Google Scholar] [CrossRef] [Green Version]
- Koutsoulidou, A.; Mastroyiannopoulos, N.P.; Furling, D.; Uney, J.B.; Phylactou, L.A. Expression of miR-1, miR-133a, miR-133b and miR-206 increases during development of human skeletal muscle. BMC Dev. Biol. 2011, 11, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mytidou, C.; Koutsoulidou, A.; Zachariou, M.; Prokopi, M.; Kapnisis, K.; Spyrou, G.M.; Anayiotos, A.; Phylactou, L.A. Age-Related Exosomal and Endogenous Expression Patterns of miR-1, miR-133a, miR-133b, and miR-206 in Skeletal Muscles. Front. Physiol. 2021, 12, 708278. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall (n = 922) | |
|---|---|
| Sex | |
| Female | 435 (47.0%) |
| Male | 473 (51.1%) |
| Unknown | 18 (1.9%) |
| Age (years) | |
| Mean (SD) | 13 (±8.5) 1 |
| Race | |
| Asian | 21 (2.3%) |
| Biracial | 36 (3.9%) |
| Black | 67 (7.2%) |
| Hispanic | 17 (1.8%) |
| White | 633 (68.4%) |
| Other | 33 (3.6%) |
| Unknown | 119 (12.9%) |
| BMI | |
| Mean (SD) | 23 (±5.3) 2 |
| Overall (n = 1225) | |
|---|---|
| RNA Stabilizer | |
| Presence | 1125 (91.8%) |
| Absence | 100 (8.2%) |
| Prandial Status | |
| Fasting | 58 (4.7%) |
| Post-Prandial | 58 (4.7%) |
| Aligner Tool | |
| Bowtie1 | 1225 (100%) |
| Bowtie2 | 381 (31.1%) |
| Collection Method | |
| Swab | 991 (80.9%) |
| Expectorant | 234 (19.1%) |
| Exercise Status | |
| Pre-Exercise | 93 (7.6%) |
| Post-Exercise | 93 (7.6%) |
| miRNAs | Prevalence | Sum | Median | Variation Coefficient | Percentage of Counts |
|---|---|---|---|---|---|
| miR-27b-3p | 1169 | 40,926,803 | 26,415 | 94.157 | 10.479 |
| miR-26a-5p | 1166 | 36,244,335 | 21,472 | 102.632 | 9.280 |
| miR-27a-3p | 1154 | 30,386,731 | 13,461 | 119.282 | 7.781 |
| let-7a-5p | 1132 | 24,298,712 | 12,728 | 117.735 | 6.222 |
| let-7c-5p | 1122 | 20,944,858 | 8860 | 133.474 | 5.363 |
| miR-203a-3p | 1162 | 20,639,200 | 8841 | 146.905 | 5.285 |
| let-7b-5p | 1137 | 19,630,177 | 6211 | 143.746 | 5.026 |
| miR-200a-3p | 1108 | 12,704,002 | 5658 | 129.103 | 3.253 |
| miR-140-5p | 855 | 12,386,415 | 5632 | 134.285 | 3.172 |
| miR-2054 | 843 | 11,951,959 | 4976 | 138.597 | 3.060 |
| miR-375-3p | 1161 | 9,732,576 | 6071 | 99.767 | 2.492 |
| miR-203b-5p | 1159 | 9,488,769 | 5195 | 125.452 | 2.430 |
| miR-223-3p | 1103 | 7,058,158 | 3209 | 124.497 | 1.807 |
| miR-205-5p | 332 | 6,941,928 | 0 | 251.122 | 1.777 |
| miR-16-5p | 1149 | 6,791,136 | 4757 | 84.338 | 1.739 |
| let-7f-5p | 1135 | 6,756,344 | 3727 | 109.492 | 1.730 |
| miR-141-3p | 873 | 6,330,255 | 30 | 253.150 | 1.621 |
| miR-22-3p | 1162 | 6,075,567 | 3065 | 117.644 | 1.556 |
| miR-147b-5p | 897 | 4,877,797 | 2960 | 114.340 | 1.249 |
| miR-23b-3p | 1143 | 3,973,076 | 2566 | 91.646 | 1.017 |
| miR-23a-3p | 1155 | 3,833,282 | 2605 | 84.696 | 0.982 |
| miR-21-5p | 544 | 3,783,555 | 8 | 309.879 | 0.969 |
| miR-92a-1-5p | 842 | 3,557,304 | 2078 | 113.313 | 0.911 |
| miR-24-3p | 1097 | 3,393,089 | 2125 | 96.327 | 0.869 |
| let-7g-5p | 1108 | 3,358,980 | 2056 | 96.653 | 0.860 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, R.; Montgomery, A.; Scipioni, A.; Jhaveri, P.; Schmidt, A.T.; Hicks, S.D. Confounding Factors Impacting microRNA Expression in Human Saliva: Methodological and Biological Considerations. Genes 2022, 13, 1874. https://doi.org/10.3390/genes13101874
Sullivan R, Montgomery A, Scipioni A, Jhaveri P, Schmidt AT, Hicks SD. Confounding Factors Impacting microRNA Expression in Human Saliva: Methodological and Biological Considerations. Genes. 2022; 13(10):1874. https://doi.org/10.3390/genes13101874
Chicago/Turabian StyleSullivan, Rhea, Austin Montgomery, Anna Scipioni, Pooja Jhaveri, Adam T. Schmidt, and Steven D. Hicks. 2022. "Confounding Factors Impacting microRNA Expression in Human Saliva: Methodological and Biological Considerations" Genes 13, no. 10: 1874. https://doi.org/10.3390/genes13101874
APA StyleSullivan, R., Montgomery, A., Scipioni, A., Jhaveri, P., Schmidt, A. T., & Hicks, S. D. (2022). Confounding Factors Impacting microRNA Expression in Human Saliva: Methodological and Biological Considerations. Genes, 13(10), 1874. https://doi.org/10.3390/genes13101874