
Phylogenetic Analyses of Some Key Genes Provide Information on Pollinator Attraction in Solanaceae

Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition and Characterization of Plant Coding Sequences
2.2. Multiple Alignments and Gene Structure
2.3. Gene Trees Reconstruction
2.4. Molecular Evolutionary Analyses
3. Results
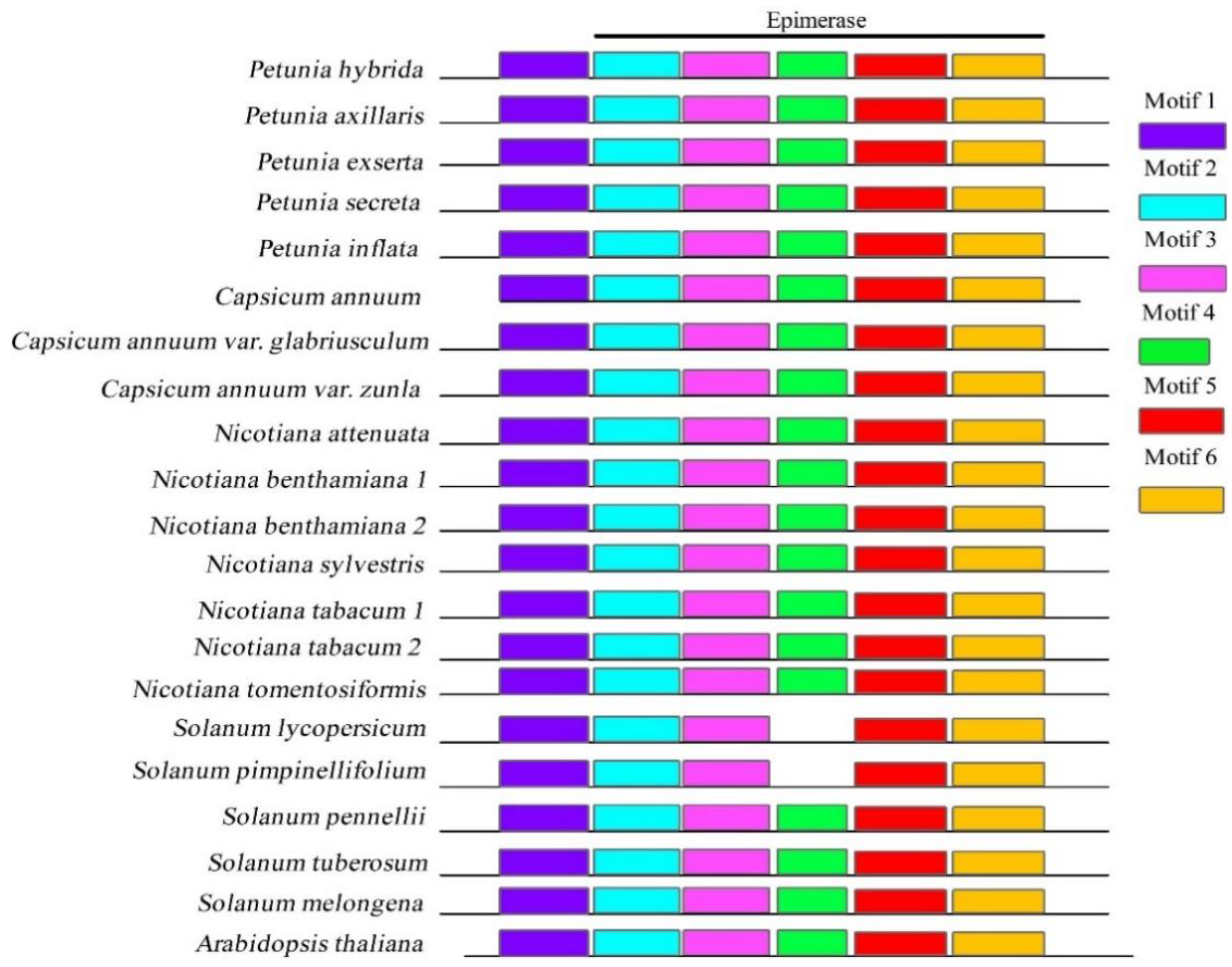
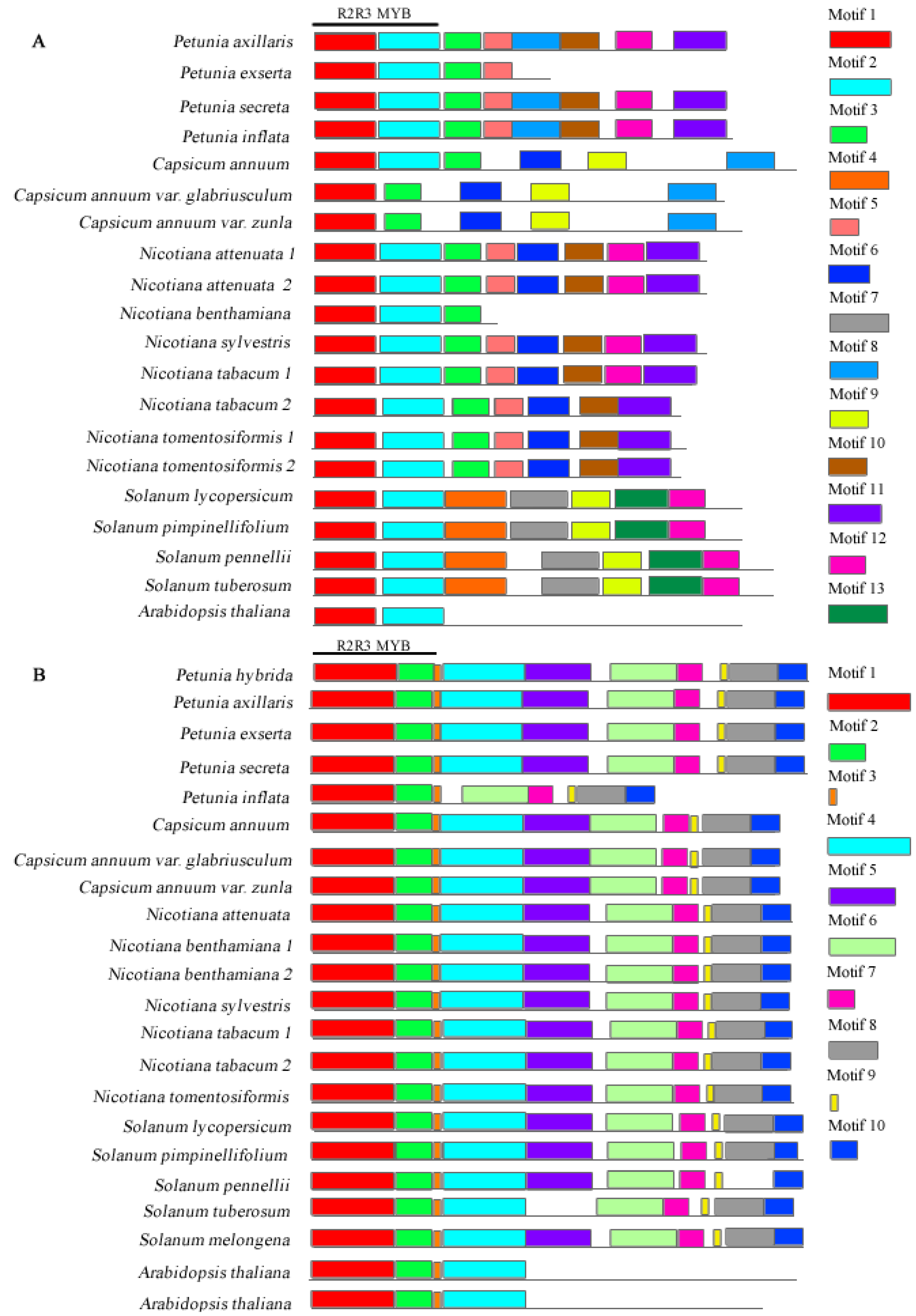
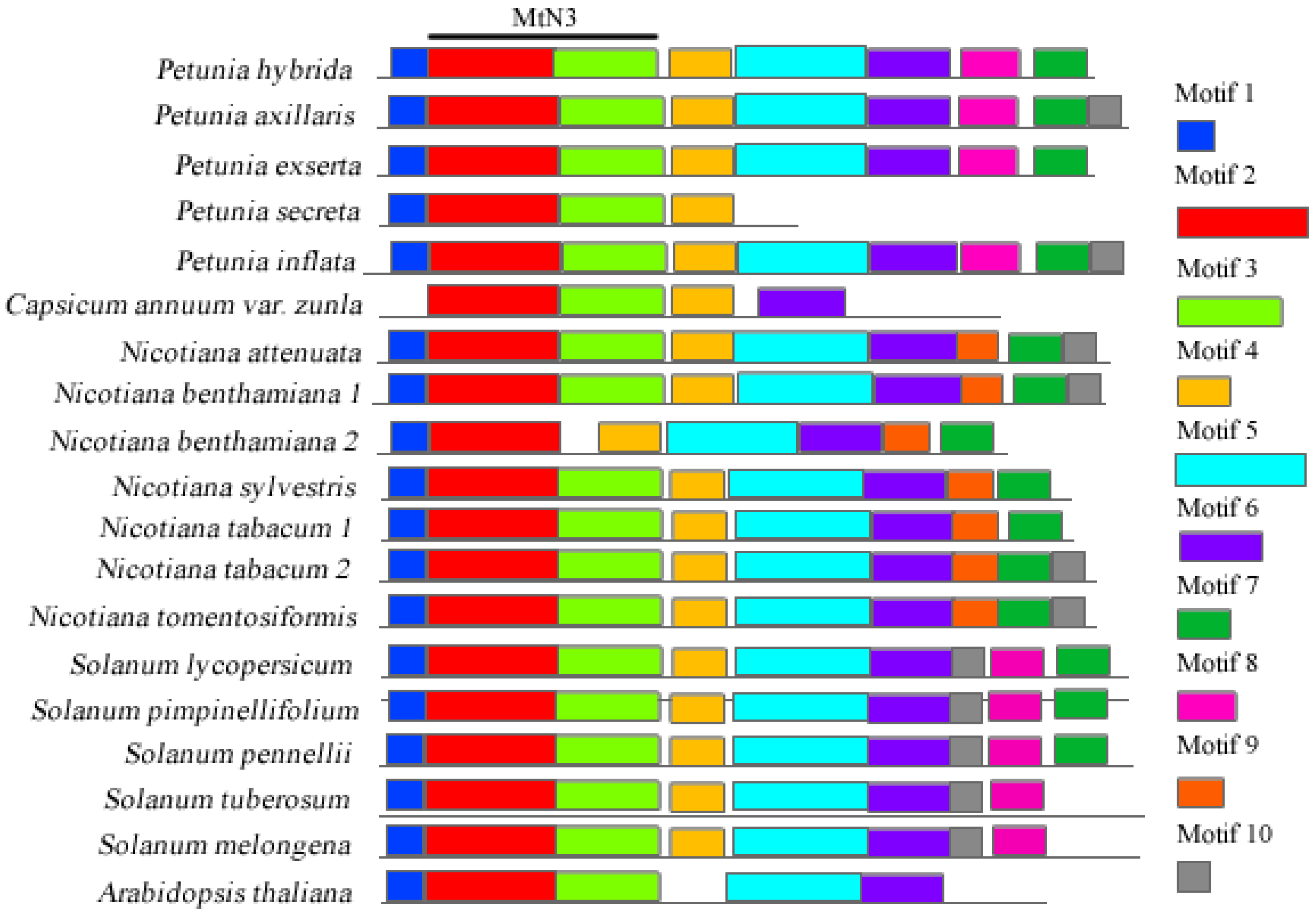
3.1. Identification of Gene Sequences in Solanaceae
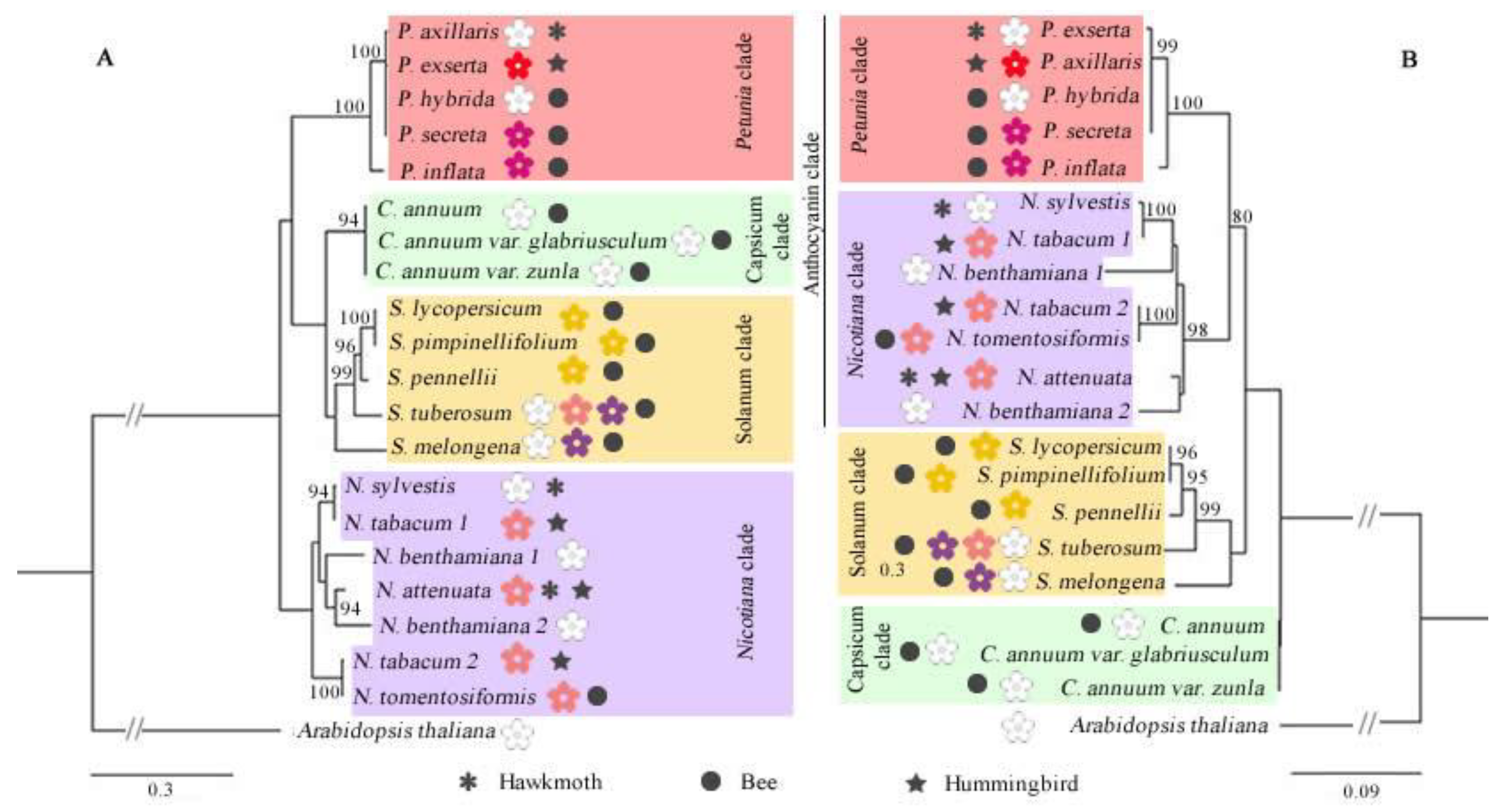
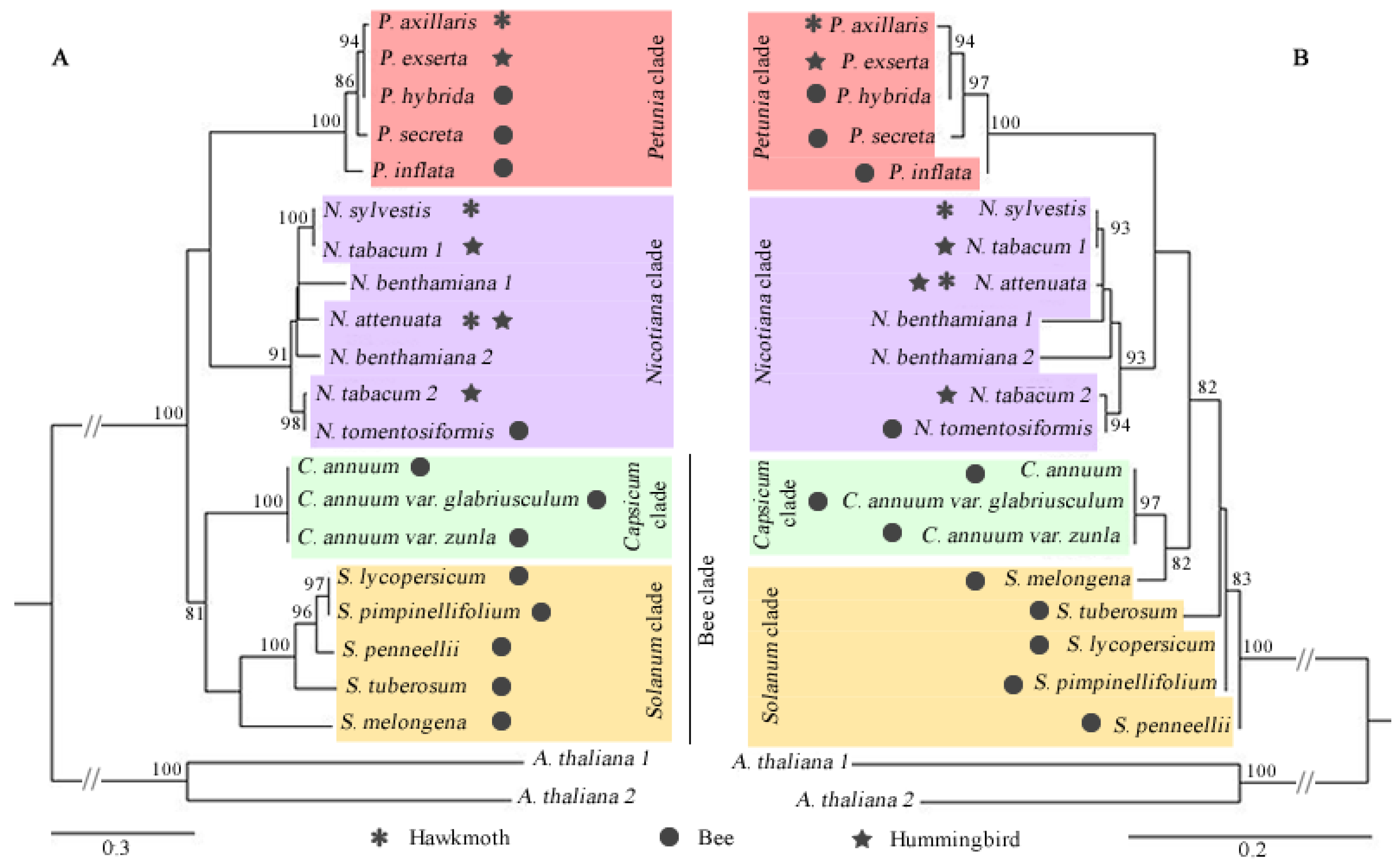
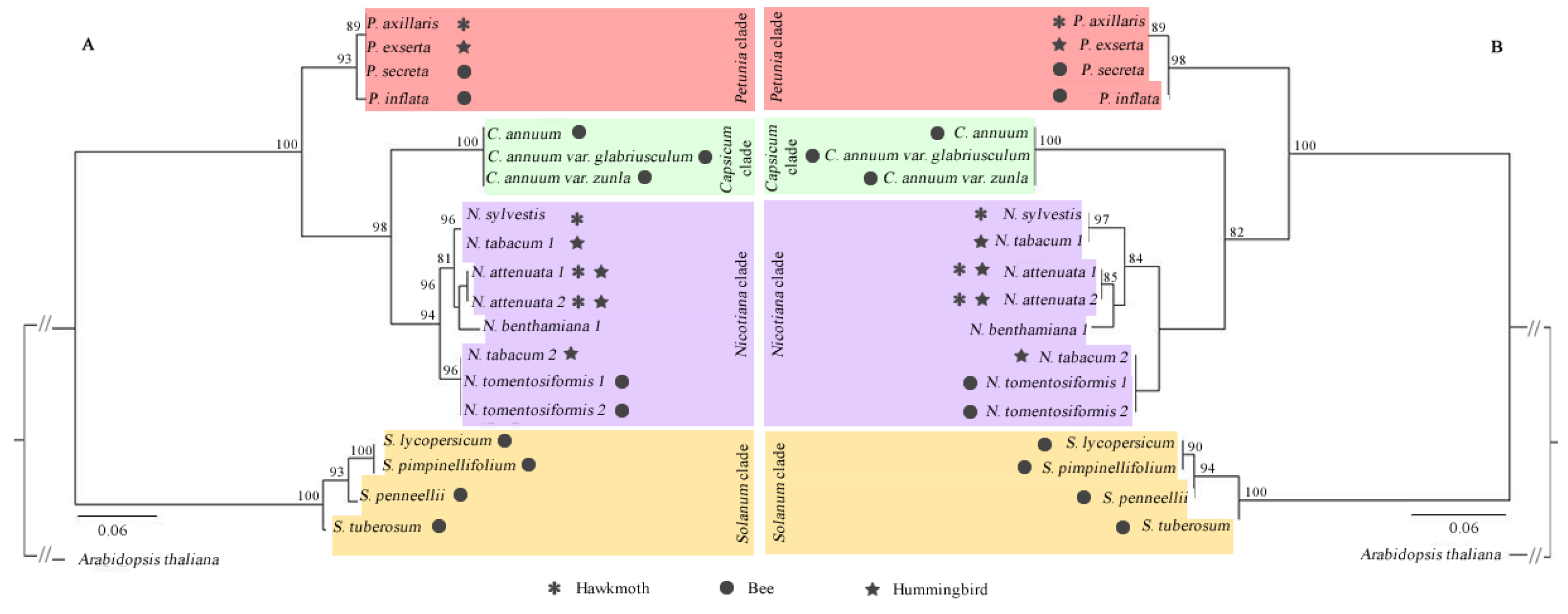
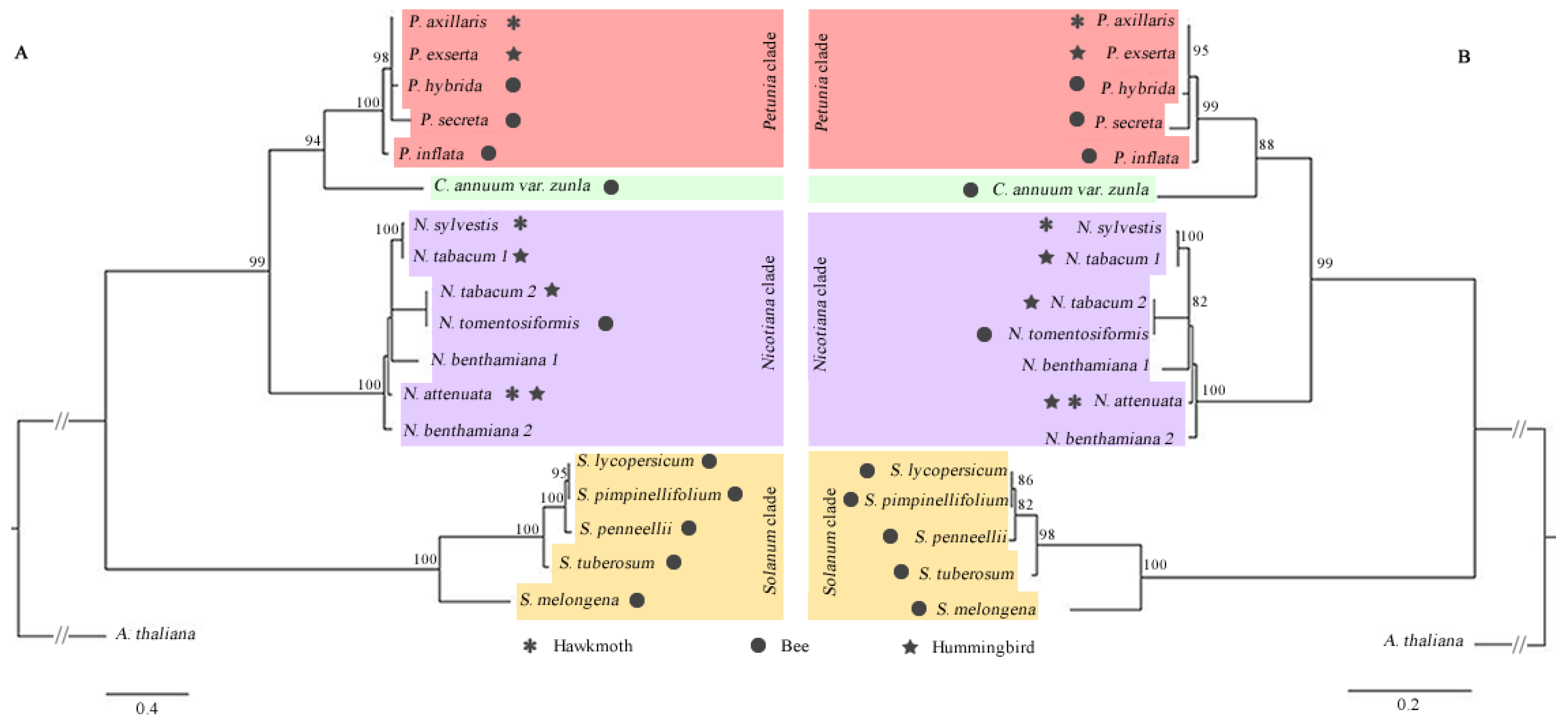
3.2. Phylogenetic Relationships
3.3. Analyses of Selection
4. Discussion
4.1. Visible Colors and UV-Light Response
4.2. Odor Emission
4.3. Floral Rewards
4.4. Gene Conservation by Purifying Selection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faegri, K.; van der Pijl, L. The Principles of Pollination Ecology, 3rd ed.; Pergamon Press: Oxford, UK, 1979; 244p. [Google Scholar]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Dellinger, A.S. Pollination syndromes in the 21st century: Where do we stand and where may we go? New Phytol. 2020, 228, 1193–1213. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.-Y.; Han, J.-X.; Han, X.-X.; Jiang, J. Genome-wide identification, phylogeny, and expression analysis of the SWEET gene family in tomato. Gene 2015, 573, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, H.; Moser, M.; Klahre, U.; Esfeld, K.; Dell’Olivo, A.; Mandel, T.; Metzger, S.; Vandenbussche, M.; Freitas, L.; Kuhlemeier, C. MYB-FL Controls gain and loss of floral uv absorbance, a key trait affecting pollinator preference and reproductive isolation. Nat. Genet. 2016, 48, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedge, K.; Lohaus, G. Nectar sugars and amino acids in day- and night-flowering Nicotiana species are more strongly shaped by pollinators’ preferences than organic acids and inorganic ions. PLoS ONE 2017, 12, e0176865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colanero, S.; Perata, P.; Gonzali, S. What’s behind purple tomatoes? Insight into the mechanisms of anthocyanin synthesis in tomato fruits. Plant Physiol. 2020, 182, 1841–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laimbeer, F.P.E.; Bargmann, B.O.; Holt, S.H.; Pratt, T.; Peterson, B.; Doulis, A.G.; Buell, C.R.; Veilleux, R.E. Characterization of the F locus responsible for floral anthocyanin production in potato. G3 Genes Genomes Genet. 2020, 10, 3871–3879. [Google Scholar] [CrossRef] [PubMed]
- Hoballah, M.E.; Gübitz, T.; Stuurman, J.; Broger, L.; Barone, M.; Mandel, T.; Dell’Olivo, A.; Arnold, M.; Kuhlemeier, C. Single gene–mediated shift in pollinator attraction in Petunia. Plant Cell 2007, 19, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Esfeld, K.; Berardi, A.E.; Moser, M.; Bossolini, E.; Freitas, L.; Kuhlemeier, C. Pseudogenization and resurrection of a speciation gene. Curr. Biol. 2018, 28, 3776–3786.e7. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, E.W.; Landis, J.B.; Kurti, A.; Lawhorn, A.J.; Litt, A. The genetic basis of flower color differences in Nicotiana tabacum. In The Tobacco Plant Genome; Ivanov, N.V., Sierro, N., Peitsch, M.C., Eds.; Compendium of Plant Genomes; Springer International Publishing: Cham, Switzerland, 2020; pp. 175–193. [Google Scholar]
- Amrad, A.; Moser, M.; Mandel, T.; de Vries, M.; Schuurink, R.C.; Freitas, L.; Kuhlemeier, C. Gain and loss of floral scent production through changes in structural genes during pollinator-mediated speciation. Curr. Biol. 2016, 26, 3303–3312. [Google Scholar] [CrossRef]
- Christin, P.-A.; Weinreich, D.M.; Besnard, G. Causes and evolutionary significance of genetic convergence. Trends Genet. 2010, 26, 400–405. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Whittall, J.B.; Harder, K.D.; Barrett, S.C.H. Non-pollinator agents of selection on floral traits. In Ecology and Evolution of Flowers; Harder, L.D., Barrett, S.C.H., Eds.; Oxford University Press: Oxford, UK, 2006; pp. 120–138. [Google Scholar]
- Preston, J.C.; Hileman, L.C. Developmental genetics of floral symmetry evolution. Trends Plant Sci. 2009, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Rosin, F.M.; Kramer, E.M. Old dogs, new tricks: Regulatory evolution in conserved genetic modules leads to novel morphologies in plants. Dev. Biol. 2009, 332, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, R.M.; Money, D.; Ghatge, V.P.; Whelan, S.; Lovell, S.C. Determining the evolutionary history of gene families. Bioinformatics 2012, 28, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Env. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Penn, O.; Privman, E.; Landan, G.; Graur, D.; Pupko, T. An alignment confidence score capturing robustness to guide tree uncertainty. Mol. Biol. Evol. 2010, 27, 1759–1767. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Bayes empirical Bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Yang, Z. Multiple hypothesis testing to detect lineages under positive selection that affects only a few sites. Mol. Biol. Evol. 2007, 24, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Proctor, M.; Yeo, P.; Lack, A. The Natural History of Pollination; Harper & Collins: London, UK, 1996; 479p. [Google Scholar]
- Craig, J.M.; Kumar, S.; Hedges, S.B. Limitations of phylogenomic data can drive inferred speciation rate shifts. Mol. Biol. Evol. 2022, 39, msac038. [Google Scholar] [CrossRef] [PubMed]
- Greppi, J.A.; Hagiwara, J.C.; Stehmann, J.R. A new species of Petunia (Solanaceae) from Corrientes, Argentina. Phytotaxa 2019, 414, 289–295. [Google Scholar] [CrossRef]
- Kaunda, J.S.; Zhang, Y.-J. The genus Solanum: An ethnopharmacological, phytochemical and biological properties review. Nat. Prod. Bioprospecting 2019, 9, 77–137. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Chase, M.W.; Clarkson, J.J. Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). Taxon 2004, 53, 73–82. [Google Scholar] [CrossRef]
- Carrizo García, C.; Barfuss, M.H.J.; Sehr, E.M.; Barboza, G.E.; Samuel, R.; Moscone, E.A.; Ehrendorfer, F. Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Ann. Bot. 2016, 118, 35–51. [Google Scholar] [CrossRef]
- Reck-Kortmann, M.; Silva-Arias, G.A.; Segatto, A.L.A.; Mäder, G.; Bonatto, S.L.; de Freitas, L.B. Multilocus Phylogeny Reconstruction: New Insights into the Evolutionary History of the Genus Petunia. Mol. Phylogenet. Evol. 2014, 81, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.M.; Caballero-Villalobos, L.; Turchetto, C.; Assis Jacques, R.; Kuhlemeier, C.; Freitas, L.B. Do we truly understand pollination syndromes in Petunia as much as we suppose? AoB Plants 2018, 10, ply057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, S. On ‘various contrivances’: Pollination, phylogeny and flower form in the Solanaceae. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, G.; Sui, Y.; Wang, M.; Zhang, L. Pollinator responses to floral colour change, nectar and scent promote reproductive fitness in Quisqualis indica (Combretaceae). Sci. Rep. 2016, 6, 24408. [Google Scholar] [CrossRef]
- Fregonezi, J.N.; Turchetto, C.; Bonatto, S.L.; Freitas, L.B. Biogeographical history and diversification of Petunia and Calibrachoa (Solanaceae) in the Neotropical Pampas grassland. Bot. J. Linn. Soc. 2013, 171, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Gübitz, T.; Hoballah, M.E.; Dell’Olivo, A.; Kuhlemeier, C. Petunia as a model system for the genetics and evolution of pollination syndromes. In Petunia: Evolutionary, Developmental and Physiological Genetics; Gerats, T., Strommer, J., Eds.; Springer: New York, NY, USA, 2009; pp. 29–49. [Google Scholar]
- Huits, H.S.M.; Gerats, A.G.M.; Kreike, M.M.; Mol, J.N.M.; Koes, R.E. Genetic control of dihydroflavonol 4-reductase gene expression in Petunia hybrida. Plant J. 1994, 6, 295–310. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; Lian, H.; Chen, H. Molecular cloning and characterization of anthocyanin biosynthesis genes in eggplant (Solanum melongena L.). Acta Physiol. Plant. 2016, 38, 163. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, Q.; Li, W.; Guo, H. Cloning and bioinformatical analysis of dihydroflavonol 4-reductase gene from pigmented potato. Plant Gene Trait. 2012, 2, 7–14. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, E.W.; Berardi, A.E.; Smith, S.D.; Litt, A. Related allopolyploids display distinct floral pigment profiles and transgressive pigments. Am. J. Bot. 2017, 104, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Chase, M.W.; Paun, O.; Fay, M.F. Hybridization and speciation in angiosperms: A role for pollinator shifts? BMC Biol. 2010, 8, 45. [Google Scholar] [CrossRef]
- McCarthy, E.W.; Arnold, S.E.J.; Chittka, L.; Le Comber, S.C.; Verity, R.; Dodsworth, S.; Knapp, S.; Kelly, L.J.; Chase, M.W.; Baldwin, I.T.; et al. The effect of polyploidy and hybridization on the evolution of floral colour in Nicotiana (Solanaceae). Ann. Bot. 2015, 115, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanaik, S.; Kong, Q.; Zaitlin, D.; Werkman, J.R.; Xie, C.H.; Patra, B.; Yuan, L. Isolation and functional characterization of a floral tissue-specific R2R3 MYB regulator from tobacco. Planta 2010, 231, 1061–1076. [Google Scholar] [CrossRef] [PubMed]
- Mäder, G.; Freitas, L.B. Biogeographical, ecological, and phylogenetic analyses clarifying the evolutionary history of Calibrachoa in South American grasslands. Mol. Phylogenet. Evol. 2019, 141, 106614. [Google Scholar] [CrossRef] [PubMed]
- Wijsman, H.J.W. On the interrelationships of certain species of Petunia II. Experimental data: Crosses between different taxa. Acta Bot. Neerl. 1983, 32, 97–107. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Wing, J.; van der Woude, K.; Souer, E.; de Vetten, N.; Mol, J.; Koes, R. Molecular analysis of the Anthocyanin2 gene of Petunia and its role in the evolution of flower color. Plant Cell 1999, 11, 1433–1444. [Google Scholar] [CrossRef] [Green Version]
- Forkmann, G.; Ruhnau, B. Distinct substrate specificity of dihydroflavonol 4-reductase from flowers of Petunia hybrida. Z. Für Nat. C 1987, 42, 1146–1148. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.T.; Ryu, S.; Yi, H.; Shin, B.; Cheong, H.; Choi, G. Alteration of a single amino acid changes the substrate specificity of dihydroflavonol 4-reductase. Plant J. 2001, 25, 325–333. [Google Scholar] [CrossRef]
- Cronk, Q.; Ojeda, I. Bird-pollinated flowers in an evolutionary and molecular context. J. Exp. Bot. 2008, 59, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Lorenz-Lemke, A.P.; Mäder, G.; Muschner, V.C.; Stehmann, J.R.; Bonatto, S.L.; Salzano, F.M.; Freitas, L.B. Diversity and natural hybridization in a highly endemic species of Petunia (Solanaceae): A molecular and ecological analysis. Mol. Ecol. 2006, 15, 4487–4497. [Google Scholar] [CrossRef]
- Berardi, A.E.; Esfeld, K.; Jäggi, L.; Mandel, T.; Cannarozzi, G.M.; Kuhlemeier, C. Complex evolution of novel red floral color in Petunia. Plant Cell 2021, 33, 2273–2295. [Google Scholar] [CrossRef]
- Chen, Z.; Niu, Y.; Liu, C.-Q.; Sun, H. Red flowers differ in shades between pollination systems and across continents. Ann. Bot. 2020, 126, 837–848. [Google Scholar] [CrossRef] [PubMed]
- White, R.H.; Stevenson, R.D.; Bennett, R.R.; Cutler, D.E.; Haber, W.A. Wavelength discrimination and the role of ultraviolet vision in the feeding behavior of hawkmoths. Biotropica 1994, 26, 427. [Google Scholar] [CrossRef]
- Fenske, M.P.; Imaizumi, T. Circadian rhythms in floral scent emission. Front. Plant Sci. 2016, 7, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenske, M.P.; Nguyen, L.P.; Horn, E.K.; Riffell, J.A.; Imaizumi, T. Circadian clocks of both plants and pollinators influence flower seeking behavior of the pollinator hawkmoth Manduca sexta. Sci. Rep. 2018, 8, 2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colquhoun, T.A.; Marciniak, D.M.; Wedde, A.E.; Kim, J.Y.; Schwieterman, M.L.; Levin, L.A.; Van Moerkercke, A.; Schuurink, R.C.; Clark, D.G. A Peroxisomally localized acyl-activating enzyme is required for volatile benzenoid formation in a Petunia × hybrida Cv. ‘Mitchell diploid’ flower. J. Exp. Bot. 2012, 63, 4821–4833. [Google Scholar] [CrossRef] [Green Version]
- Yon, F.; Kessler, D.; Joo, Y.; Kim, S.-G.; Baldwin, I.T. Fitness consequences of a clock pollinator filter in Nicotiana attenuata flowers in nature. J. Integr. Plant Biol. 2017, 59, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Loughrin, J.N.; Hamilton-Kemp, T.R.; Andersen, R.A.; Hildebrand, D.F. Volatiles from flowers of Nicotiana sylvestris, N. otophora and Malus × domestica: Headspace components and day/night changes in their relative concentrations. Phytochemistry 1990, 29, 2473–2477. [Google Scholar] [CrossRef]
- Kessler, D.; Bing, J.; Haverkamp, A.; Baldwin, I.T. The defensive function of a pollinator-attracting floral volatile. Funct. Ecol. 2019, 33, 1223–1232. [Google Scholar] [CrossRef]
- Brandenburg, A.; Kuhlemeier, C.; Bshary, R. Hawkmoth pollinators decrease seed set of a low-nectar Petunia axillaris line through reduced probing time. Curr. Biol. 2012, 22, 1635–1639. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Gilet, T.; Bush, J.W.M. Optimal concentrations in nectar feeding. Proc. Natl. Acad. Sci. USA 2011, 108, 16618–16621. [Google Scholar] [CrossRef]
- Stuurman, J.; Hoballah, M.E.; Broger, L.; Moore, J.; Basten, C.; Kuhlemeier, C. Dissection of floral pollination syndromes in Petunia. Genetics 2004, 168, 1585–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, A.; Kevan, P.; Shipp, L.; Khosla, S.; McGarvey, B. The impact of greenhouse tomato (Solanales: Solanaceae) floral volatiles on bumble bee (Hymenoptera: Apidae) pollination. Environ. Entomol. 2012, 41, 855–864. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Younts, T.L.B.; Muday, G.K. Flavonols control pollen tube growth and integrity by regulating ROS homeostasis during high-temperature stress. Proc. Natl. Acad. Sci. USA 2018, 115, 11188–11197. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Han, Z.; Zhang, J.; Hu, Y.; Song, T.; Yao, Y. The balance of expression of dihydroflavonol 4-reductase and flavonol synthase regulates flavonoid biosynthesis and red foliage coloration in crabapples. Sci. Rep. 2015, 5, 12228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Chen, H.; Yang, R.; Ran, M. Identification of DFR as a promoter of anthocyanin accumulation in poinsettia (Euphorbia pulcherrima, Willd. Ex Klotzsch) bracts under short-day conditions. Sci. Hortic. 2018, 236, 158–165. [Google Scholar] [CrossRef]
- Shi, S.; Liu, Y.; He, Y.; Li, L.; Li, D.; Chen, H. R2R3-MYB transcription factor SmMYB75 promotes anthocyanin biosynthesis in eggplant (Solanum melongena L.). Sci. Hortic. 2021, 282, 110020. [Google Scholar] [CrossRef]
- Li, J.; Qin, M.; Qiao, X.; Cheng, Y.; Li, X.; Zhang, H.; Wu, J. A new insight into the evolution and functional divergence of SWEET transporters in chinese white pear (Pyrus bretschneideri). Plant Cell Physiol. 2017, 58, 839–850. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Ploidy | Copy Number | Pollinator | |||
|---|---|---|---|---|---|---|
| DFR | ODO1 | MYB-FL | NEC1 | |||
| Petunia axillaris | 2n | 1 | 1 | 1 | 1 | Ha |
| Petunia exserta | 2n | 1 | 1 | 1 | 1 | Hu |
| Petunia hybrida | 2n | 1 | 1 | 0 | 1 | B |
| Petunia inflata | 2n | 1 | 1 | 1 | 1 | B |
| Petunia secreta | 2n | 1 | 1 | 1 | 1 | B |
| Capsicum annum | 2n | 1 | 1 | 1 | 1 | B |
| Capsicum annum var. glabriusculum | 2n | 1 | 1 | 1 | 0 | B |
| Capsicum annuum var. zunla | 2n | 1 | 1 | 1 | 0 | B |
| Nicotiana attenuata | 2n | 1 | 1 | 2 | 1 | Ha/Hu |
| Nicotiana benthamiana | 4n | 2 | 2 | 1 | 2 | X |
| Nicotiana sylvestris | 2n | 1 | 1 | 1 | 1 | Ha |
| Nicotiana tabacum | 4n | 2 | 2 | 2 | 2 | Hu |
| Nicotiana tomentosiformis | 2n | 1 | 1 | 2 | 1 | B |
| Solanum lycopersicum | 2n | 1 | 1 | 1 | 1 | B |
| Solanum melogena | 2n | 1 | 1 | 0 | 1 | B |
| Solanum pennellii | 2n | 1 | 1 | 1 | 1 | B |
| Solanum pimpinellifolium | 2n | 1 | 1 | 1 | 1 | B |
| Solanum tuberosum | 2n | 1 | 1 | 1 | 1 | B |
| Arabidopsis thaliana | 2n | 1 | 2 | 1 | 1 | - |
| Total | 21 | 22 | 20 | 19 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, A.G.; Guzmán-Rodriguez, S.; Freitas, L.B. Phylogenetic Analyses of Some Key Genes Provide Information on Pollinator Attraction in Solanaceae. Genes 2022, 13, 2278. https://doi.org/10.3390/genes13122278
Pereira AG, Guzmán-Rodriguez S, Freitas LB. Phylogenetic Analyses of Some Key Genes Provide Information on Pollinator Attraction in Solanaceae. Genes. 2022; 13(12):2278. https://doi.org/10.3390/genes13122278
Chicago/Turabian StylePereira, Aléxia G., Sebastián Guzmán-Rodriguez, and Loreta B. Freitas. 2022. "Phylogenetic Analyses of Some Key Genes Provide Information on Pollinator Attraction in Solanaceae" Genes 13, no. 12: 2278. https://doi.org/10.3390/genes13122278
APA StylePereira, A. G., Guzmán-Rodriguez, S., & Freitas, L. B. (2022). Phylogenetic Analyses of Some Key Genes Provide Information on Pollinator Attraction in Solanaceae. Genes, 13(12), 2278. https://doi.org/10.3390/genes13122278