
Complete Chloroplast Genome Sequences of Four Species in the Caladium Genus: Comparative and Phylogenetic Analyses


 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Structure of the Chloroplast Genomes of the Four Caladium Species
2.2. Analysis of Contraction and Expansion of the IR Region
2.3. GView Analysis
2.4. Analysis of Chloroplast Microsatellites and Repeat Sequences
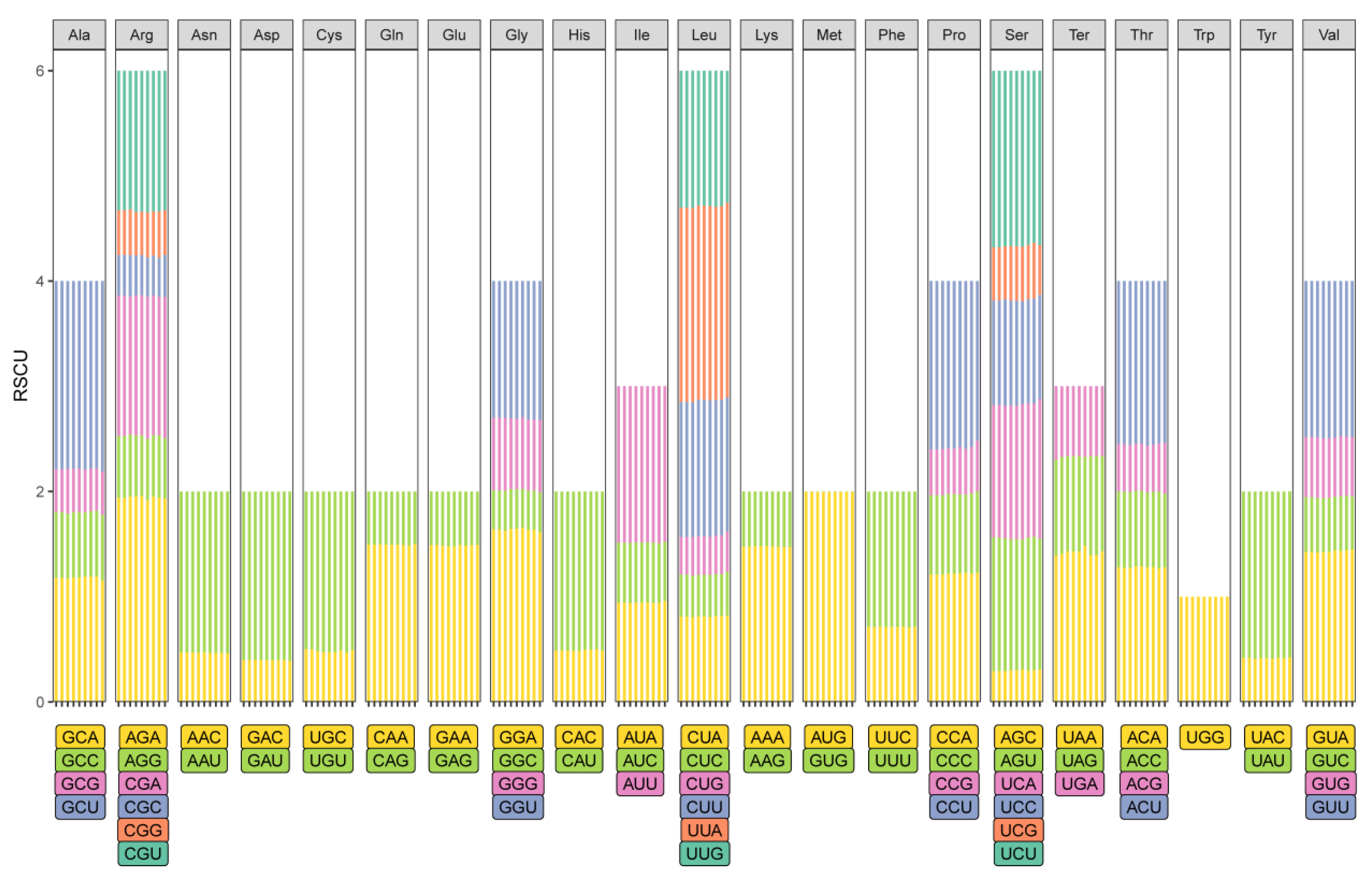
2.5. Analysis of Selection Pressure and Codon Bias
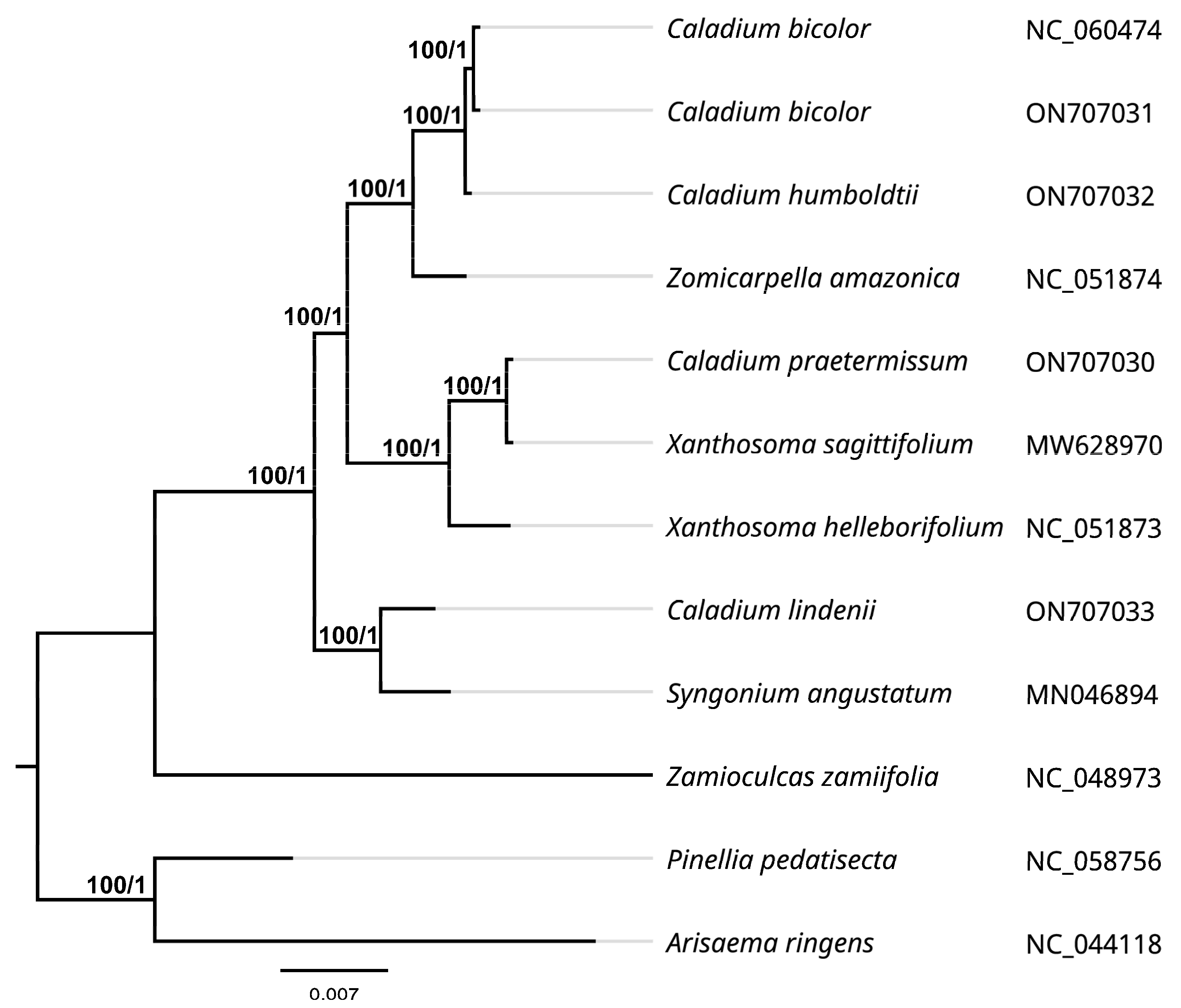
2.6. Phylogenetic Relationship Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. DNA Extraction and High-Throughput Sequencing
4.3. Chloroplast Genome Assembly and Annotation
4.4. Comparative Analysis of Chloroplast Genomes
4.5. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.S.; Chen, J.J.; Cao, Y.M.; Duan, J.X.; Cai, X.D. Induction of tetraploids in ‘Red Flash’ Caladium using colchicine and oryzalin: Morphological, cytological, photosynthetic and chilling tolerance analysis. Sci. Hortic. 2020, 275, 109524. [Google Scholar] [CrossRef]
- Cao, Z.; Deng, Z.A. Morphological, cytological and molecular marker analyses of ‘Tapestry’ Caladium variants reveal diverse genetic changes and enable association of leaf coloration pattern loci with molecular markers. Plant Cell Tiss. Org. 2020, 143, 363–375. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, Y.S.; Duan, J.X.; Cao, Y.M.; Cai, X.D. Morphological, cytological, and pigment analysis of leaf color variants regenerated from long-term subcultured Caladium callus. In Vitro Cell Dev. Biol.-Plant 2021, 57, 60–71. [Google Scholar] [CrossRef]
- Croat, T.B.; Delannay, X.; Ortiz, O.O.; Jiménez, P.D. A review of the Aroid Tribe Caladieae with the description of three new species of Caladium and seven new species of Syngonium (Araceae). Novon J. Bot. Nomencl. 2019, 27, 38–64. [Google Scholar] [CrossRef]
- Akhigbemen, A.M.; Ozolua, R.I.; Bafor, E.E.; Okwuofu, E.O. Evaluation of some neuropharmacological effects of Caladium bicolor aiton (Araceae) leaf extracts in mice. Metab. Brain Dis. 2019, 34, 537–544. [Google Scholar] [CrossRef]
- Isah, T. Changes in the biochemical parameters of albino, hyperhydric and normal green leaves of Caladium bicolor cv. “Bleeding hearts” in vitro long-term cultures. J. Photoch. Photobiol. B 2019, 191, 88–98. [Google Scholar] [CrossRef]
- Deng, Z.A.; Harbaugh, B.K. Independent inheritance of leaf shape and main vein color in Caladium. J. Am. Soc. Hortic. Sci. 2006, 131, 53–58. [Google Scholar] [CrossRef]
- Ahmed, E.U.; Hayashi, T.; Yazawa, S. Leaf color stability during plant development as an index of leaf color variation among micropropagated Caladium. HortScience 2004, 39, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Kokalis-Burelle, N.; Brito, J.A.; Hartman, R.D. Susceptibility of seven Caladium (Caladium × hortulanum) cultivars to Meloidogyne arenaria, M. enterolobii, M. floridensis, M. incognita, and M. javanica. J. Nematol. 2017, 49, 457–461. [Google Scholar]
- Cao, Z.; Deng, Z.A. De novo assembly, annotation, and characterization of root transcriptomes of three Caladium cultivars with a focus on necrotrophic pathogen resistance/defense-related genes. Int. J. Mol. Sci. 2017, 18, 712. [Google Scholar] [CrossRef]
- Hussain, R.; Younis, A.; Riaz, A.; Tariq, U.; Ali, S.; Ali, A.; Raza, S. Evaluating sustainable and environment friendly substrates for quality production of potted Caladium. Int. J. Recycl. Org. 2017, 6, 13–21. [Google Scholar] [CrossRef]
- Cao, Z.; Sui, S.Z.; Cai, X.D.; Yang, Q.; Deng, Z.A. Somaclonal variation in ‘Red Flash’ Caladium: Morphological, cytological and molecular characterization. Plant Cell Tiss. Org. 2016, 126, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Sui, S.Z.; Yang, Q.; Deng, Z.A. Inheritance of rugose leaf in Caladium and genetic relationships with leaf shape, main vein color, and leaf spotting. J. Am. Soc. Hortic. Sci. 2016, 141, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Ekeke, C.; Agbagwa, I.O. Anatomical characteristics of Nigerian variants of Caladium bicolor (Aiton) Vent. (Araceae). Afr. J. Plant Sci. 2016, 10, 121–129. [Google Scholar]
- Ahmed, E.U.; Hayashi, T.; Yazawa, S. Auxins increase the occurrence of leaf-colour variants in Caladium regenerated from leaf explants. Sci. Hortic. 2004, 100, 153–159. [Google Scholar] [CrossRef]
- Deng, Z.A.; Peres, N.A. ‘Sea Foam Pink’ Caladium. HortScience 2019, 54, 1637–1640. [Google Scholar] [CrossRef]
- Yu, J.L.; Boyd, N.S. Tolerance of Caladium cultivars florida cardinal and florida fantasy to sulfonylurea herbicides. HortScience 2018, 53, 850–858. [Google Scholar] [CrossRef]
- Cai, X.D.; Cao, Z.; Xu, S.X.; Deng, Z.A. Induction, regeneration and characterization of tetraploids and variants in ‘Tapestry’ Caladium. Plant Cell Tiss. Org. 2015, 120, 689–700. [Google Scholar] [CrossRef]
- Maia, A.C.D.; Schlindwein, C. Caladium bicolor (Araceae) and Cyclocephala celata (Coleoptera, Dynastinae): A well-established pollination system in the northern atlantic rainforest of pernambuco, Brazil. Plant Biol. 2006, 8, 529–534. [Google Scholar] [CrossRef]
- Deng, Z.A.; Goktepe, F.; Harbaugh, B.K. Inheritance of leaf Spots and their genetic relationships with leaf shape and vein color in Caladium. J. Am. Soc. Hortic. Sci. 2008, 133, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Madison, M. Notes on Caladium (Araceae) and its allies. Selbyana 1981, 5, 342–377. [Google Scholar]
- Croat, T. Taxonomic status of neotropical Araceae. Aroideana 1994, 17, 33–60. [Google Scholar]
- Deng, Z.A.; Goktepe, F.; Harbaugh, B.K.; Hu, J.G. Assessment of genetic diversity and relationships among Caladium cultivars and species using molecular markers. J. Am. Soc. Hortic. Sci. 2007, 132, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Deng, Z.A. Interspecific genome size and chromosome number variation shed new light on species classification and evolution in Caladium. J. Am. Soc. Hortic. Sci. 2014, 139, 449–459. [Google Scholar] [CrossRef]
- Loh, J.P.; Kiew, R.; Kee, A.; Gan, L.H.; Gan, Y.Y. Amplified fragment length polymorphism (AFLP) provides molecular markers for the identification of Caladium bicolor cultivars. Ann. Bot. 1999, 84, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The complete chloroplast genome sequence of Pelargonium × hortorum: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef]
- Firetti, F.; Zuntini, A.R.; Gaiarsa, J.W.; Oliveira, R.S.; Lohmann, L.G.; Van Sluys, M.A. Complete chloroplast genome sequences contribute toplant species delimitation: A case study of the Anemopaegma species complex. Am. J. Bot. 2017, 104, 1493–1509. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.S.; Li, P.; Qiu, Y.X. The complete chloroplast genomes of three Cardiocrinum (Liliaceae) species: Comparative genomic and phylogenetic analyses. Front. Plant Sci. 2017, 7, 2054. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.R.; Kim, K.; Lee, B.Y.; Lim, C.E. Complete chloroplast genomes of all six Hosta species occurring in Korea: Molecular structures, comparative, and phylogenetic analyses. BMC Genom. 2019, 20, 833. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.L.; Li, Q.; Hu, Z.G.; Li, X.W.; Chen, S.L. The complete Amomum kravanh chloroplast genome sequence and phylogenetic analysis of the commelinids. Molecules 2017, 22, 1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.X.; Nie, J.; Xiao, L.; Hu, Z.G.; Wang, B. Comparative chloroplast genome analysis of rhubarb botanical origins and development of specific identification markers. Molecules 2018, 23, 2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.H.; Shi, S.; Li, J.L.; Jing, Y.; Wang, L.; Yang, X.Y.; Guo, L.; Zhou, S.L.; Sun, F.J. Phylogeny of Maleae (Rosaceae) based on multiple chloroplast regions: Implications to genera Circumscription. Biomed. Res. Int. 2018, 2018, 7627191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.M.; Qi, X.J.; Chen, J.Y.; Sun, L.M.; Zhong, Y.P.; Fang, J.B.; Hu, C.G. The complete chloroplast genome sequence of Actinidia arguta using the PacBio RS II platform. PLoS ONE 2018, 13, e0197393. [Google Scholar] [CrossRef]
- Li, D.M.; Zhao, C.Y.; Liu, X.F. Complete chloroplast genome sequences of Kaempferia galanga and Kaempferia elegans: Molecular structures and comparative analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, L.H.M.; Lohmann, L.G. Plastome rearrangements in the “Adenocalymma-Neojobertia” clade (Bignonieae, Bignoniaceae) and its phylogenetic implications. Front. Plant Sci. 2017, 8, 1875. [Google Scholar] [CrossRef] [Green Version]
- Thode, V.A.; Lohmann, L.G. Comparative chloroplast genomics at low taxonomic levels: A case study using Amphilophium (Bignonieae, Bignoniaceae). Front. Plant Sci. 2019, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Chiapella, J.O.; Barfuss, M.H.J.; Xue, Z.Q.; Greimler, J. The plastid genome of Deschampsia cespitosa (Poaceae). Molecules 2019, 24, 216. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.X.; Chen, X.L.; Nie, L.P.; Sun, W.; Hu, H.Y.; Lin, Y.L.; Li, H.T.; Zheng, X.L.; Song, J.Y.; Yao, H. Comparison and phylogenetic analysis of chloroplast genomes of three medicinal and edible Amomum species. Int. J. Mol. Sci. 2019, 20, 4040. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.X.; Nie, L.P.; Sun, W.; Xu, Z.C.; Wang, Y.; Yu, J.; Song, J.Y.; Yao, H. Comparative and phylogenetic analyses of ginger (Zingiber offcinale) in the family Zingiberaceae based on the complete chloroplast genome. Plants 2019, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Barrett, C.F.; Baker, W.J.; Comer, J.R.; Conran, J.G.; Lahmeyer, S.C.; Leebens-Mack, J.H.; Li, J.; Lim, G.S.; MayfieldJones, D.R.; Perez, L.; et al. Plastid genomes reveal support for deep phylogenetic relationships and extensive rate variation among palms and other commelinid monocots. N. Phytol. 2016, 209, 855–870. [Google Scholar] [CrossRef]
- Ferrarini, M.; Moretto, M.; Ward, J.A.; Surbanovski, N.; Stevanovic, V.; Giongo, L.; Viola, R.; Cavalieri, D.; Velasco, R.; Cestaro, A.; et al. An evaluation of the PacBio RS platform for sequencing and de novo assembly of a chloroplast genome. BMC Genom. 2013, 14, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.H.; Gui, S.T.; Guan, Z.W.; Pan, L.; Wang, S.Z.; Ke, W.D.; Liang, D.Q.; Ding, Y. A precise chloroplast genome of Nelumbo nucifera (Nelumbonaceae) evaluated with Sanger, Illumina MiSeq, and PacBio RS II sequencing platforms: Insight into the plastid evolution of basal eudicots. BMC Plant Biol. 2014, 14, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.W.; Hu, Z.G.; Lin, X.H.; Li, Q.; Gao, H.H.; Luo, G.A.; Chen, S.L. High-throughput pyrosequencing of the complete chloroplast genome of Magnolia officinalis and its application in species identification. Acta Pharm. Sin. 2012, 47, 124–130. [Google Scholar]
- Li, Y.F.; Sylvester, S.P.; Li, M.; Zhang, C.; Li, X.; Duan, Y.F.; Wang, X.R. The complete plastid genome of Magnolia zenii and genetic comparison to Magnoliaceae species. Molecules 2019, 24, 261. [Google Scholar] [CrossRef] [PubMed]
- Brunkard, J.O.; Runkel, A.M.; Zambryski, P.C. Chloroplast extend stromules independently and in response to internal redox signals. Proc. Natl. Acad. Sci. USA 2015, 112, 10044–10049. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.M.; Deng, Y.F.; Wang, J. The complete chloroplast genomes of Echinacanthus species (Acanthaceae): Phylogenetic relationships, adaptive evolution, and screening of molecular markers. Front. Plant Sci. 2019, 9, 1989. [Google Scholar] [CrossRef] [Green Version]
- Pauwels, M.; Vekemans, X.; Godé, C.; Frérot, H.; Castric, V.; Saumitou-Laprade, P. Nuclear and chloroplast DNA phylogeography reveals vicariance among european populations of the model species for the study of metal tolerance, Arabidopsis halleri (Brassicaceae). N. Phytol. 2012, 193, 916–928. [Google Scholar] [CrossRef]
- Wu, L.W.; Nie, L.P.; Wang, Q.; Xu, Z.C.; Wang, Y.; He, C.N.; Song, J.Y.; Yao, H. Comparative and phylogenetic analyses of the chloroplast genomes of species of Paeoniaceae. Sci. Rep. 2021, 11, 14643. [Google Scholar] [CrossRef]
- Li, L.; Wu, Q.P.; Fang, L.; Wu, K.L.; Li, M.Z.; Zeng, S.J. Comparative chloroplast genomics and phylogenetic analysis of Thuniopsis and closely related genera within Coelogyninae (Orchidaceae). Front. Genet. 2022, 13, 850201. [Google Scholar] [CrossRef]
- Hanson, G.; Coller, J. Codon optimality, bias and usage in translation and mRNA decay. Nat. Rev. Mol. Cell Biol. 2017, 19, 2030. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wei, F.; Cai, Z.; Wei, Y.; Khan, A.; Miao, J.; Wei, K. Analysis of codon usage bias and evolution in the chloroplast genome of Mesona chinensis Benth. Dev. Genes Evol. 2021, 231, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; He, J.; Jia, X.; Qi, Q.; Liang, Z.; Zheng, H.; Ping, Y.; Liu, S.; Sun, J. Analysis of codon usage bias of mitochondrial genome in Bombyx moriand its relation to evolution. BMC Evol. Biol. 2014, 14, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Wei, L.L.; Ma, L.; Wu, Z.Q.; Gu, C.H.; Chen, K. Comparative analyses of chloroplast genomes from 13 Lagerstroemia (Lythraceae) species: Identification of highly divergent regions and inference of phylogenetic relationships. Plant Mol. Biol. 2020, 102, 659–676. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Sun, M.H.; Jiang, Z.S.; Shu, H.R.; Zhang, S.Z. Genome-wide analysis of the synonymous codon usage patterns in apple. J. Integr. Agr. 2016, 15, 983–991. [Google Scholar] [CrossRef]
- Ye, Y.J.; Xu, Y.C.; Li, D.M.; Tan, J.J.; Liu, J.M. Characterization of EST-SSR markers in Curcuma kwangsiensis S. K. Lee & C. F. Liang based on RNA sequencing and its application for phylogenetic relationship analysis and core collection construction. Genet. Resour. Evol. 2021, 68, 1503–1516. [Google Scholar]
- Dierckysens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. Organellar Genome DRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- Meade, J.C.; Shah, P.H.; Lushbaugh, W.B. Trichomonas vaginalis: Analysis of codon usage. Exp. Parasitol. 1997, 87, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Amiryousefi, A.; Hyvonen, J.; Poczai, P. Irscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2008, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; Domselaar, G.V. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.V.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Names | Size (bp)/GC Content (%) | Number of Gene | ||||||
|---|---|---|---|---|---|---|---|---|
| Genome | LSC | SSC | IR | PCGs | RNA/rRNA | RNA/tRNA | Total Genes | |
| C. bicolor | 162,933/35.87 | 89,209/34.17 | 21,170/29.07 | 26,277/41.5 | 86 | 37 | 8 | 131 |
| C. humboldtii | 162,776/35.91 | 88,986/34.26 | 21,236/28.99 | 26,277/41.5 | 86 | 37 | 8 | 131 |
| C. praetermissum | 165,286/35.64 | 91,168/33.76 | 21,150/29.27 | 26,484/41.42 | 86 | 37 | 8 | 131 |
| C. lindenii | 168,888/35.09 | 93,162/33.26 | 22,782/27.87 | 26,472/41.43 | 86 | 37 | 8 | 131 |
| Syngonium angustatum | 164,929/35.72 | 90,714/33.94 | 21,559/28.98 | 26,328/41.53 | 86 | 37 | 8 | 131 |
| Zomicarpella amazonica | 162,729/35.82 | 90,811/34.33 | 21,656/28.76 | 25,131/41.55 | 85 | 37 | 8 | 130 |
| Zamioculcas zamiifolia | 167,405/35.7 | 91,357/34.02 | 19,326/29.47 | 28,361/40.52 | 86 | 37 | 8 | 131 |
| Xanthosoma helleborifolium | 164,418/35.84 | 90,833/33.88 | 20,705/41.53 | 26,440/41.53 | 86 | 37 | 8 | 131 |
| Xanthosoma sagittifolium | 165,169/35.67 | 91,122/33.79 | 21,079/29.36 | 26,484/41.42 | 86 | 37 | 8 | 131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Liu, J.; Zhou, Y.; Zhu, G.; Tan, J.; Xu, Y. Complete Chloroplast Genome Sequences of Four Species in the Caladium Genus: Comparative and Phylogenetic Analyses. Genes 2022, 13, 2180. https://doi.org/10.3390/genes13122180
Ye Y, Liu J, Zhou Y, Zhu G, Tan J, Xu Y. Complete Chloroplast Genome Sequences of Four Species in the Caladium Genus: Comparative and Phylogenetic Analyses. Genes. 2022; 13(12):2180. https://doi.org/10.3390/genes13122180
Chicago/Turabian StyleYe, Yuanjun, Jinmei Liu, Yiwei Zhou, Genfa Zhu, Jianjun Tan, and Yechun Xu. 2022. "Complete Chloroplast Genome Sequences of Four Species in the Caladium Genus: Comparative and Phylogenetic Analyses" Genes 13, no. 12: 2180. https://doi.org/10.3390/genes13122180
APA StyleYe, Y., Liu, J., Zhou, Y., Zhu, G., Tan, J., & Xu, Y. (2022). Complete Chloroplast Genome Sequences of Four Species in the Caladium Genus: Comparative and Phylogenetic Analyses. Genes, 13(12), 2180. https://doi.org/10.3390/genes13122180