

Plastid Phylogenomics of Paeonia and the Evolution of Ten Flower Types in Tree Peony



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species of Paeoniaceae and Cultivars Represented by 10 Flower Types
2.2. Sampling, DNA Extraction and Sequencing
2.3. Assembly and Annotation of Chloroplast Genomes
2.4. Structure, Comparison and Phylogenetic Analysis of Chloroplast Genomes
2.5. Reconstruction of Ancestral Characteristics for Flower Architecture
3. Results
3.1. Basic Characteristics of the Chloroplast Genome of 10 Tree Peony Flower Types
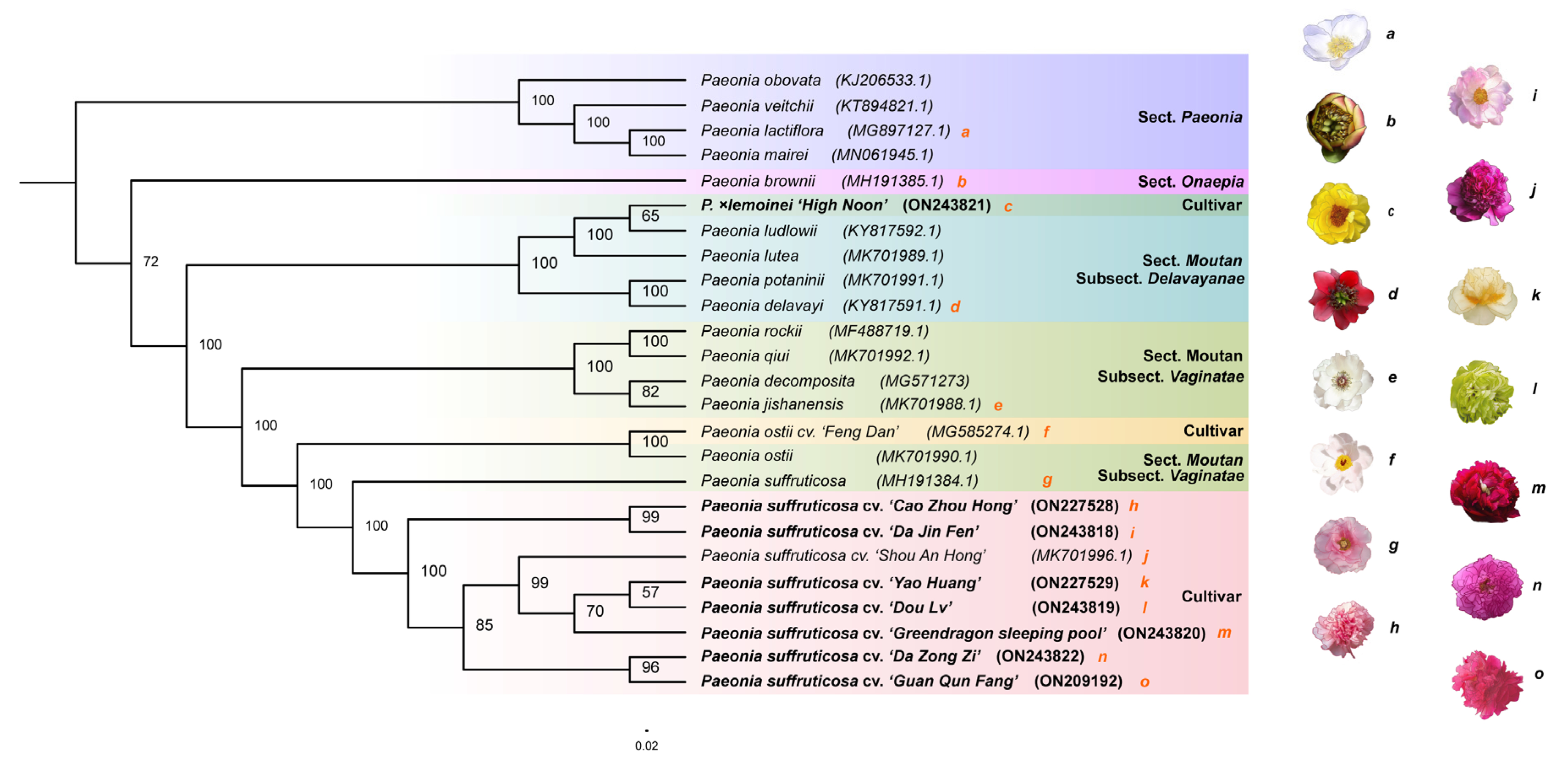
3.2. Phylogenetic Analysis of the Chloroplast Genomes of 15 Paeoniaceae Species and 10 Tree Peony Flower Types
3.3. Reconstruction of Evolutionary Trajectory of Flower Characteristics of Paeoniaceae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrews, H.C. Paeonia suffruticosa. Bot. Repos. 1804, 6, 373. [Google Scholar]
- Worsdell, W.C. A study of the vascular system in certain orders of the Ranales. Ann. Bot 1908, 22, 663. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, M.; Li, S.; Chen, Q.; Teixeira da Silva, J.A.; Wang, A.; Yu, X.; Wang, L. Germplasm resources and genetic breeding of Paeonia: A systematic review. Hortic. Res. 2020, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Stern, F.C. A Study of the Genus Paeonia. R. Hortic. Soc. Lond. 1946, 1, 155. [Google Scholar]
- Lynch, R.I. A new classification of the genus Paeonia. J. R. Hortic. Soc. 1890, 12, 428–445. [Google Scholar]
- Hong, D.Y.; Pan, K.Y. A revision of the Paeonia suffruticosa complex (Paeoniaceae). Nord. J. Bot. 1999, 19, 289–300. [Google Scholar] [CrossRef]
- Hong, D.Y.; Pan, K.Y. Taxonomical history and revision of Paeonia sect. Moutan (Paeoniaceae). Acta Phytotaxon. Sin. 1999, 37, 351–368. [Google Scholar]
- Hong, D.Y.; Zhou, S.L.; He, X.J.; Yuan, J.H.; Zhang, Y.L.; Cheng, F.Y.; Zeng, X.L.; Wang, Y.; Zhang, X.X. Current status of wild tree peony species with special reference to conservation. Biodivers. Sci. 2017, 25, 781–793. [Google Scholar] [CrossRef]
- Li, Z.; Yang, K.; Zhao, Y.; Lin, P. Genetic diversity analysis based on sequence-related amplified polymorphism (SRAP) reveals the genetic background of Xinan tree peony group cultivars from different geographical provenances. Hortic. Environ. Biotechnol. 2018, 59, 567–574. [Google Scholar] [CrossRef]
- Wang, L.; Shiraishi, A.; Hashimoto, F.; Aoki, N.; Shimizu, K.; Sakata, Y. Analysis of Petal Anthocyanins to Investigate Flower Coloration of Zhongyuan (Chinese) and Daikon Island (Japanese) Tree Peony Cultivars. J. Plant Res. 2001, 114, 33–43. [Google Scholar] [CrossRef]
- America Peony Society (APS). Peony Registry. Available online: https://americanpeonysociety.org (accessed on 18 May 2022).
- Hong, D.Y. Notes on taxonomy of Paeonia sect. Moutan DC. (Paeoniaceae). Acta Phytotaxon. Sin. 2005, 43, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Pan, K.Y. Paeonia. Flora of China. Beijing Sci. Press 1979, 27, 37. [Google Scholar]
- Li, J.J. Studies on the Origin of Chinese Mudan (Tree Peony). J. Beijing For. Univ. 1998, 20, 22–26. [Google Scholar]
- Cheng, F.Y. Advances in the Breeding of tree peonies and a Cultivar System for the Cultivar Group. International Journal of Plant Breeding 2007, 2, 89–104. [Google Scholar]
- Pei, Y.L. Preliminary study on the origin of cultivated peony. Northern Horticulture. 1996, 2, 55. [Google Scholar]
- Meng, L.; Zheng, G.S. Phylogenetic Relationship Analysis among Chinese Wild Species and Cultivars of Paeonia sect.Moutan Using RAPD Markers. Sci. Silvae Sin. 2004, 40, 110–115. [Google Scholar]
- Hou, X.G.; Yin, W.L.; Li, J.J.; Wang, H.F. Phylogenetic relationship of dwarf tree peony cultivars by AFLP analysis. J. Beijing For. Univ. 2006, 28, 73–77. [Google Scholar]
- Yu, H.P. Development of simple sequence repeat (SSR) markers from Paeonia ostii and its unilization in genetic relationship analysis. Beijing For. Univ. 2013, 164, 58–64. [Google Scholar]
- Chou, H.; Tang, H. Interspecific Relationship among the Wild Species of Paeonia Sect. Moutan DC with ITS and matK Sequence. Bull. Bot. Res. 2017, 37, 603–612. [Google Scholar] [CrossRef]
- Lin, Q.; Zhou, Z.; Zhao, X.; Pan, K.; Hong, D. Interspecific Relationships among the Wild Species of Paeonia Sect.Moutan DC.Based on DNA Sequences of Adh Gene Family. Acta Hortic. Sin. 2004, 31, 627–632. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, Z.Q.; Lin, Q.B.; Pan, K.Y.; Hong, D.Y. Molecular evidence for the interspecific relationships in Paeonia sect. Moutan: PCR-RFLP and sequence analysis of glycerol-3-phosphate acyltransferase (GPAT) gene. Acta Phytotaxon. Sin. 2004, 42, 236–244. [Google Scholar]
- Yuan, T.; Wang, L.Y. Pollen Morphology of Several Tree Peony Wild Species and Discussion on Its Evolution and Taxonomy. J. Beijing For. Univ. 1999, 21, 17–21. [Google Scholar]
- Wang, L.Y. Atlas of Tree Peony Varieties in China; Beijing China For. Press: Beijing, China, 1997; pp. 150–200. ISBN 978-7-5038-1770-0. [Google Scholar]
- Zhao, L.Y. Cultivation and Appreciation of Tree Peony in China; Beijing Gold. Shield Press: Beijing, China, 2004; pp. 46–50. ISBN 7-5082-2880-4. [Google Scholar]
- Wang, L.Y.; Yuan, T. Pictorial Record of Chinese Tree Peony Varieties: Continuation; Beijing China For. Press: Beijing, China, 2015; pp. 129–131. ISBN 978-7-5038-7915-9. [Google Scholar]
- Li, J.J.; Zhang, X.F.; Zhao, X.Q. Tree Peony of China; Beijing Encycl. China Press: Beijing, China, 2011; pp. 49–89. ISBN 978-7-5000-8522-5. [Google Scholar]
- Wang, L.Y. Observation on the morphological of flower bud differentiation of cultivars of tree peony and the analysis on the formation of flower forms. Acta Hortic. Sin. 1986, 13, 203–208. [Google Scholar]
- Hosoki, T.; Kimura, D.; Hasegawa, R.; Nagasako, T.; Nishimoto, K.; Ohta, K.; Sugiyama, M.; Haruki, K. Comparative study of Chinese tree peony cultivars by random amplified polymorphic DNA (RAPD) analysis. Sci. Hortic. 1997, 70, 67–72. [Google Scholar] [CrossRef]
- Yu, H. He Ze Tree Peony; Shandong Science and Technology Press: Shandong, China, 1980; pp. 15–50. [Google Scholar]
- Bruun-Lund, S.; Clement, W.L.; Kjellberg, F.; Rønsted, N. First plastid phylogenomic study reveals potential cyto-nuclear discordance in the evolutionary history of Ficus L. (Moraceae). Mol. Phylogenetics Evol. 2017, 109, 93–104. [Google Scholar] [CrossRef]
- Li, H.; Yi, T.; Gao, L.; Ma, P.; Zhang, T.; Yang, J.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Wang, R.; Liu, K.; Zhang, X.; Chen, W.; Qu, X.; Fan, S. Comparative Plastomes and Phylogenetic Analysis of Cleistogenes and Closely Related Genera (Poaceae). Front. Plant Sci. 2021, 12, 638597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-D.; Jin, J.-J.; Chen, S.-Y.; Chase, M.W.; Soltis, D.E.; Li, H.-T.; Yang, J.-B.; Li, D.-Z.; Yi, T.-S. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. New Phytol. 2017, 214, 1355–1367. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.A.; Azim, M.K. Variations in intergenic spacer rpl20-rps12 of mango (Mangifera indica) chloroplast DNA: Implications for cultivar identification and phylogenetic analysis. Plant Syst. Evol. 2011, 292, 249–255. [Google Scholar] [CrossRef]
- Suzuki, J.Y.; Geib, S.M.; Carlsen, M.M.; Henriquez, C.L.; Amore, T.D.; Sim, S.B.; Matsumoto, T.K.; Keith, L.M.; Myers, R.Y. Development of chloroplast single nucleotide polymorphisms (SNPs) as a tool towards interspecies typing of Anthurium germplasm. Acta Horticulturae. 2017, 1167. [Google Scholar] [CrossRef]
- Drouin, G.; Daoud, H.; Xia, J. Relative rates of synonymous substitutions in the mitochondrial, chloroplast and nuclear genomes of seed plants. Mol. Phylogenetics Evol. 2008, 49, 827–831. [Google Scholar] [CrossRef]
- Hu, S.Y. A cytological study of plastid inheritance in angiosperms. Acta Bot. Sin. 1997, 39, 363–371. [Google Scholar]
- Zhang, Q.; Liu, Y.; Sodmergen. Examination of the Cytoplasmic DNA in Male Reproductive Cells to Determine the Potential for Cytoplasmic Inheritance in 295 Angiosperm Species. Plant Cell Physiol. 2003, 44, 941–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.-L.; Zou, X.-H.; Zhou, Z.-Q.; Liu, J.; Xu, C.; Yu, J.; Wang, Q.; Zhang, D.-M.; Wang, X.-Q.; Ge, S.; et al. Multiple species of wild tree peonies gave rise to the ‘king of flowers’, Paeonia suffruticosa Andrews. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Nie, L.; Xu, Z.; Li, P.; Wang, Y.; He, C.; Song, J.; Yao, H. Comparative and Phylogenetic Analysis of the Complete Chloroplast Genomes of Three Paeonia Section Moutan Species (Paeoniaceae). Front. Genet. 2020, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Guo, S.; Xu, J.; He, L.; Carlson, J.E.; Hou, X. Phylogenetic analysis based on chloroplast genome uncover evolutionary relationship of all the nine species and six cultivars of tree peony. Ind. Crop. Prod. 2020, 153, 112567. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, 18. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.-J.; Moore, M.J.; Li, D.-Z.; Yi, T.-S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Poczai, P.; Hyvönen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An Online Program for the Versatile Plotting of Organelle Genomes. Front. Genet. 2020, 11, 576124. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.M.; Li, W.-H. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal alignment: Finding rearrangements during alignment. Bioinformatics 2003, 19, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Mayor, C.; Brudno, M.; Schwartz, J.R.; Poliakov, A.; Rubin, E.M.; Frazer, K.A.; Pachter, L.S.; Dubchak, I. VISTA: Visualizing global DNA sequence alignments of arbitrary length. Bioinformatics 2000, 16, 1046–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Kuma, K.-i.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- iPlant. Flora Reipublicae Popularis Sinicae. Available online: http://www.iplant.cn/frps (accessed on 9 May 2022).
- Maddison, G.K.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. 2004. Available online: http://mesquiteproject.org (accessed on 21 May 2022).
- Xiang, Y.; Huang, C.-H.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of Rosaceae Fruit Types Based on Nuclear Phylogeny in the Context of Geological Times and Genome Duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fennoy, S.L.; Bailey-serres, J. Synonymous codon usage in Zea mays L. nuclear genes is varied by levels of C and G-ending codons. Nucleic Acids Res. 1993, 21, 5294–5300. [Google Scholar] [CrossRef] [PubMed]
- Ikemura, T. Codon usage and tRNA content in unicellular and multicellular organisms. Mol. Biol. Evol. 1985, 2, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, F. Biological Genome Evolution and Codon Usage; National Defense Industry Press: Beijing, China, 2014; pp. 19–51. ISBN 978-7-118-09366-7. [Google Scholar]
- Kaundun, S.S.; Matsumoto, S. Heterologous nuclear and chloroplast microsatellite amplification and variation in tea, Camellia sinensis. Genome 2002, 45, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. CHAPTER 2—Plastid Chromosomes: Structure and Evolution. Mol. Biol. Plast. 1991, 7, 5–53. [Google Scholar] [CrossRef]
- Han, X.Y.; Wang, L.S.; Shu, Q.Y.; Liu, Z.A.; Xu, S.X.; Tetsumura, T. Molecular Characterization of Tree Peony Germplasm Using Sequence-Related Amplified Polymorphism Markers. Biochem. Genet. 2008, 46, 162–179. [Google Scholar] [CrossRef]
- Wang, L.Y. A Map of Tree Peony Cultivars in China; Beijing China For. Publ. House: Beijing, China, 1997; pp. 150–200. ISBN 978-7-5038-1770-0. [Google Scholar]
- Yu, L.; He, L.X.; Li, J.J. Comparative Studies on Chromosome in Varieties of Paeonia rockii and Paeonia suff ruticosa. Acta Hortic. Sin. 1997, 24, 79–83. [Google Scholar]
- Han, X.; Wang, L.; Liu, Z.a.; Jan, D.R.; Shu, Q. Characterization of sequence-related amplified polymorphism markers analysis of tree peony bud sports. Sci. Hortic. 2008, 115, 261–267. [Google Scholar] [CrossRef]
- Jiang, Z.; Geng, X.; Zhu, Z.; Sheng, Q. Research Progress on Cross Breeding of Tree Peony. Mol. Plant Breed. 2021. Available online: http://kns.cnki.net/kcms/detail/46.1068.s.20210527.1903.018.html (accessed on 17 October 2022).
- Shi, Q. Study on Genetic Diversity of Chinese Traditional Cultivars of Tree Peony. Chin. Acad. For. 2012. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1012423632.nh&DbName=CMFD2012 (accessed on 17 October 2022).
- Wei, J.; Peng, Y.; Yang, L. Comparison of Floral Organ Morphology and Development of Single and Double Petaled Malus halliana. Acta Bot. Boreal 2021, 41, 2072–2079. [Google Scholar] [CrossRef]
- Jiang, S.; Yi, X.-W.; Xu, T.-L.; Yang, Y.; Yu, C.; Luo, L.; Cheng, T.-R.; Wang, J.; Zhang, Q.-X.; Pan, H.-T. Genetic analysis of petal number in Rosa. Plant Sci. J. 2021, 39, 142–151. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Liu, Q.-L. Research Advances in the Formation Mechanism and Genetic Characters of Double Flowers. Acta Bot. Boreal 2009, 29, 0832–0841. [Google Scholar]
- Zhu, G.; Li, J.; Ni, S.; Fan, Z.-Q.; Yin, H.-F.; Li, X.; Zhou, X.-W. The potential role of B-function gene involved in floral development for double flowers formation in Camellia changii Ye. Afr. J. Biotechnol. 2011, 10, 16757–16762. [Google Scholar]
- Lehmann, N.L.; Sattler, R. Floral Development and Homeosis in Actaea rubra (Ranunculaceae). Int. J. Plant Sci. 1994, 155, 658–671. [Google Scholar] [CrossRef]
- Zhao, H.-J.; Zhang, W.-T. Floral type classification and genesis of Paeonia lactiflora. North. Hortic. 1999, 05, 26–27. [Google Scholar]
- Gao, Z.; Mu, S.; Li, X.; Pan, H. The evolvement rule analysis of tree peony flower types using molecular mechanism of model plants flower development. Adv. Ornam. Hortic. Res. China 2009, 2009, 129–133. [Google Scholar]
- Irish, V. The ABC model of floral development. Curr. Biol. 2017, 27, 887–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.Q. Screening of candidate genes regulating the anemone flower type and study of expression and regulation mechanism in herbaceous peony (Paeonia lactiflora Pall). Yangzhou Univ. 2019, 10–15. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1020025126.nh&DbName=CDFD2020 (accessed on 17 October 2022).
- Lin, Z.; Damaris, R.N.; Shi, T.; Li, J.; Yang, P. Transcriptomic analysis identifies the key genes involved in stamen petaloid in lotus (Nelumbo nucifera). BMC Genom. 2018, 19, 554. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | All Length | GC% | LSC Length | GC% | SSC Length | GC% | IR Length | GC% | GenBank Accession Numbers |
|---|---|---|---|---|---|---|---|---|---|
| P. ostii cv. ‘Feng Dan’ | 152,153 bp | 38.3 | 85,373 bp | 36.7 | 17,054 bp | 32.7 | 24,863 bp | 43.1 | MG585274.1 |
| P. suffruticosa cv. ‘Da Jin Fen’ | 152,835 bp | 38.3 | 84,485 bp | 36.6 | 17,056 bp | 32.6 | 25,647 bp | 43.1 | ON243818 |
| P.×lemoine ‘High Noon’ | 152,519 bp | 38.4 | 84,213 bp | 36.7 | 17,026 bp | 32.8 | 25,640 bp | 43.1 | ON243821 |
| P. suffruticosa cv. ‘Da Zong Zi’ | 152,820 bp | 38.4 | 84,471 bp | 36.6 | 17,051 bp | 32.6 | 25,649 bp | 43.1 | ON243822 |
| P. suffruticosa cv. ‘Greendragon sleeping pool’ | 152,820 bp | 38.4 | 84,475 bp | 36.6 | 17,051 bp | 32.6 | 25,647 bp | 43.1 | ON243820 |
| P. suffruticosa cv. ‘Yao Huang’ | 152,819 bp | 38.4 | 84,474 bp | 36.6 | 17,051 bp | 32.6 | 25,647 bp | 43.1 | ON227529 |
| P. suffruticosa cv. ‘Shou An Hong’ | 152,819 bp | 38.4 | 84,474 bp | 36.6 | 17,051 bp | 32.6 | 25,647 bp | 43.1 | MK701996.1 |
| P. suffruticosa cv. ‘Dou Lv’ | 152,819 bp | 38.4 | 84,474 bp | 36.6 | 17,051 bp | 32.6 | 25,647 bp | 43.1 | ON243819 |
| P. suffruticosa cv. ‘Cao Zhou Hong’ | 152,835 bp | 38.3 | 84,485 bp | 36.6 | 17,056 bp | 32.6 | 25,647 bp | 43.1 | ON227528 |
| P. suffruticosa cv. ‘Guan Qun Fang’ | 152,820 bp | 38.4 | 84,471 bp | 36.6 | 17,053 bp | 32.6 | 25,649 bp | 43.1 | ON209192 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Huang, X.-C.; Wang, Y.-L.; Zhang, R.-J.; Shi, D.-Y.; Li, T.-F.; Zhou, G.-C.; Xue, J.-Y. Plastid Phylogenomics of Paeonia and the Evolution of Ten Flower Types in Tree Peony. Genes 2022, 13, 2229. https://doi.org/10.3390/genes13122229
Li W, Huang X-C, Wang Y-L, Zhang R-J, Shi D-Y, Li T-F, Zhou G-C, Xue J-Y. Plastid Phylogenomics of Paeonia and the Evolution of Ten Flower Types in Tree Peony. Genes. 2022; 13(12):2229. https://doi.org/10.3390/genes13122229
Chicago/Turabian StyleLi, Wen, Xin-Cheng Huang, Yi-Lei Wang, Rui-Ju Zhang, Dong-Yan Shi, Teng-Fei Li, Guang-Can Zhou, and Jia-Yu Xue. 2022. "Plastid Phylogenomics of Paeonia and the Evolution of Ten Flower Types in Tree Peony" Genes 13, no. 12: 2229. https://doi.org/10.3390/genes13122229
APA StyleLi, W., Huang, X. -C., Wang, Y. -L., Zhang, R. -J., Shi, D. -Y., Li, T. -F., Zhou, G. -C., & Xue, J. -Y. (2022). Plastid Phylogenomics of Paeonia and the Evolution of Ten Flower Types in Tree Peony. Genes, 13(12), 2229. https://doi.org/10.3390/genes13122229