
Demographic Reconstruction of Antarctic Fur Seals Supports the Krill Surplus Hypothesis

 ,
,  , , , and
, , , and
Abstract
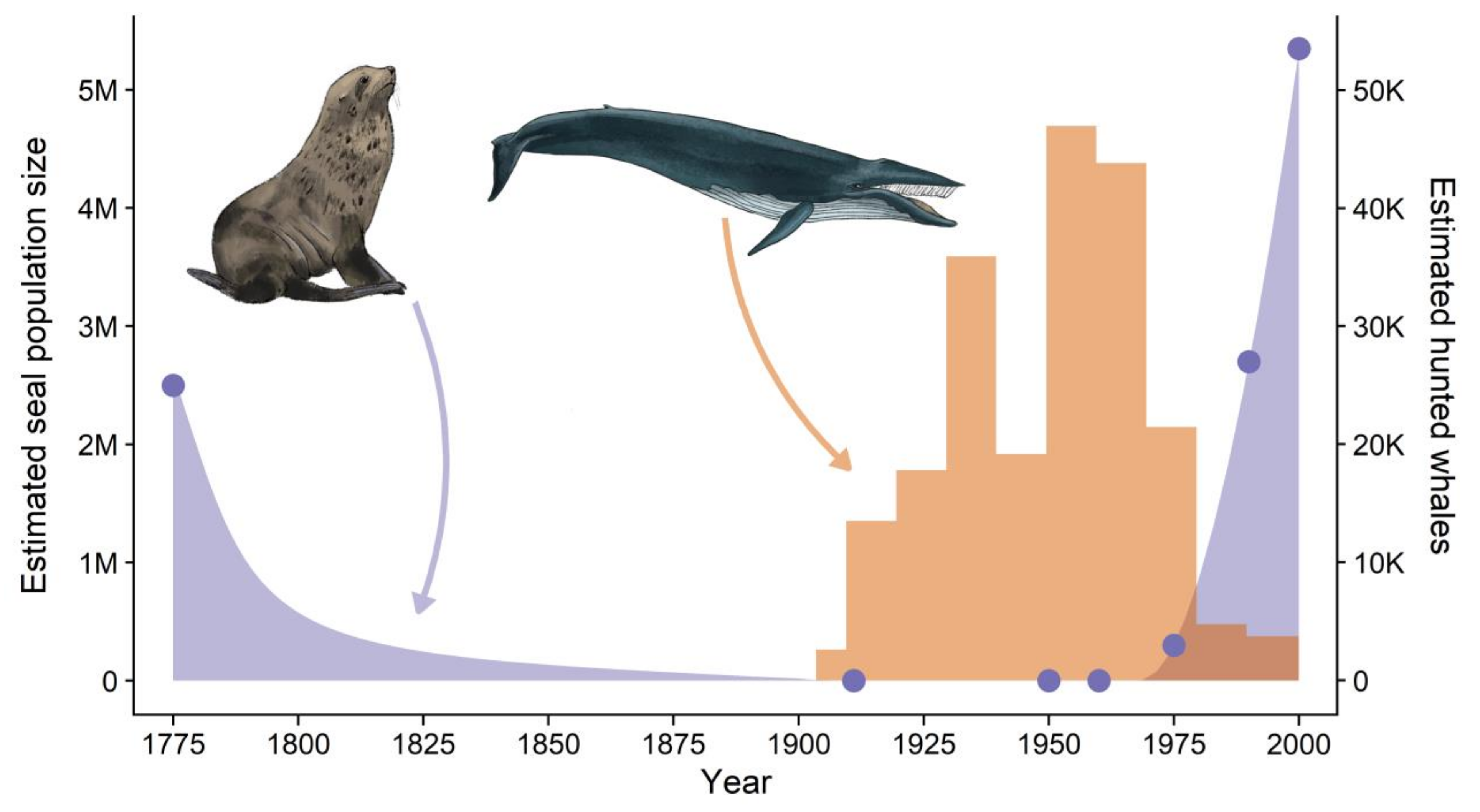
:1. Introduction
2. Materials and Methods
2.1. RAD Sequencing Data
2.2. Bioinformatic Data Processing
2.3. Demographic Modelling
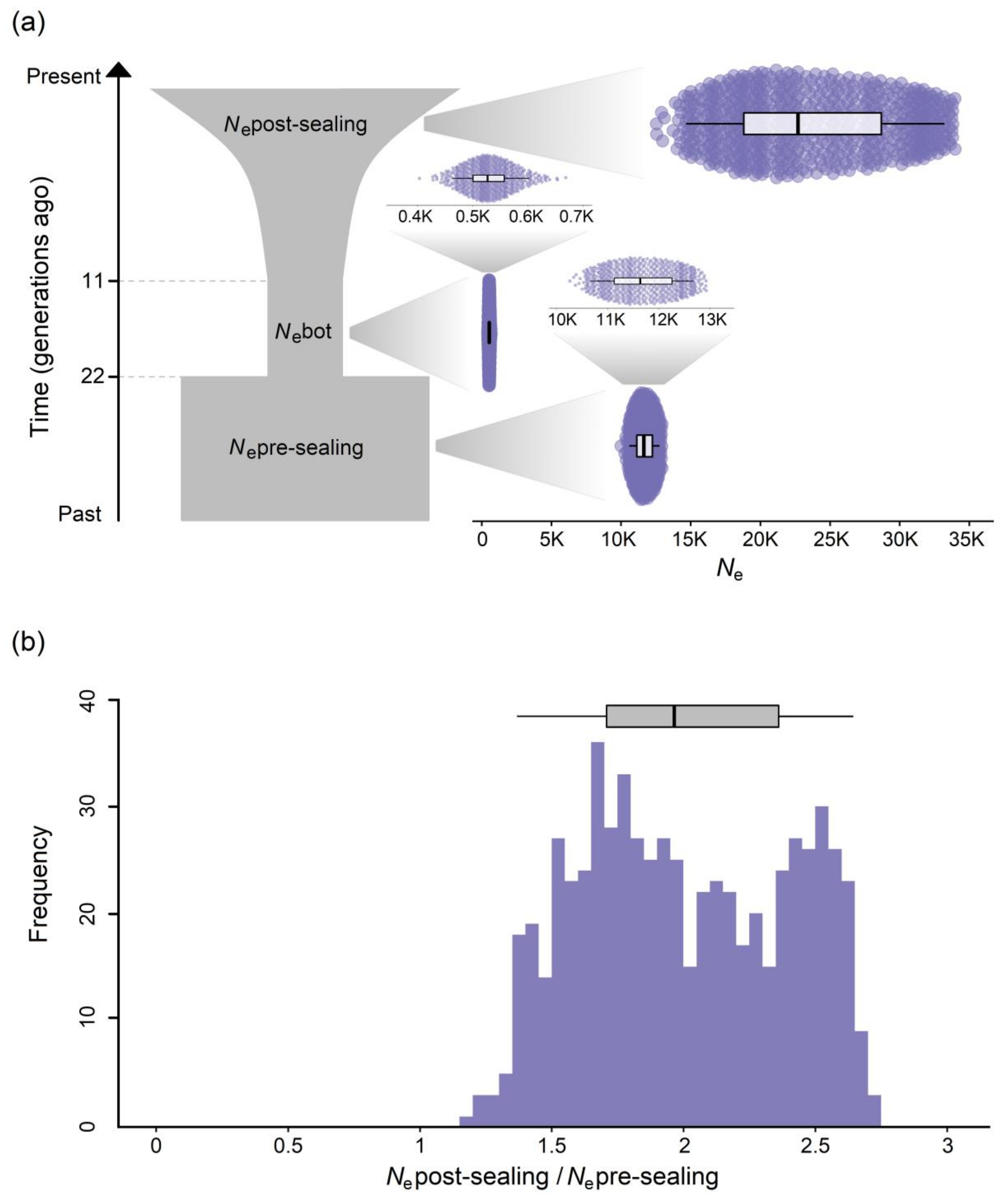
3. Results
4. Discussion
4.1. Bottleneck Inference
4.2. Pre- and Post-Sealing Effective Population Sizes
4.3. The Role of Bottom-Up Effects
4.4. Caveats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enquist, B.J.; Abraham, A.J.; Harfoot, M.B.J.; Malhi, Y.; Doughty, C.E. The megabiota are disproportionately important for biosphere functioning. Nat. Commun. 2020, 11, 699. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Roman, J.; Faurby, S.; Wolf, A.; Haque, A.; Bakker, E.S.; Malhi, Y.; Dunning, J.B.; Svenning, J.-C. Global nutrient transport in a world of giants. Proc. Natl. Acad. Sci. USA 2016, 113, 868–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofman, R.J. Sealing, whaling and krill fishing in the Southern Ocean: Past and possible future effects on catch regulations. Polar Rec. 2017, 53, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Surma, S.; Pakhomov, E.A.; Pitcher, T.J. Effects of Whaling on the Structure of the Southern Ocean Food Web: Insights on the “Krill Surplus” from Ecosystem Modelling. PLoS ONE 2014, 9, e114978. [Google Scholar] [CrossRef]
- Mori, M.; Butterworth, D.S. A first step towards modelling the krill-predator dynamics of the Antarctic ecosystem. CCAMLR Sci. 2006, 13, 217–277. [Google Scholar]
- Laws, R.M. Seals and whales of the Southern Ocean. Philosphical Trans. R. Soc. Lond. B 1977, 279, 81–96. [Google Scholar]
- Trathan, P.N.; Ratcliffe, N.; Masden, E.A. Ecological drivers of change at South Georgia: The krill surplus, or climate variability. Ecography 2012, 35, 983–993. [Google Scholar] [CrossRef]
- Trivelpiece, W.Z.; Hinke, J.T.; Miller, A.K.; Reiss, C.S.; Trivelpiece, S.G.; Watters, G.M. Variability in krill biomass links harvesting and climate warming to penguin population changes in Antarctica. Proc. Natl. Acad. Sci. USA 2011, 108, 7625–7628. [Google Scholar] [CrossRef] [Green Version]
- Rocha, C.R., Jr.; Clapham, P.; Ivashchenko, Y. Emptying the oceans: A summary of industrial whaling catches in the 20th century. Mar. Fish. Rev. 2015, 76, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Smetacek, V. Are declining Antarctic krill stocks a result of global warming or decimation of the whales? In Invited Lecture, Scientific Debate on Impacts of Global Warming on Polar Ecosystems; Durarte, C.M., Ed.; BBVA Foundation: Madrid, Spain, 2008. [Google Scholar]
- Savoca, M.S.; Czapanskiy, M.F.; Kahane-Rapport, S.R.; Gough, W.T.; Fahlbusch, J.A.; Bierlich, K.C.; Segre, P.S.; Di Clemente, J.; Penry, G.S.; Wiley, D.N.; et al. Baleen whale prey consumption based on high-resolution foraging measurements. Nature 2021, 599, 85–90. [Google Scholar] [CrossRef]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P.; Loeb, V.; Ross, R.; Quetin, L.; Schmidt, K.; Fretwell, P.; Murphy, E.; et al. Oceanic circumpolar habitats of Antarctic krill. Mar. Ecol. Prog. Ser. 2008, 362, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Forcada, J.; Hoffman, J.I. Climate change selects for heterozygosity in a declining fur seal population. Nature 2014, 511, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Croxall, J.P.; Trathan, P.N.; Murphy, E.J. Environmmental change and Antarctic seabird populations. Science 2002, 297, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Nicol, S.; Croxall, J.P.; Trathan, P.; Gales, N.; Murphy, E. Paradigm misplaced? Antarctic marine ecosystems are affected by climate change as well as biological processes and harvesting. Antarct. Sci. 2007, 19, 291–295. [Google Scholar] [CrossRef]
- Ainley, D.; Ballard, G.; Ackley, S.; Blight, L.K.; Eastman, J.T.; Emslie, S.D.; Lescroel, A.; Olmastroni, S.; Townsend, S.E.; Tynan, C.T.; et al. Paradigm lost, or is top-down forcing no longer significant in the Antarctic marine ecosystem? Antarct. Sci. 2007, 19, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, D.A.; Johnston, N.M.; Caulkett, A.P.; Jones, V.J. Palaeolimnology of Antarctic fur seal Arctocephalus gazella populations and implications for Antarctic management. Biol. Conserv. 1998, 83, 145–154. [Google Scholar] [CrossRef]
- Emslie, S.D.; Polito, M.J.; Patterson, W.P. Stable isotope analysis of ancient and modern gentoo penguin egg membrane and the krill surplus hypothesis in Antarctica. Antarct. Sci. 2013, 25, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Emslie, S.D.; Patterson, W.P. Abrupt recent shift in 13C and 15N values in Adelie penguin eggshell in Antarctica. Proc. Natl. Acad. Sci. USA 2007, 104, 11666–11669. [Google Scholar] [CrossRef] [Green Version]
- Croxall, J.P. Southern ocean environmental changes: Effects on seabird, seal and whale populations. Philosphical Trans. R. Soc. Lond. B 1992, 338, 319–328. [Google Scholar]
- Fraser, W.R.; Trivelpiece, W.Z.; Ainley, D.G.; Trivelpiece, S.G. Increases in Antarctic penguin populations: Reduced competition with whales or a loss of sea ice due to environmental warming? Polar Biol. 1992, 11, 521–531. [Google Scholar] [CrossRef]
- Ruegg, K.C.; Anderson, E.C.; Scott Baker, C.; Vant, M.; Jackson, J.A.; Palumbi, S.R. Are Antarctic minke whales unusually abundant because of 20th century whaling? Mol. Ecol. 2010, 19, 281–291. [Google Scholar] [CrossRef] [PubMed]
- The Institute Of Cetacean Research. ICR Comments on the Paper ‘Are Antarctic Minke Whales Uniusually Abundant Because of 20th Century Whaling? Available online: https://www.icrwhale.org/pdf/100120Release.pdf (accessed on 15 February 2022).
- Frankham, R. Effective population size/adult population size ratios in wildlife: A review. Genet. Res. 1995, 66, 95–107. [Google Scholar] [CrossRef]
- Peart, C.R.; Tusso, S.; Pophaly, S.D.; Botero-Castro, F.; Wu, C.C.; Aurioles-Gamboa, D.; Baird, A.B.; Bickham, J.W.; Forcada, J.; Galimberti, F.; et al. Determinants of genetic variation across eco-evolutionary scales in pinnipeds. Nat. Ecol. Evol. 2020, 4, 1095–1104. [Google Scholar] [CrossRef]
- Nielsen, R. Estimation of population parameters and recombination rates from single nucleotide polymorphisms. Genetics 2000, 154, 931–942. [Google Scholar] [CrossRef]
- Nei, M.; Maruyama, T.; Chakraborty, R. The Bottleneck Effect and Genetic Variability in Populations. Evolution 1975, 29, 1. [Google Scholar] [CrossRef]
- Tajima, F. The effect of change in population size on DNA polymorphism. Genetics 1989, 123, 579–601. [Google Scholar] [CrossRef]
- Slatkin, M.; Hudson, R.R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 1991, 129, 555–562. [Google Scholar] [CrossRef]
- Marth, G.T.; Czabarka, E.; Murvai, J.; Sherry, S.T. The Allele Frequency Spectrum in Genome-Wide Human Variation Data Reveals Signals of Differential Demographic History in Three Large World Populations. Genetics 2004, 166, 351–372. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.M.; Hudson, R.R. Maximum-Likelihood Estimation of Demographic Parameters Using the Frequency Spectrum of Unlinked Single-Nucleotide Polymorphisms. Genetics 2004, 168, 1699–1712. [Google Scholar] [CrossRef] [Green Version]
- Gutenkunst, R.N.; Hernandez, R.D.; Williamson, S.H.; Bustamante, C.D. Inferring the joint demographic history of multiple populations from multidimensional SNP frequency data. PLoS Genet. 2009, 5, e1000695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H. The joint allele frequency spectrum of multiple populations: A coalescent theory approach. Theor. Popul. Biol. 2012, 81, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Dupanloup, I.; Huerta-Sanchez, E.; Sousa, V.C.; Foll, M. Robust demographic inference from genomic and SNP data. PLoS Genet. 2013, 9, e1003905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaspinas, A.-S.; Westaway, M.C.; Muller, C.; Sousa, V.C.; Lao, O.; Alves, I.; Bergström, A.; Athanasiadis, G.; Cheng, J.Y.; Crawford, J.E.; et al. A genomic history of Aboriginal Australia. Nature 2016, 538, 207–214. [Google Scholar] [CrossRef]
- Batini, C.; Hallast, P.; Zadik, D.; Delser, P.M.; Benazzo, A.; Ghirotto, S.; Arroyo-Pardo, E.; Cavalleri, G.L.; De Knijff, P.; Dupuy, B.M.; et al. Large-scale recent expansion of European patrilineages shown by population resequencing. Nat. Commun. 2015, 6, 7152. [Google Scholar] [CrossRef] [Green Version]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresco, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Roesti, M.; Kueng, B.; Moser, D.; Berner, D. The genomics of ecological vicariance in threespine stickleback fish. Nat. Commun. 2015, 6, 8767. [Google Scholar] [CrossRef] [Green Version]
- Pujolar, J.M.; Dalén, L.; Hansen, M.M.; Madsen, J. Demographic inference from whole-genome and RAD sequencing data suggests alternating human impacts on goose populations since the last ice age. Mol. Ecol. 2017, 26, 6270–6283. [Google Scholar] [CrossRef]
- Nunziata, S.O.; Lance, S.L.; Scott, D.E.; Lemmon, E.M.; Weisrock, D.W. Genomic data detect corresponding signatures of population size change on an ecological time scale in two salamander species. Mol. Ecol. 2017, 26, 1060–1074. [Google Scholar] [CrossRef]
- Pedersen, C.-E.T.; Albrechtsen, A.; Etter, P.D.; Johnson, E.A.; Orlando, L.; Chikhi, L.; Siegismund, H.R.; Heller, R. A southern African origin and cryptic structure in the highly mobile plains zebra. Nat. Ecol. Evol. 2018, 2, 491–498. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Boyd, I.L.; Amos, W. Male reproductive strategy and the importance of maternal status in the Antarctic fur seal Arctocephalus gazella. Evolution 2003, 57, 1917–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.I.; Trathan, P.N.; Amos, W. Genetic tagging reveals extreme site fidelity in territorial male Antarctic fur seals Arctocephalus gazella. Mol. Ecol. 2006, 15, 3841–3847. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Forcada, J. Extreme natal philopatry in female Antarctic fur seals (Arctocephalus gazella). Mamm. Biol. 2012, 77, 71–73. [Google Scholar] [CrossRef]
- Bonin, C.A.; Goebel, M.E.; Hoffman, J.I.; Burton, R.S. High male reproductive skew in a low density Antarctic fur seal (Arctocephalus gazella) breeding colony. Behav. Ecol. 2014, 68, 597–604. [Google Scholar] [CrossRef]
- Boyd, I.L. Pup production and distribution of breeding Antarctic fur seals (Arctocephalus gazella) at South Georgia. Antarct. Sci. 1993, 5, 17–24. [Google Scholar] [CrossRef]
- Weddell, J. A Voyage Towards the South Pole Performed in the Years 1822–1824; Longman, Hurst, Rees, Orme, Brown and Green: London, UK, 1827. [Google Scholar]
- Larsen, C.A. Report of the Interdepartmental Committee on Research and Development in the Dependencies of the Falklands Islands; Sage Publications, Inc.: Thousand Oaks, CA, USA; London, UK, 1920. [Google Scholar]
- Payne, M.R. Growth of a fur seal population. Philos. Trans. R. Soc. London. B Biol. Sci. 1997, 279, 67–79. [Google Scholar] [CrossRef]
- Scientific Committee on Antarctic Research, Expert Group on Seals. Status of Stocks; Scientific Committee on Antarctic Research: Cambridge, UK, 2008. [Google Scholar]
- Payne, M.R. Population size and age determination in the Antarctic fur seal Arctocephalus gazella. Mammal Rev. 1978, 8, 67–73. [Google Scholar] [CrossRef]
- Bonner, W.N. Impact of fur seals on the terrestrial environment at South Georgia. In Antarctic Nutrient Cycles and Food Webs; Siegfried, W.R., Condy, P.R., Laws, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 641–646. [Google Scholar]
- Croxall, J.P.; McCann, T.S.; Prince, P.A.; Rothery, P. Reproductive performance of seabirds and seals at South Georgia and Signey Island, South Orkney Islands, 1976–1987: Implications for Southern Ocean monitoring studies. In Antarctic Ocean and Resources Variability; Sahrhage, D., Ed.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 262–285. [Google Scholar]
- Foley, C.M.; Lynch, H.J. A method to estimate pre-exploitation population size. Conserv. Biol. 2020, 34, 256–265. [Google Scholar] [CrossRef]
- Laws, R.M. Population increase of fur seals at South Georgia. Polar Rec. 1973, 16, 856–858. [Google Scholar] [CrossRef]
- Bonner, W.N. The status of the Antarctic fur seal Arctocephalus gazella. In Proceedings of the Advisory Committee on Marine Resources Research, Scientific Consultation on Marine Mammals, Bergen, Norway, 31 August 1976. [Google Scholar]
- Leaper, R.; Bannister, J.L.; Branch, T.A.; Clapham, P.; Donovan, G.; Matsuoka, K.; Reilly, S.; Zerbini, A. A Review of Abundance, Trends and Foraging Parameters of Baleen Whales in the Southern Hemisphere; IWC Scientific Committee: Cambridge, UK, 2008. [Google Scholar]
- Hoffman, J.I.; Grant, S.M.; Forcada, J.; Phillips, C.D. Bayesian inference of a historical bottleneck in a heavily exploited marine mammal. Mol. Ecol. 2011, 20, 3989–4008. [Google Scholar] [CrossRef]
- Humble, E.; Dasmahapatra, K.K.; Martinez-Barrio, A.; Gregorio, I.; Forcada, J.; Polikeit, A.C.; Goldsworthy, S.D.; Goebel, M.E.; Kalinowski, J.; Wolf, J.B.W.; et al. RAD Sequencing and a hybrid Antarctic fur seal genome assembly reveal rapidly decaying linkage disequilibrium, global population structure and evidence for inbreeding. G3 Genes Genomes Genet. 2018, 8, 2709–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, R.; Kaiser, S.; Stainfield, C.; Toscani, C.; Fox-Clarke, C.; Paijmans, A.J.; Costa Castro, C.; Vendrami, D.; Forcada, J.; Hoffman, J.I. Low heritability and high phenotypic plasticity of cortisol in response to environmental heterogeneity in fur seals. bioRxiv, 2021; Preprint. [Google Scholar] [CrossRef]
- Etter, P.D.; Preston, J.L.; Bassham, S.; Cresko, W.A.; Johnson, E.A. Local de novo assembly of RAD paired-end contigs using short sequencing reads. PLoS ONE 2011, 6, e18561. [Google Scholar] [CrossRef] [PubMed]
- Catchen, J.M.; Amores, A.; Hohenlohe, P.; Cresko, W.; Postlethwait, J.H. Stacks: Building and genotyping Loci de novo from short-read sequences. G3 Genes Genomes Genet. 2011, 1, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Peart, C.R.; Williams, C.; Pophaly, S.D.; Johnson, J.; Neely, B.A.; Gulland, F.; Adams, D.; Ng, B.L.; Cheng, W.; Goebel, M.E.; et al. Hi-C scaffolded short- and long-read genome assemblies of the California sea lion are broadly consistent for syntenic inference across 45 million years of evolution. Mol. Ecol. Resour. 2021, 21, 2455–2470. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Korneliussen, T.S.; Albrechtsen, A.; Nielsen, R. ANGSD: Analysis of Next Generation Sequencing Data. BMC Bioinform. 2014, 15, 356. [Google Scholar] [CrossRef] [Green Version]
- Forcada, J.; Trathan, P.N.; Murphy, E.J. Life history buffering in Antarctic mammals and birds against changing patterns of climate and environmental variation. Glob. Change Biol. 2008, 14, 2473–2488. [Google Scholar] [CrossRef]
- Dornburg, A.; Brandley, M.C.; McGowen, M.R.; Near, T.J. Relaxed Clocks and Inferences of Heterogeneous Patterns of Nucleotide Substitution and Divergence Time Estimates across Whales and Dolphins (Mammalia: Cetacea). Mol. Biol. Evol. 2012, 29, 721–736. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Inference of human population history from individual whole-genome sequences. Nature 2011, 475, 493–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffel, M.A.; Humble, E.; Paijmans, A.J.; Acevedo-Whitehouse, K.; Chilvers, B.L.; Dickerson, B.; Galimberti, F.; Gemmell, N.J.; Goldsworthy, S.D.; Nichols, H.J.; et al. Demographic histories and genetic diversity across pinnipeds are shaped by human exploitation, ecology and life-history. Nat. Commun. 2018, 9, 4836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paijmans, A.J.; Stoffel, M.A.; Bester, M.N.; Cleary, A.C.; De Bruyn, P.J.N.; Forcada, J.; Goebel, M.E.; Goldsworthy, S.D.; Guinet, C.; Lydersen, C.; et al. The genetic legacy of extreme exploitation in a polar vertebrate. Sci. Rep. 2020, 10, 5089. [Google Scholar] [CrossRef] [PubMed]
- Cleary, A.C.; Bester, M.; Forcada, J.; Goebel, M.; Goldsworthy, S.D.; Guinet, C.; Hoffman, J.I.; Kovacs, K.M.; Lydersen, C.; Lowther, A.D. Prey differences drive local genetic adaptation in Antarctic fur seals. Mar. Ecol. Prog. Ser. 2019, 628, 195–209. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Bauer, E.; Paijmans, A.J.; Humble, E.; Beckmann, L.M.; Kubetschek, C.; Christaller, F.; Krocker, N.; Fuchs, B.; Moreras, A.; et al. A global cline in a colour polymorphism suggests a limited contribution of gene flow towards the recovery of a heavily exploited marine mammal. R. Soc. Open Sci. 2018, 5, 181227. [Google Scholar] [CrossRef] [Green Version]
- Wynen, L.P.; Goldsworthy, S.D.; Guinet, C.; Bester, M.N.; Boyd, I.L.; Gjertz, I.; Hofmeyr, G.J.; White, R.W.; Slade, R. Postsealing genetic variation and population structure of two species of fur seal (Arctocephalus gazella and A. tropicalis). Mol. Ecol. 2000, 9, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Palstra, F.P.; Ruzzante, D.E. Genetic estimates of contemporary effective population size: What can they tell us about the importance of genetic stochasticity for wild population persistence? Mol. Ecol. 2008, 17, 3428–3447. [Google Scholar] [CrossRef]
- Nagel, R.; Stainfield, C.; Fox-Clarke, C.; Toscani, C.; Forcada, J.; Hoffman, J.I. Evidence for an Allee effect in a declining fur seal population. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202882. [Google Scholar] [CrossRef]
- Han, E.; Sinsheimer, J.S.; Novembre, J. Characterizing Bias in Population Genetic Inferences from Low-Coverage Sequencing Data. Mol. Biol. Evol. 2014, 31, 723–735. [Google Scholar] [CrossRef] [Green Version]
- Fountain, E.D.; Pauli, J.N.; Reid, B.N.; Palsbøll, P.J.; Peery, M.Z. Finding the right coverage: The impact of coverage and sequence quality on single nucleotide polymorphism genotyping error rates. Mol. Ecol. Resour. 2016, 16, 966–978. [Google Scholar] [CrossRef]
- Shafer, A.B.A.; Peart, C.R.; Tusso, S.; Maayan, I.; Brelsford, A.; Wheat, C.W.; Wolf, J.B.W. Bioinformatic processing of RAD-seq data dramatically impacts downstream population genetic inference. Methods Ecol. Evol. 2017, 8, 907–917. [Google Scholar] [CrossRef]
- Le, S.Q.; Durbin, R. SNP detection and genotyping from low-coverage sequencing data on multiple diploid samples. Genome Res. 2011, 21, 952–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, S.; Fefferman, C.; Patterson, N. Can one learn history from the allelic spectrum? Theor. Popul. Biol. 2008, 73, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Mill, H.R. The Seige of the South Pole: The Story of Antarctic Exploration; Alston Rivers: London, UK, 1905. [Google Scholar]
- Fanning, E. Voyages and Discoveries in the South Seas 1792–1832; Marine Research Society: Salem, MA, USA, 1924. [Google Scholar]



{kind=link}
{kind=link}
| Model | Max(log10(likelihood)) a | Number of Parameters | AIC | Nepre-Sealing | Nebot | Nepost-Sealing |
|---|---|---|---|---|---|---|
| Bottleneck model | −882,361.6 | 3 | 4,063,431 | 12,506 | 534 | 29,319 |
| Null model | −882,378.8 | 1 | 4,063,506 | – | – | 14,789 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffman, J.I.; Chen, R.S.; Vendrami, D.L.J.; Paijmans, A.J.; Dasmahapatra, K.K.; Forcada, J. Demographic Reconstruction of Antarctic Fur Seals Supports the Krill Surplus Hypothesis. Genes 2022, 13, 541. https://doi.org/10.3390/genes13030541
Hoffman JI, Chen RS, Vendrami DLJ, Paijmans AJ, Dasmahapatra KK, Forcada J. Demographic Reconstruction of Antarctic Fur Seals Supports the Krill Surplus Hypothesis. Genes. 2022; 13(3):541. https://doi.org/10.3390/genes13030541
Chicago/Turabian StyleHoffman, Joseph I., Rebecca S. Chen, David L. J. Vendrami, Anna J. Paijmans, Kanchon K. Dasmahapatra, and Jaume Forcada. 2022. "Demographic Reconstruction of Antarctic Fur Seals Supports the Krill Surplus Hypothesis" Genes 13, no. 3: 541. https://doi.org/10.3390/genes13030541
APA StyleHoffman, J. I., Chen, R. S., Vendrami, D. L. J., Paijmans, A. J., Dasmahapatra, K. K., & Forcada, J. (2022). Demographic Reconstruction of Antarctic Fur Seals Supports the Krill Surplus Hypothesis. Genes, 13(3), 541. https://doi.org/10.3390/genes13030541