
Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.)

 ,
,  , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Materials
2.2. Field Evaluation
2.3. Statistical Analysis
2.3.1. Estimation of Genetic Components
2.3.2. Estimation of the Advantage over Better Parent and Commercial Variety
The Advantage over Better Parent
The Advantage over Commercial Variety
3. Results
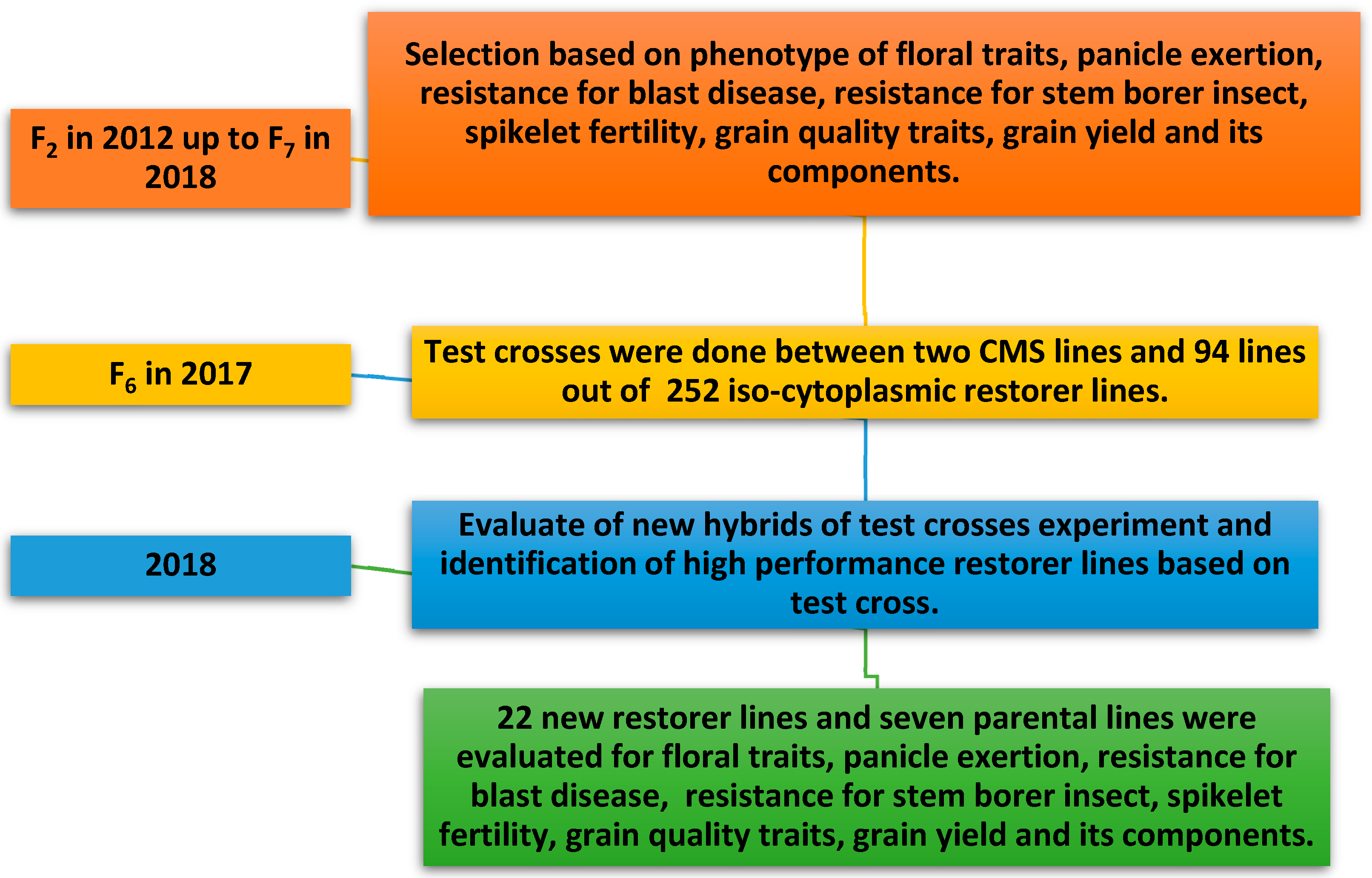
3.1. Developing New Iso-Cytoplasmic Restorers Lines
3.2. Test Cross Experiment
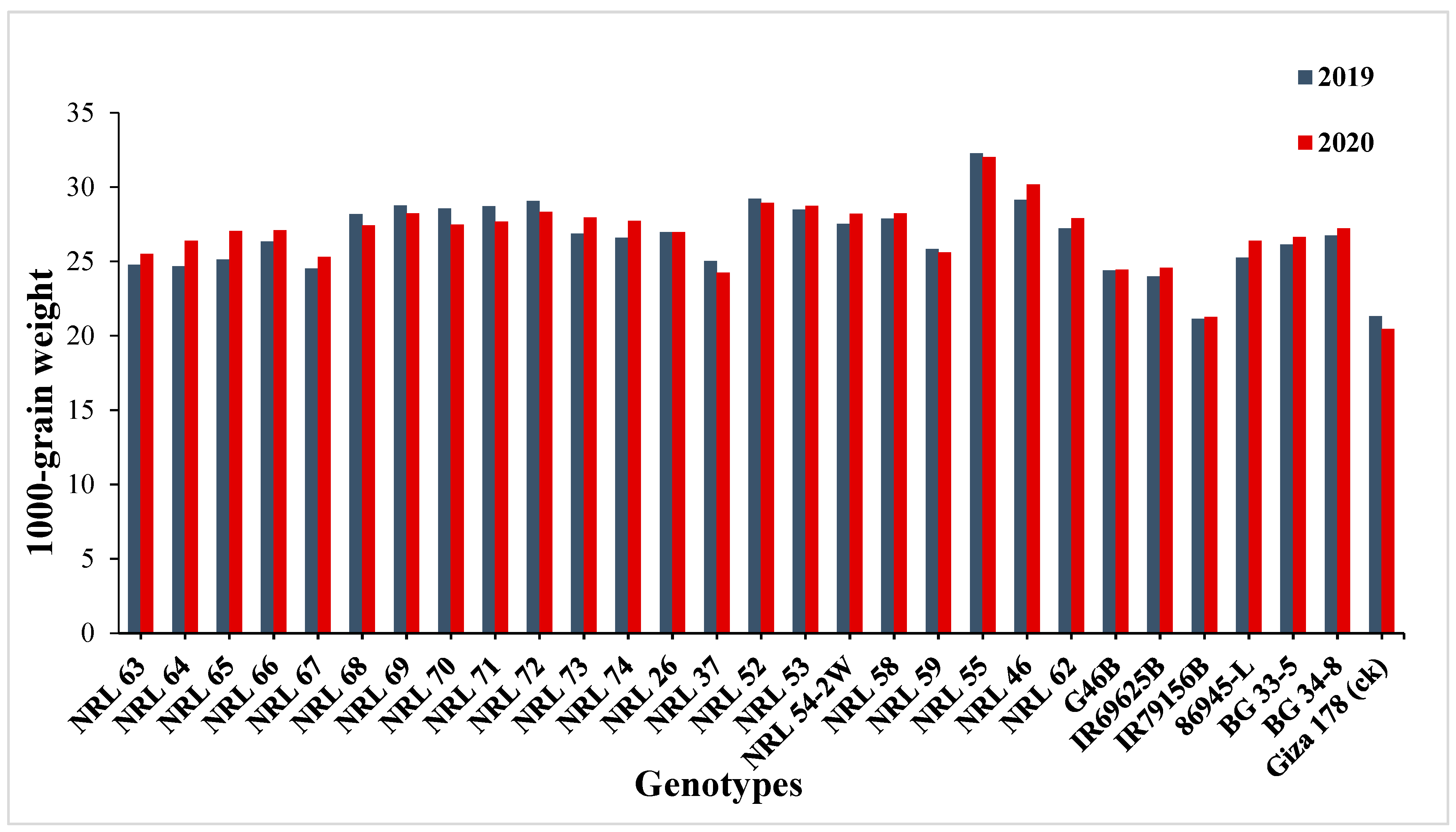
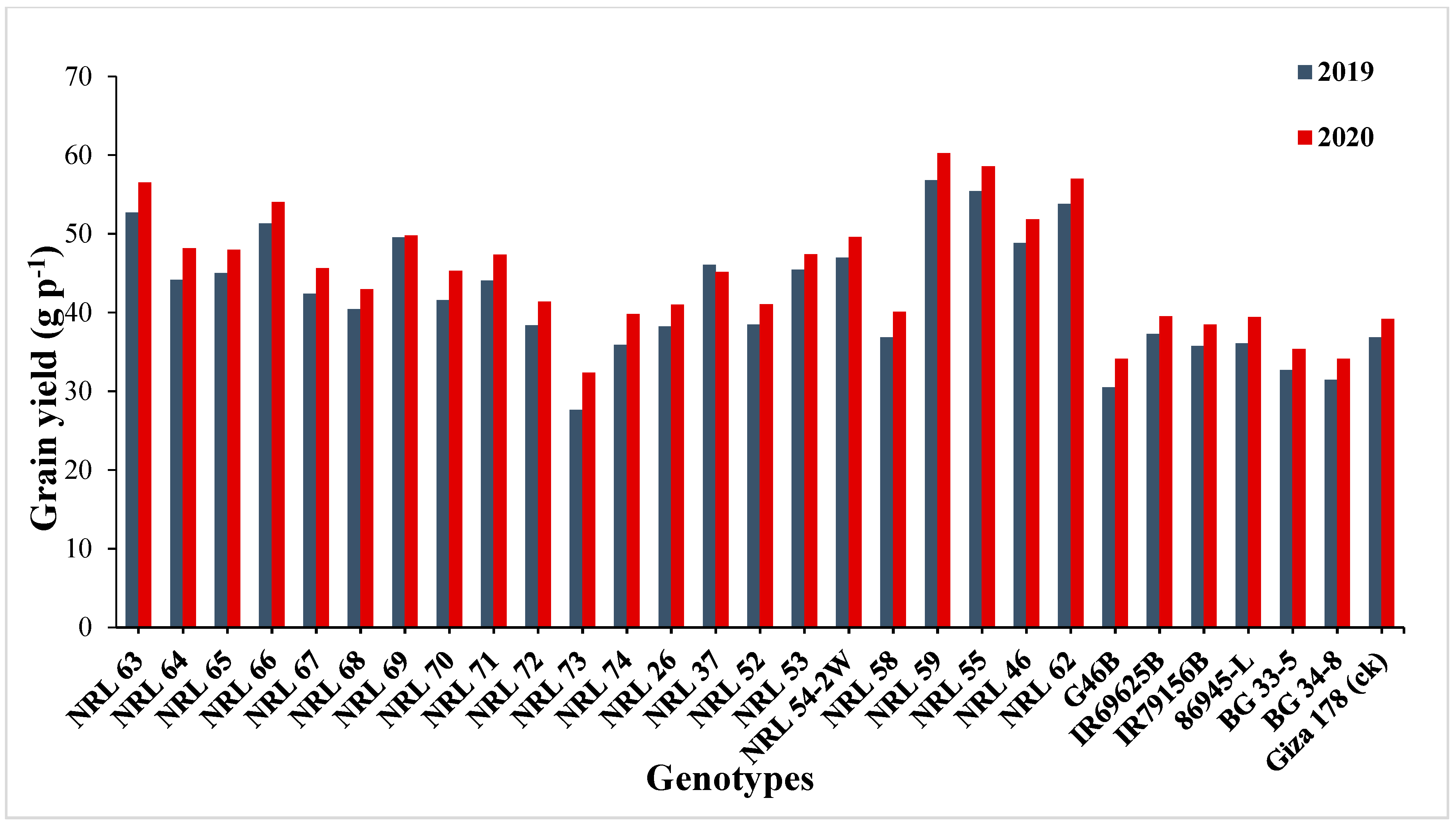
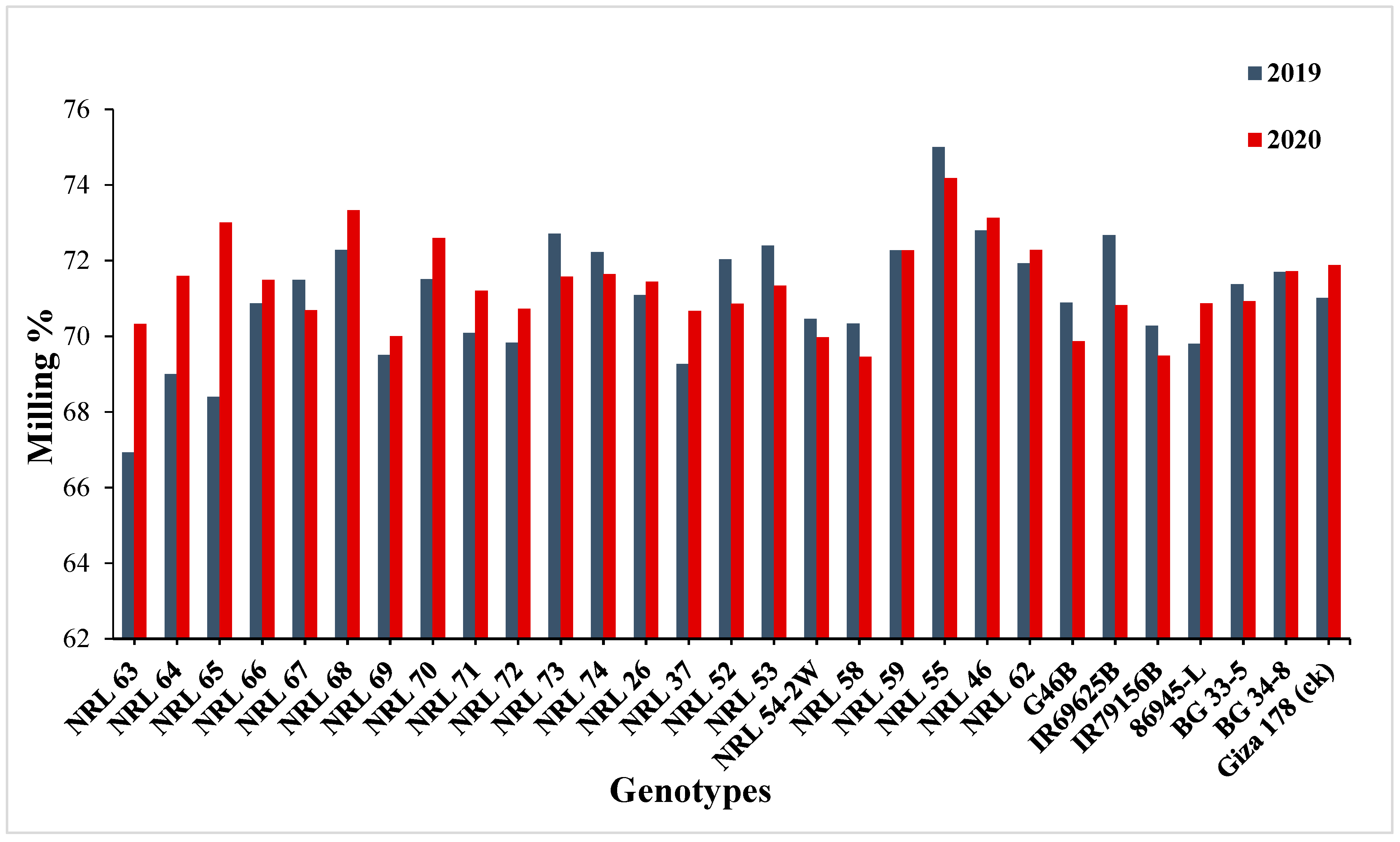
3.3. Mean Performance
3.4. Analysis of Variance
3.5. Phenotypic, Genotypic Coefficient of Variation, and Genetic Advance
3.6. Estimation of Heritability
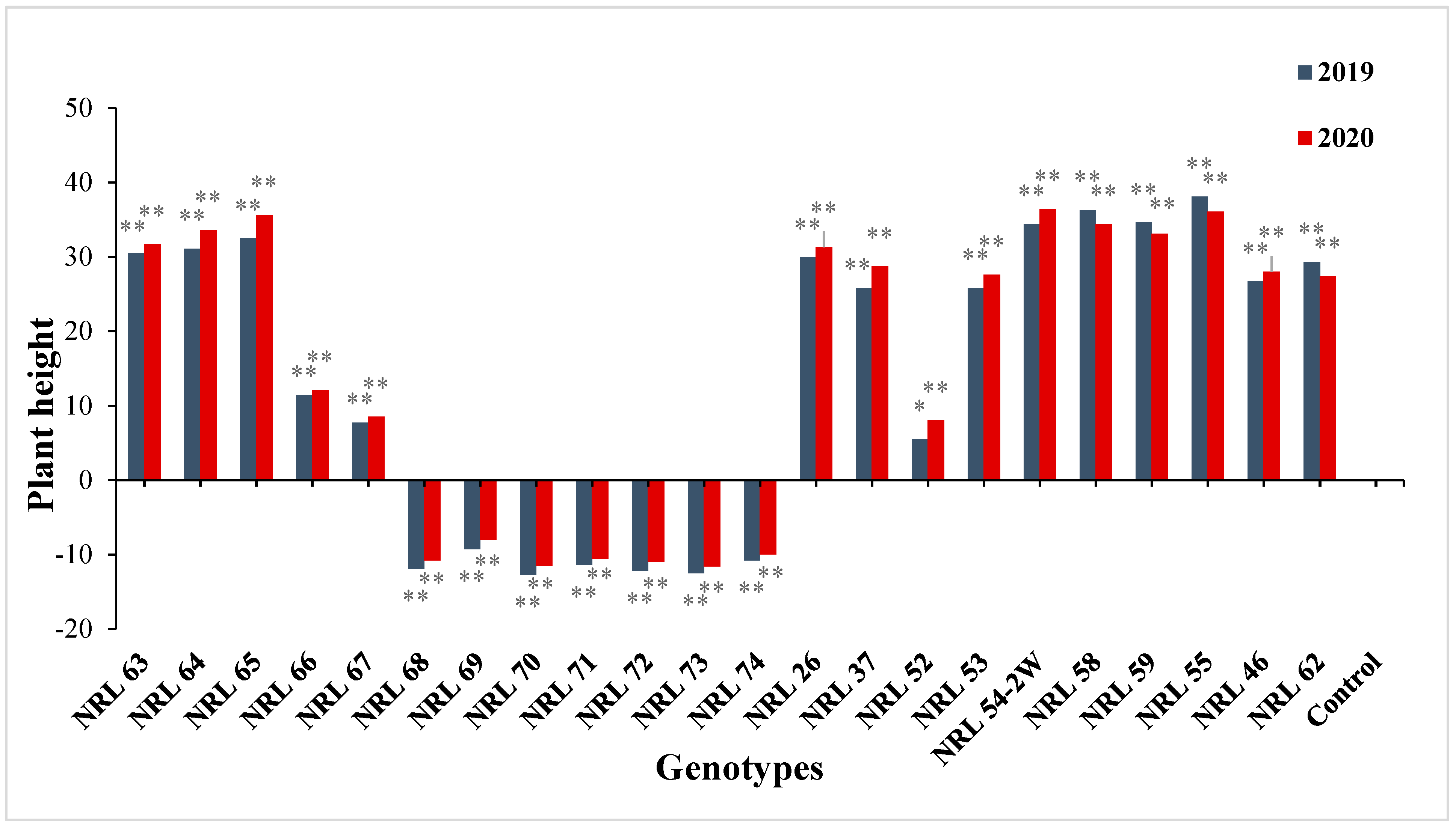
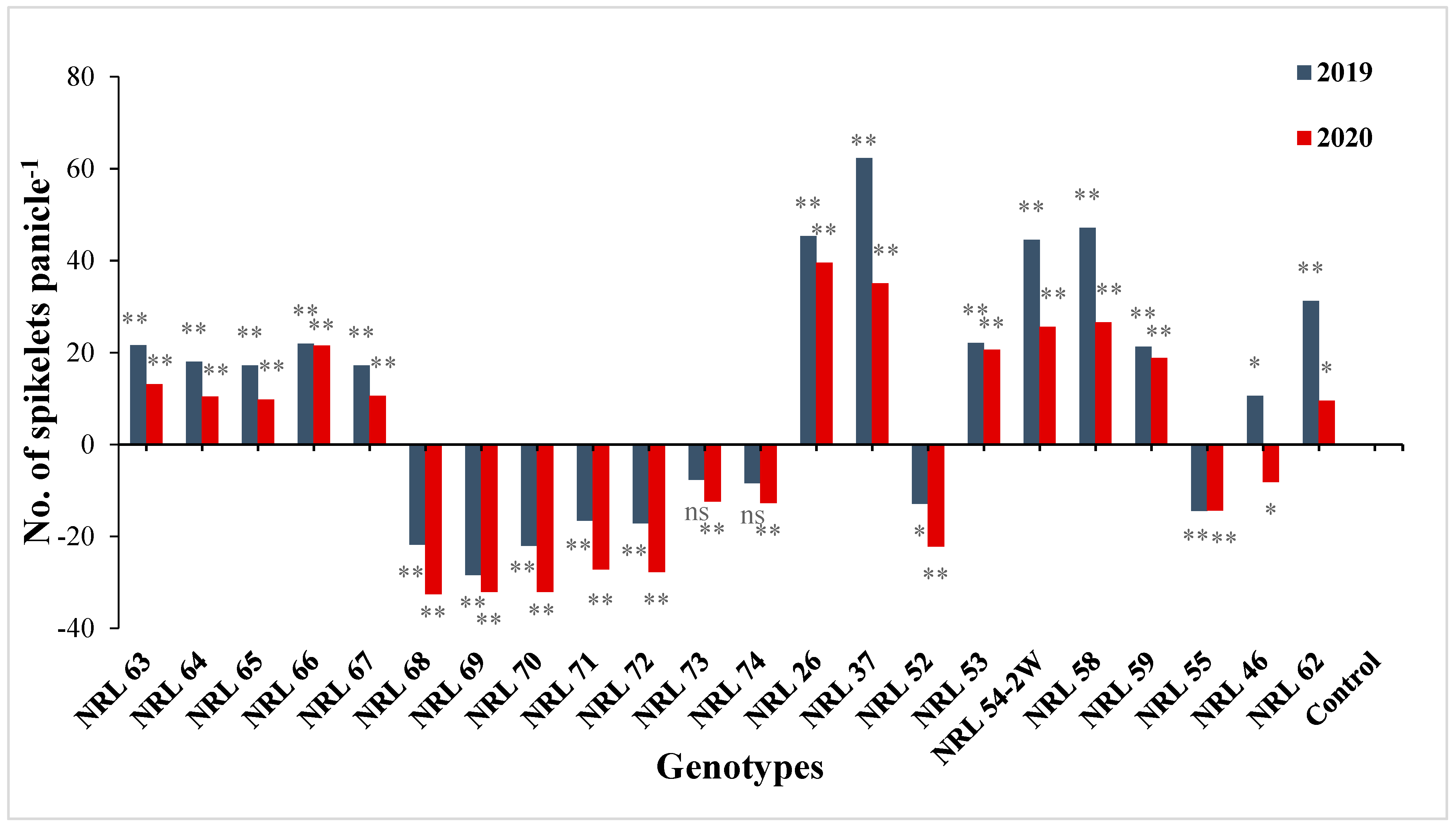
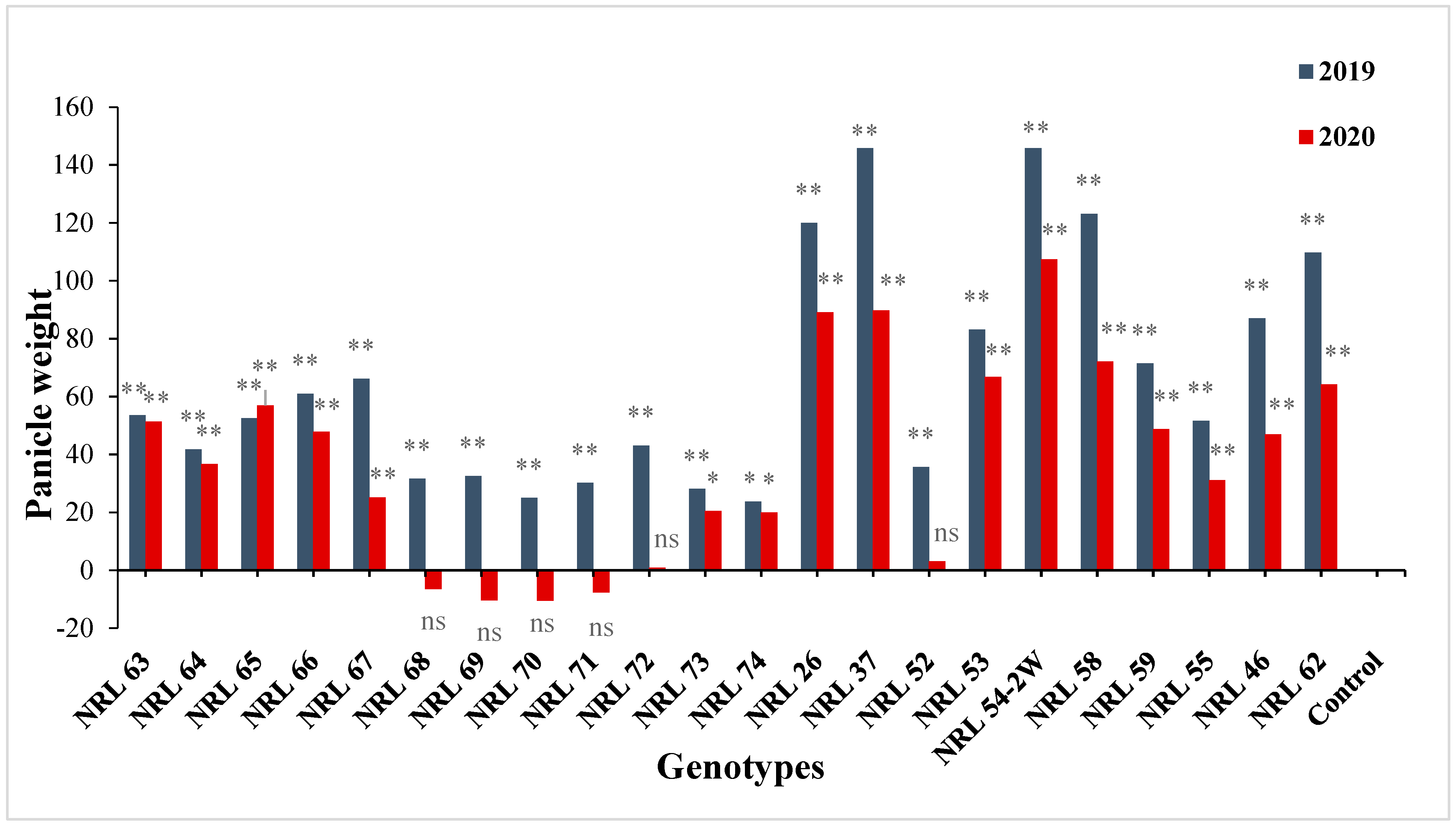
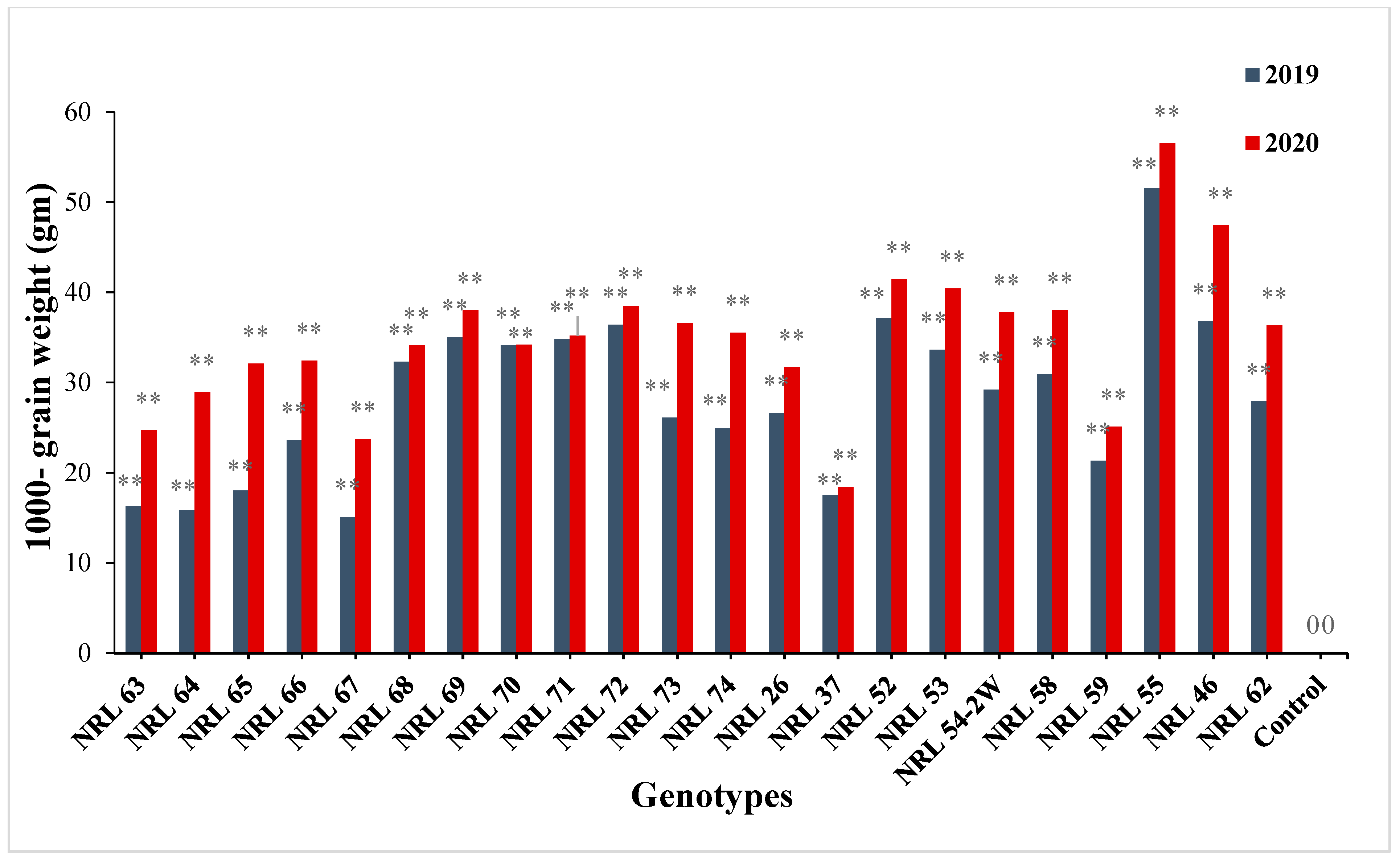
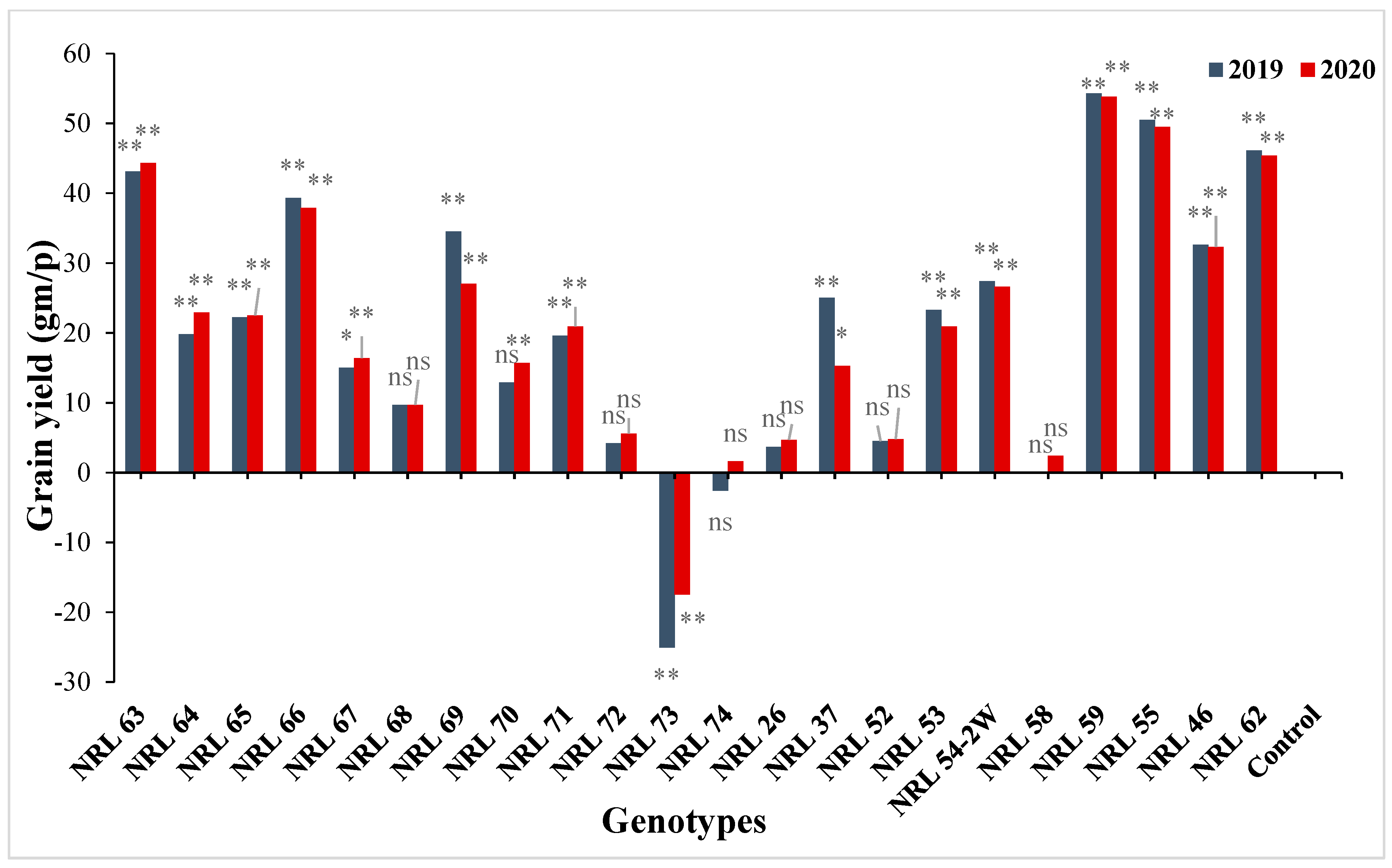
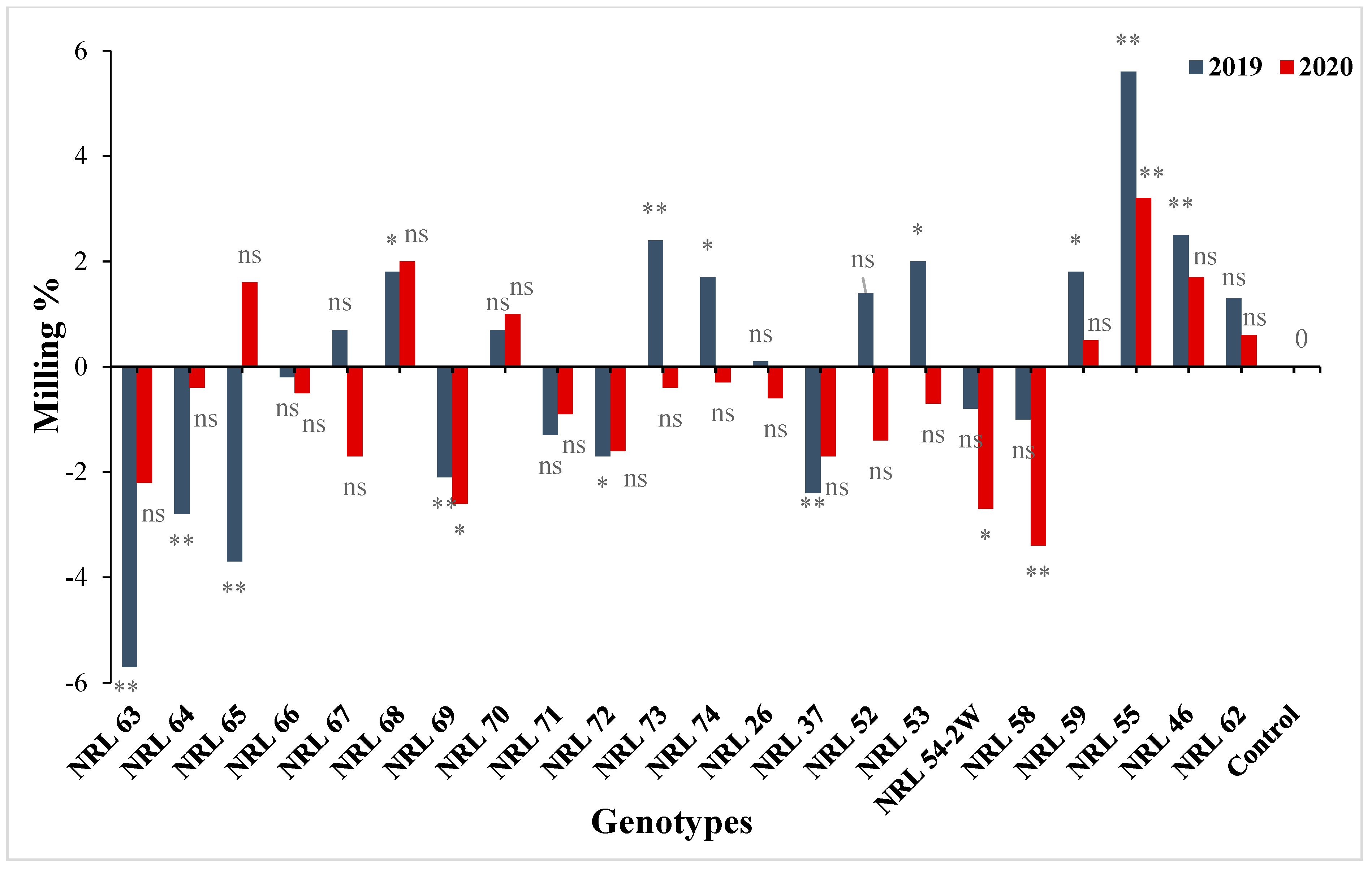
3.7. The Advantage over Better Parent and Commercial Variety
4. Discussion
4.1. Developing New Iso-Cytoplasmic Restorers Lines
4.2. Mean Performance
4.3. Test Cross Experiment
4.4. Analysis of Variance
4.5. Phenotypic, Genotypic Coefficient of Variation and Genetic Advance
4.6. Estimation of Heritability
4.7. The Advantage over Better Parent and Commercial Variety
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brarr, B.A.; Koehler, C.S.; Smith, R.F. Crop Losses, Rice: Field Losses to Insects, Diseases, Weeds, and Other Pests; University of California/USAID Pest Management and Related Environmental Protection Project Berkeley: Berkeley, CA, USA, 1975; p. 64. [Google Scholar]
- Al-Harbi, N.; Al Attar, N.; Hikal, D.; Mohamed, S.; Latef, A.A.; Ibrahim, A.; Abdein, M. Evaluation of Insecticidal Effects of Plants Essential Oils Extracted from Basil, Black Seeds and Lavender against Sitophilus oryzae. Plants 2021, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Ahmad, E.; Singh, D.; Singh, M.K. Identification of selection parameters for evaluating superior rice hybrids. Oryza-An Int. J. Rice 2018, 55, 230. [Google Scholar] [CrossRef]
- Zhu, Y. Fifty years of hybrid rice research in China. Chin. Sci. Bull. 2016, 61, 3740–3747. [Google Scholar] [CrossRef]
- Itabashi, E.; Iwata, N.; Fujii, S.; Kazama, T.; Toriyama, K. The fertility restorer gene, Rf2, for Lead Rice-type cytoplasmic male sterility of rice encodes a mitochondrial glycine-rich protein. Plant J. 2010, 65, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R. Interactions of Mitochondrial and Nuclear Genes That Affect Male Gametophyte Development. Plant Cell 2004, 16, S154–S169. [Google Scholar] [CrossRef]
- Fujii, S.; Toriyama, K. Genome Barriers between Nuclei and Mitochondria Exemplified by Cytoplasmic Male Sterility. Plant Cell Physiol. 2008, 49, 1484–1494. [Google Scholar] [CrossRef]
- Akagi, H.; Yokozeki, Y.; Inagaki, A.; Nakamura, A.; Fujimura, T. A codominant DNA marker closely linked to the rice nuclear restorer gene, Rf-1, identified with inter-SSR fingerprinting. Genome 1996, 39, 1205–1209. [Google Scholar] [CrossRef]
- Ichikawa, N.; Kishimoto, N.; Inagaki, A.; Nakamura, A.; Koshino, Y.; Yokozeki, Y.; Oka, M.; Samoto, S.; Akagi, H.; Higo, K.; et al. A rapid PCR-aided selection of a rice line containing the Rf-1 gene which is involved in restoration of the cytoplasmic male sterility. Mol. Breed. 1997, 3, 195–202. [Google Scholar] [CrossRef]
- Zhang, G.; Lu, Y.; Bharaj, T.S.; Virmani, S.S.; Huang, N. Mapping of the Rf-3 nuclear fertility-restoring gene for WA cytoplasmic male sterility in rice using RAPD and RFLP markers. Theor. Appl. Genet. 1997, 94, 27–33. [Google Scholar] [CrossRef]
- Yao, F.; Xu, C.; Yu, S.; Li, J.; Gao, Y.; Li, X.; Zhang, Q. Mapping and genetic analysis of two fertility restorer loci in the wild-abortive cytoplasmic male sterility system of rice (Oryza sativa L.). Euphytica 1997, 98, 183–187. [Google Scholar] [CrossRef]
- Shen, Y.W.; Guan, Z.Q.; Lu, J.; Zhuang, J.Y.; Zheng, K.L.; Gao, M.W.; Wang, X.M. Linkage analysis of a fertility restoring mutant generated from CMS rice. Theor. Appl. Genet. 1998, 97, 261–266. [Google Scholar] [CrossRef]
- Bharaj, T.S.; Virmani, S.S.; Khush, G.S. Chromosomal location of fertility restoring genes for ‘wild abortive’ cytoplasmic male sterility using primary trisomics in rice. Euphytica 1995, 83, 169–173. [Google Scholar] [CrossRef]
- Tan, X.L.; Vanavichit, A.; Amornsilpa, S.; Trangoonrung, S. Genetic analysis of rice CMS-WA fertility restoration based on QTL mapping. Theor. Appl. Genet. 1998, 97, 994–999. [Google Scholar] [CrossRef]
- Zhu, L.; Lu, C.; Li, P.; Shen, L.; Xu, Y.; He, P.; Chen, Y. Using doubled haploid populations of rice for quantitative trait locus mapping. Rice Genet. Collect. 2008, 3, 631–636. [Google Scholar] [CrossRef]
- Huang, Q.; He, Y.; Jin, R.; Huang, H.; Zhu, Y. Tagging of the restorer gene for rice HL-type CMS using microsatellite markers. Rice Genet. Newsl. 1999, 16, 75–76. [Google Scholar]
- Awad-Allah, M.M.A. Application of Genetic Engineering Tools on Rice Genome. Master’s Thesis, Al-Azhar University Faculty of Agriculture, Cairo, Egypt, 2006. [Google Scholar]
- Awad-Allah, M.M.A. Integrated Restorer Fertility and Wide Compatibility Genes for Producing Promising Restorer Lines in Rice. Ph.D. Thesis, Al-Azhar University Faculty of Agriculture, Cairo, Egypt, 2011. [Google Scholar]
- Ren, G.; Yan, L.; Xie, H. Retrospective and perspective on indica three-line hybrid rice breeding research in China. Chin. Sci. Bull. 2016, 61, 3748–3760. [Google Scholar] [CrossRef]
- Liao, C.; Yan, W.; Chen, Z.; Xie, G.; Deng, X.W.; Tang, X. Innovation and development of the third-generation hybrid rice technology. Crop J. 2021, 9, 693–701. [Google Scholar] [CrossRef]
- Dai, Z.; Lu, Q.; Luan, X.; Ouyang, L.; Guo, J.; Liang, J.; Zhu, H.; Wang, W.; Wang, S.; Zeng, R.; et al. Development of a platform for breeding by design of CMS restorer lines based on an SSSL library in rice (Oryza sativa L.). Breed. Sci. 2016, 66, 768–775. [Google Scholar] [CrossRef]
- Sabar, M.; Akthar, M. Rice germplasm collection, conservation and utilization for hybrid rice. In Proceedings of the Seminar on Sustainable Utilization of Plant Genetic Resources for Agricultural Production, Islamabad, Pakistan, 17–19 December 2002; pp. 55–61. [Google Scholar]
- Kumar, B.P.; Bhowmick, P.K.; Singh, V.J.; Malik, M.; Gupta, A.K.; Seth, R.; Nagarajan, M.; Krishnan, S.G.; Singh, A.K. Marker-assisted identification of restorer gene(s) in iso-cytoplasmic restorer lines of WA cytoplasm in rice and assessment of their fertility restoration potential across environments. Physiol. Mol. Biol. Plants 2017, 23, 891–909. [Google Scholar] [CrossRef]
- Kumar, A.; Bhowmick, P.K.; Krishnan, S.G.; Singh, A.K. Development and evaluation of iso-cytoplasmic rice restorer lines for different agro-morphological traits. Indian J. Genet. Plant Breed. 2017, 77, 493. [Google Scholar] [CrossRef]
- Hashim, S.; Ding, P.; Ismail, M.F.; Ramli, A. Floral traits and flowering behaviors of Malaysian rice cytoplasmic male sterile and maintainer lines and its relationship with out-crossing performance. Aust. J. Crop Sci. 2021, 15, 180–186. [Google Scholar] [CrossRef]
- Yang, G.P.; Maroof, M.S.; Xu, C.G.; Zhang, Q.; Biyashev, R.M. Comparative analysis of microsatellite DNA polymorphism in landraces and cultivars of rice. Mol. Genet. Genom. 1994, 245, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Watabe, T.; Nishioka, R.; Thanh, P.T.; Ishii, T. Identification of Quantitative Trait Loci Controlling Floral Morphology of Rice Using a Backcross Population between Common Cultivated Rice, Oryza sativa and Asian Wild Rice, O. rufipogon. Am. J. Plant Sci. 2017, 8, 734–744. [Google Scholar] [CrossRef]
- Appalaswamy, A.; Reddy, G.L.K. Genetic divergence and heterosis studies of mungbean (Vigna radiata L.) Wilczek. Legume Res. 2004, 21, 115–118. [Google Scholar]
- Alam, S.; Tester, M.; Fiene, G.; Mousa, M. Early Growth Stage Characterization and the Biochemical Responses for Salinity Stress in Tomato. Plants 2021, 10, 712. [Google Scholar] [CrossRef]
- International Rice Research Institute (IRRI). Standard Evaluation System for Rice, 5th ed.; International Rice Research Institute (IRRI): Los Banos, PA, USA, 2014; p. 57. ISBN 9789712203046. [Google Scholar]
- Xangsayasane, P.; Xie, F.; Borromeo, T.H. Borromeo. Grain quality evaluation of Lao high yielding cultivars. Lao J. Agric. For. 2009, 19, 30–45. [Google Scholar]
- Panse, V.G.; Sukhatme, P.V. Statistical Methods for Agricultural Workers; ICAR: New Delhi, India, 1961; p. 361. [Google Scholar]
- Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis. Biom. Methods Quant. Genet. Anal. 1977, 34, 723. [Google Scholar] [CrossRef]
- Falconer, D.S. Introduction to Qualitative Genetics, 2nd ed.; Longman: London, UK, 1989; p. 340. [Google Scholar]
- Burton, G.W. Qualitative inheritance in grasses. In Proceedings of the 6th International Grassland Congress, State College, PA, USA, 17–23 August 1952. [Google Scholar]
- Lush, J.L. Intrasire correlations or regressions of offspring on dam as a method of estimating heritability of characteristics. Proc. Am. Soc. Anim. Nutr. 1940, 1, 293–301. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of Genetic and Environmental Variability in Soybeans 1. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Sivasubramanian, S.; Madhavamenon, P. Genotypic and phenotypic variability in rice. Madras Agric. J. 1973, 60, 1093–1096. [Google Scholar]
- Mather, K. Biometrical Genetics, 3rd ed.; Cambridge University Press: London, UK, 1949; 158p. [Google Scholar]
- Wynne, J.C.; Emery, D.A.; Rice, P.W. Combining Ability Estimates in Arachis hypogaea L. II. Field Performance of F1 Hybrids 1. Crop Sci. 1970, 10, 713–715. [Google Scholar] [CrossRef]
- Gangashetti, M.G.; Jena, K.K.; Shenoy, V.V.; Freeman, W.H. Inheritance of elongated uppermost internode and identification of RAPD marker linked to eui gene in rice. Curr. Sci. 2004, 87, 469–475. [Google Scholar]
- Nachimuthu, V.V. Evaluation of Rice Genetic Diversity and Variability in a Population Panel by Principal Component Analysis. Indian J. Sci. Technol. 2014, 7, 1555–1562. [Google Scholar] [CrossRef]
- Mondo, V.H.V.; Nascente, A.S.; Neves, P.D.C.F.; Taillebois, J.E.; Neto, M.O.A.C.; Développement, B.C. Flowering synchronization in hybrid rice parental lines. Aust. J. Crop Sci. 2016, 10, 1213–1221. [Google Scholar] [CrossRef]
- Sindhu, J.S.; Kumar, I. Quality seed production in hybrid rice. In Proceedings of the 20th Session of International Rice Commis-sion, (IRC’ 02), Bangkok, Thailand, 23–26 July 2002. [Google Scholar]
- Vishwakarma, A.; Singh, J.K.; Singh, R.K.; Jat, A.L. Productivity and profitability of rice (Oryza sativa) hybrids as influenced by date of transplanting and age of seedling under system of rice intensification. Ind. J. Agron. 2016, 61, 315–320. [Google Scholar]
- Bhuiyan, M.S.H.; Zahan, A.; Khatun, H.; Iqbal, M.; Alam, F.; Manir, M.R. Yield performance of newly devel-oped test crossed hybrid rice variety. IJAAR 2014, 5, 48–54. [Google Scholar]
- Mahalingam, A.; Saraswathi, R.; Ramalingam, J.; Jayaraj, T. Genetics of floral traits in cytoplasmic male sterile (CMS) and restorer lines of hybrid rice (Oryza sativa L.). Pak. J. Bot. 2013, 45, 1897–1904. [Google Scholar]
- Priyanka, R.M.; Thiyagarajan, K.; Rajendran, S.P.B.R. Identification of restorers and maintainers among the locally adapted genotypes for hybrid development in rice (Oryza sativa L.). Electron. J. Plant Breed. 2016, 7, 814. [Google Scholar] [CrossRef]
- Joshi, B.K.; Subedi, L.P.; Gurung, S.B.; Sharma, R.C. Pollen and Spikelet Analysis in F1 Rice Hybrids and their Parents. Nepal. Agric. Res. J. 2014, 8, 120–126. [Google Scholar] [CrossRef]
- Awad-Allah, M.M.A. Genetical analysis and detection for wide compatibility traits of some rice genotypes under egyptian conditions. J. Sustain. Agric. Sci. 2016, 42, 99–125. [Google Scholar] [CrossRef]
- Toriyama, K.; Kazama, T.; Sato, T.; Fukuta, Y.; Oka, M. Development of cytoplasmic male sterile lines and restorer lines of various elite Indica Group rice cultivars using CW-CMS/Rf17 system. Rice 2019, 12, 73. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.R.; Venkanna, V.; Parimala, K.; Hari, Y.; Satish Chandra, B.; Lingaiah, N.; Prasad, K.R.; Rao, P.J.M. Developing of Heterotic Hybrids through Allocytoplasmic Restorer Improvement In Rice (Oryza sativa L.). In Proceedings of the 1st Indian Rice Congress—2020. Rice Research and Development for Achieving Sustainable Development Goals, Cuttack, India, 8–9 December 2020; ICAR-NRRI: Cuttack, India, 2020; pp. 27–28. [Google Scholar]
- Prasad, K.R.K.; Suneetha, Y. Fertility Restoration Studies of Promising Genotypes in Rice (Oryza sativa L.). In Proceedings of the 1st Indian Rice Congress—2020. Rice Research and Development for Achieving Sustainable Development Goals, Cuttack, India, 8–9 December 2020; ICAR-NRRI: Cuttack, India, 2020; pp. 40–42. [Google Scholar]
- Roy, A.K.; Miha, M.A.K.; Nath, B.C.; Ghosal, S.; Biswas, P.L. Genetic parameters and correlation Among Floral Characters in Local Fine Rice. J. Sci. Technol. 2008, 6, 75–79. [Google Scholar]
- Priyanka, R.M.; Thiyagarajan, K.; Bharathi, S.P.; Rajendran, R. Studies on genetic variability for floral and grain quality traits in rice (Oryza sativa L.). Electron. J. Plant Breed. 2017, 8, 609. [Google Scholar] [CrossRef]
- Radha, T.; Satheesh Kumar, P.; Saravanan, K.; Suganthi, S. Genetic variabilty, association and path analysis for grain yield and quality traits in Rice (Oryza sativa L.). Plant Arch. 2019, 19, 2522–2526. [Google Scholar]
- Adjah, K.L.; Abe, A.; Adetimirin, V.O.; Asante, M.D. Genetic variability, heritability and correlations for milling and grain appearance qualities in some accessions of rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2020, 26, 1309–1317. [Google Scholar] [CrossRef]
- Genetic variability and association analysis in rice (Oryza sativa L.) germplasm based on biometrical and floral traits. Electron. J. Plant Breed. 2021, 12, 912–917. [CrossRef]
- Salimath, P.M.; Shashidhar, H.E.; Mohankumar, H.D.; Patil, S.S.; Vamadevaiah, H.M.; Janagoudar, B.S. Analysis of genetic variability in interspesific backcross inbred lines in rice (Oryza sativa L.). Kanataka J. Agric. Sci. 2010, 23, 563–565. [Google Scholar]
- Prajapati, M.K.; Singh, C.M.; Suresh, B.G.; Lavanya, G.R.; Jadav, P. Genetic parameters for grain yield and its component characters in rice (Oryza sativa L.). Electron. J. Plant Breed. 2011, 2, 235–238. [Google Scholar]
- Venkatesan, M.; Elangaimannan, R.; Thilagavathi, T. Quantitative Analysis of Rice (Oryza sativa L.). Plant Arch. 2019, 19, 1027–1031. [Google Scholar]
- Behera, P.P.; Singh, S.K.; Singh, D.K.; Longkho, K. Genetic Parameters Study for Yield and Yield Contributing Characters in Rice (Oryza sativa L.) Genotypes with High Grain Zinc Content. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 357–364. [Google Scholar] [CrossRef]
- Singh, B.; Singh, R. Floral variability and correlation studies among selected male sterile lines in rice. J. Biotechnol. Crop Sci. 2013, 2, 34–36. [Google Scholar]
- Buelah, J.; Reddy, V.R.; Srinivas, B.; Balram, N. Studies on Combining Ability and Gene Action for Yield and Quality Traits in Hybrid Rice (Oryza sativa L.). Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1282–1290. [Google Scholar] [CrossRef]
- Sankar, P.D.; Sheeba, A.; Anbumalarmathi, J. Variability and character association of cultivars of rice (Oryza sativa L.). Agric. Sci. Digest 2006, 26, 182–184. [Google Scholar]
- Islam, M.; Raffi, S.; Hossain, M.; Hasan, A. Analysis of genetic variability, heritability and genetic advance for yield and yield associated traits in some promising advanced lines of rice. Progress. Agric. 2015, 26, 26–31. [Google Scholar] [CrossRef]
- Awad-Allah, M.M.A. Heterosis and Combining ability Estimates using Line x Tester Analysis to Develop Wide Compatibility and Restorer Lines in Rice. J. Agric. Chem. Biotechnol. 2020, 11, 383–393. [Google Scholar] [CrossRef]
- Yadav, A.K.; Vyas, R.P.; Yadav, V.K.; Kumar, V. Exploitation of heterobeltiosis and economic heterosis for yield and its component traits in rice (Oryza sativa L.). Plant Arch. 2020, 20, 4555–4563. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Lines | Parentage | Description |
|---|---|---|---|
| 1 | NRL 63 | IR79156A/86945-L | New developed restorer |
| 2 | NRL 64 | IR79156A/86945-L | New developed restorer |
| 3 | NRL 65 | IR79156A/86945-L | New developed restorer |
| 4 | NRL 66 | G46A/Giza 178 | New developed restorer |
| 5 | NRL 67 | G46A/Giza 178 | New developed restorer |
| 6 | NRL 68 | G46A/BG 33-5 | New developed restorer |
| 7 | NRL 69 | G46A/BG 33-5 | New developed restorer |
| 8 | NRL 70 | G46A/BG 33-5 | New developed restorer |
| 9 | NRL 71 | G46A/BG 33-5 | New developed restorer |
| 10 | NRL 72 | G46A/BG 33-5 | New developed restorer |
| 11 | NRL 73 | G46A/BG 33-5 | New developed restorer |
| 12 | NRL 74 | G46A/BG 33-5 | New developed restorer |
| 13 | NRL 26 | G46A/BG 33-5 | New developed restorer |
| 14 | NRL 37 | G46A/BG 33-5 | New developed restorer |
| 15 | NRL 52 | G46A/86945-L | New developed restorer |
| 16 | NRL 53 | G46A/86945-L | New developed restorer |
| 17 | NRL 54-2 | G46A/BG 34-8 | New developed restorer |
| 18 | NRL 58 | G46A/BG 34-8 | New developed restorer |
| 19 | NRL 59 | G46A/BG 34-8 | New developed restorer |
| 20 | NRL 55 | IR69625A/BG 34-8 | New developed restorer |
| 21 | NRL 46 | IR69625A/BG 34-8 | New developed restorer |
| 22 | NRL 62 | IR69625A/BG 34-8 | New developed restorer |
| 23 | G46A, B | Erjiu’ai 7/V41B//Zhenshan 97/Ya’aizao | cytoplasmic male sterile and its maintainer line |
| 24 | IR69625A, B | not available | cytoplasmic male sterile and its maintainer line |
| 25 | IR79156A, B | not available | cytoplasmic male sterile and its maintainer line |
| 26 | 86945-L | not available | Restorer line 1 |
| 27 | BG 33-5 | not available | Restorer line 1, drought tolerant high grain quality |
| 28 | BG 34-8 | not available | Restorer line 1, drought tolerant high grain quality |
| 29 | Giza 178 (ck) | Giza 175/Milyang 49 | Commercial Restorer line, |
| Mean Sum of Squares | CV | |||||||
|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | |||||||
| Rep. | Genotype | Error | Rep | Genotype | Error | 2019 | 2020 | |
| d.f | 2 | 28 | 56 | 2 | 28 | 56 | ||
| days to 50% of heading (day) | 0.10 | 248.64 ** | 0.49 | 0.87 | 246.99 ** | 0.50 | 0.736 | 0.737 |
| plant height (cm) | 7.99 | 820.95 ** | 6.53 | 13.35 | 805.15 ** | 10.33 | 2.52 | 3.05 |
| number of panicles /plant | 1.83 | 19.30 ** | 2.81 | 2.21 | 12.44 ** | 2.58 | 10.05 | 9.47 |
| panicle length (cm) | 0.09 | 16.60 ** | 0.55 | 1.12 | 17.60 ** | 0.67 | 3.22 | 3.50 |
| number of spikelets /panicle | 193.31 | 3979.85 ** | 77.79 | 109.21 | 4078.95 ** | 57.38 | 5.20 | 4.47 |
| spikelet fertility percentage | 5.05 | 19.09 ** | 2.87 | 1.57 | 12.01 ** | 1.25 | 1.85 | 1.2 |
| number of filled grains per panicle | 81.26 | 3290.61 ** | 73.46 | 103.89 | 3442.09 ** | 49.37 | 5.53 | 4.47 |
| panicle weight (g) | 0.02 | 3.50 ** | 0.11 | 0.11 | 3.85 ** | 0.14 | 7.58 | 8.39 |
| 1000-grain weight | 0.20 | 17.34 ** | 0.24 | 0.08 | 16.51 ** | 0.18 | 1.84 | 1.57 |
| grain yield/plant | 11.30 | 180.31 ** | 10.46 | 0.47 | 173.85 ** | 8.37 | 7.69 | 6.44 |
| Anther length (mm) | 0.08 | 0.51 ** | 0.04 | 0.07 | 0.52 ** | 0.04 | 9.05 | 9.13 |
| Anther breadth (mm) | 0.0003 | 0.015 ** | 0.0004 | 0.0001 | 0.014 ** | 0.0003 | 4.59 | 4.1 |
| Filament length (mm) | 0.34 | 1.24 ** | 0.16 | 0.37 | 1.44 ** | 0.16 | 5.98 | 6.04 |
| Duration of floret opening (min) | 15.998 | 1292.18 ** | 12.67 | 12.09 | 1291.53 ** | 9.71 | 2.74 | 2.41 |
| Hulling (%) | 0.88 | 3.92 ** | 0.47 | 0.58 | 3.28 ** | 0.62 | 0.85 | 0.98 |
| Milling (%) | 0.91 | 7.73 ** | 0.50 | 0.73 | 4.08 ** | 1.06 | 1.00 | 1.44 |
| Head rice % | 3.98 | 262.29 ** | 2.68 | 25.79 | 251.13 ** | 9.11 | 2.82 | 5.17 |
| GCV% | PCV% | GA | Heritability % | GA% | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| days to 50% of heading (day) | 9.54 | 9.48 | 9.57 | 9.51 | 18.68 | 18.62 | 99.41 | 99.40 | 19.60 | 19.47 |
| plant height (cm) | 15.79 | 15.51 | 15.98 | 15.81 | 33.54 | 32.90 | 97.65 | 96.25 | 32.15 | 31.35 |
| number of panicles per plant | 14.06 | 10.70 | 17.28 | 14.29 | 3.93 | 2.80 | 66.19 | 10.70 | 23.56 | 16.51 |
| panicle length (cm) | 10.03 | 10.13 | 10.54 | 10.71 | 4.54 | 4.62 | 90.64 | 89.33 | 19.67 | 19.71 |
| number of spikelets per panicle | 21.27 | 21.62 | 21.90 | 22.08 | 72.17 | 73.86 | 94.36 | 95.90 | 42.57 | 43.62 |
| spikelet fertility percentage | 2.54 | 2.04 | 3.15 | 2.37 | 3.87 | 3.36 | 65.29 | 74.22 | 4.23 | 3.62 |
| number of filled grains per panicle | 21.14 | 21.41 | 21.85 | 21.87 | 65.26 | 67.81 | 93.59 | 95.82 | 42.13 | 43.16 |
| panicle weight (g) | 23.84 | 24.53 | 25.02 | 25.92 | 2.09 | 2.17 | 90.82 | 89.53 | 46.81 | 47.80 |
| 1000-grain weight (g) | 8.98 | 8.70 | 9.17 | 8.84 | 4.82 | 4.73 | 95.98 | 96.85 | 18.13 | 17.63 |
| grain yield/plant | 17.88 | 16.53 | 19.46 | 17.74 | 14.24 | 14.26 | 84.41 | 86.82 | 33.84 | 31.73 |
| Anther length (mm) | 17.61 | 17.92 | 19.80 | 20.11 | 0.73 | 0.73 | 79.13 | 79.37 | 32.27 | 32.88 |
| Anther breadth (mm) | 15.59 | 15.05 | 16.18 | 15.59 | 0.14 | 0.14 | 92.84 | 93.1 | 30.93 | 29.9 |
| Filament length (mm) | 9.02 | 9.87 | 10.83 | 11.57 | 1.03 | 1.15 | 69.46 | 72.74 | 15.49 | 17.34 |
| Duration of floret opening (min) | 15.87 | 15.95 | 16.10 | 16.13 | 41.92 | 42.11 | 97.11 | 97.78 | 32.21 | 32.49 |
| Hulling (%) | 1.340 | 1.17 | 1.59 | 1.53 | 1.87 | 1.48 | 71.18 | 58.58 | 2.33 | 1.84 |
| Milling (%) | 2.19 | 1.41 | 2.40 | 2.01 | 2.91 | 17.54 | 82.68 | 48.84 | 4.09 | 2.03 |
| Head rice (%) | 16.01 | 15.39 | 16.25 | 16.24 | 18.87 | 17.54 | 97.00 | 89.85 | 32.47 | 30.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awad-Allah, M.M.A.; Elekhtyar, N.M.; El-Abd, M.A.-E.-M.; Abdelkader, M.F.M.; Mahmoud, M.H.; Mohamed, A.H.; El-Diasty, M.Z.; Said, M.M.; Shamseldin, S.A.M.; Abdein, M.A. Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.). Genes 2022, 13, 458. https://doi.org/10.3390/genes13030458
Awad-Allah MMA, Elekhtyar NM, El-Abd MA-E-M, Abdelkader MFM, Mahmoud MH, Mohamed AH, El-Diasty MZ, Said MM, Shamseldin SAM, Abdein MA. Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.). Genes. 2022; 13(3):458. https://doi.org/10.3390/genes13030458
Chicago/Turabian StyleAwad-Allah, Mamdouh M. A., Nehal M. Elekhtyar, Mohamed Abd-El-Moaty El-Abd, Mohamed F. M. Abdelkader, Mohamed H. Mahmoud, Azza H. Mohamed, Mohamed Z. El-Diasty, Manal M. Said, Sahar A. M. Shamseldin, and Mohamed A. Abdein. 2022. "Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.)" Genes 13, no. 3: 458. https://doi.org/10.3390/genes13030458
APA StyleAwad-Allah, M. M. A., Elekhtyar, N. M., El-Abd, M. A. -E. -M., Abdelkader, M. F. M., Mahmoud, M. H., Mohamed, A. H., El-Diasty, M. Z., Said, M. M., Shamseldin, S. A. M., & Abdein, M. A. (2022). Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.). Genes, 13(3), 458. https://doi.org/10.3390/genes13030458