
The Complete Chloroplast Genomes of Primula obconica Provide Insight That Neither Species nor Natural Section Represent Monophyletic Taxa in Primula (Primulaceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, DNA Extraction, and Sequencing
2.2. Genome Assembly and Annotation
2.3. Reference Mapping, Variants Calling, and SSR Analysis
2.4. Comparative Analysis of the Chloroplast Genomes
2.5. Population Structure Analysis of P. obconica subsp. obconica
2.6. Phylogenetic Analysis
3. Results
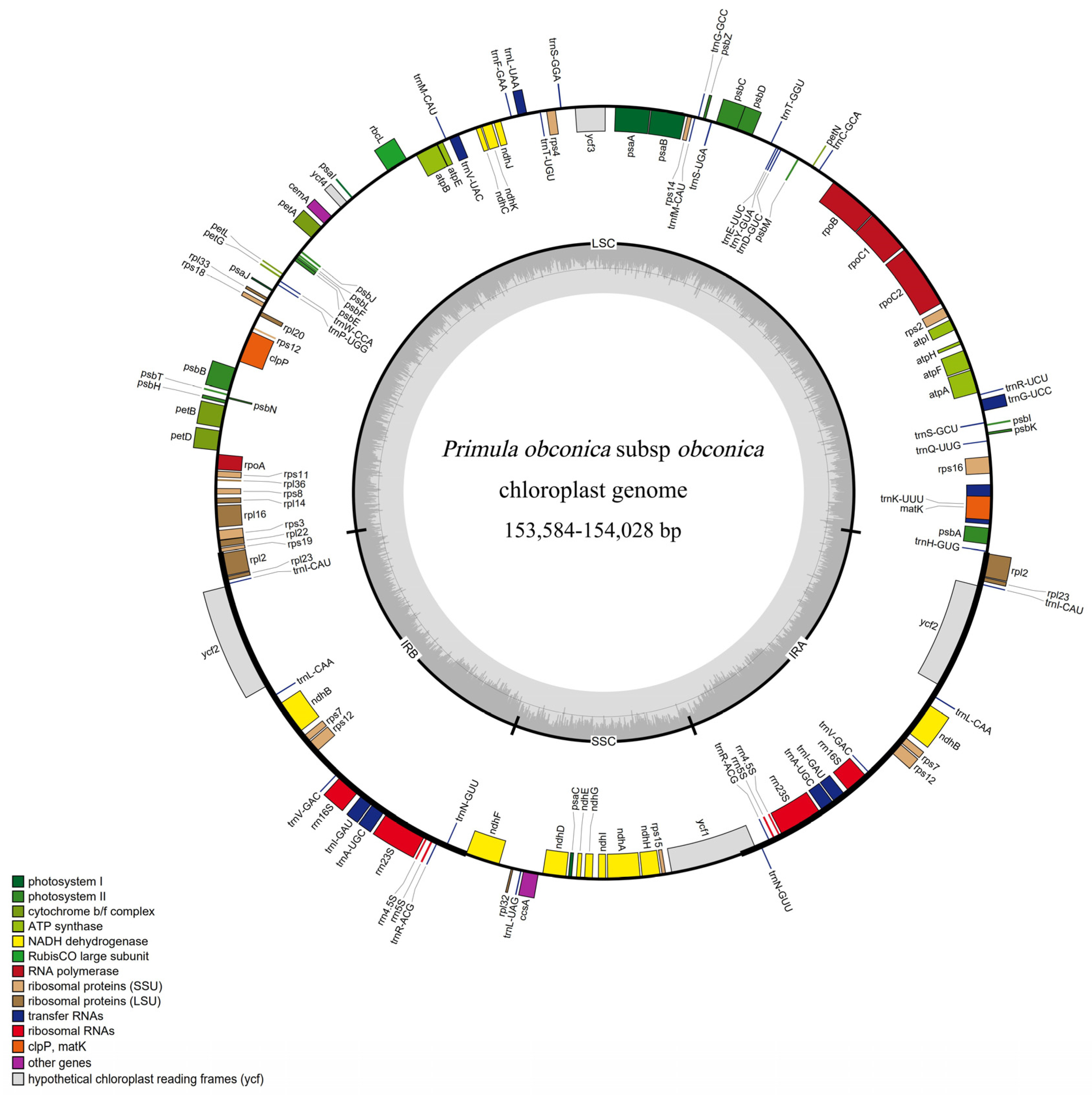
3.1. General Features of the P. obconica subsp. obconica Chloroplast Genome
3.2. Chloroplast Genomic SNP Variations
3.3. Simple Sequence Repeat Analysis
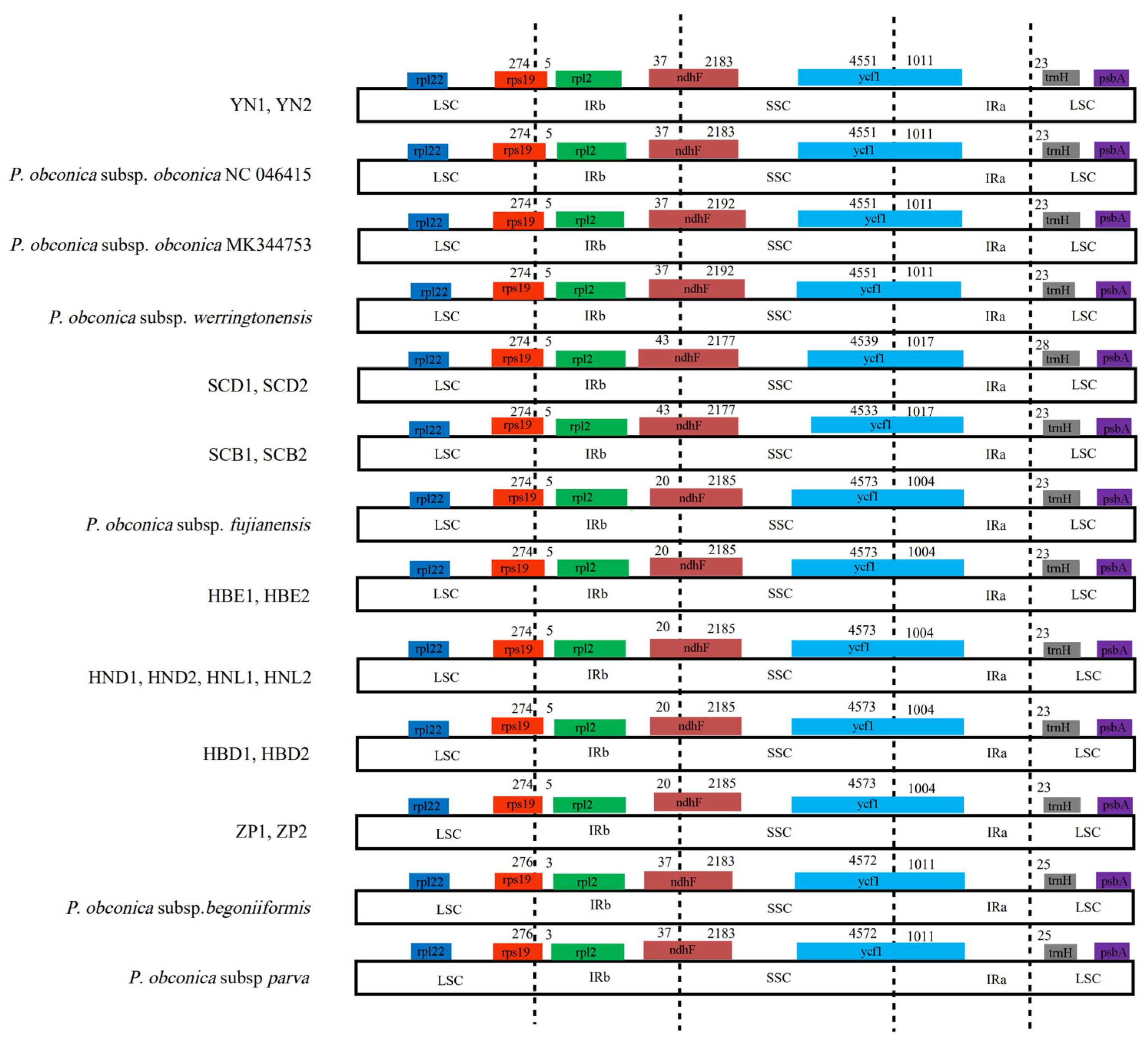
3.4. IR Expansion and Contraction
3.5. Comparative Analysis of the Chloroplast Genomes
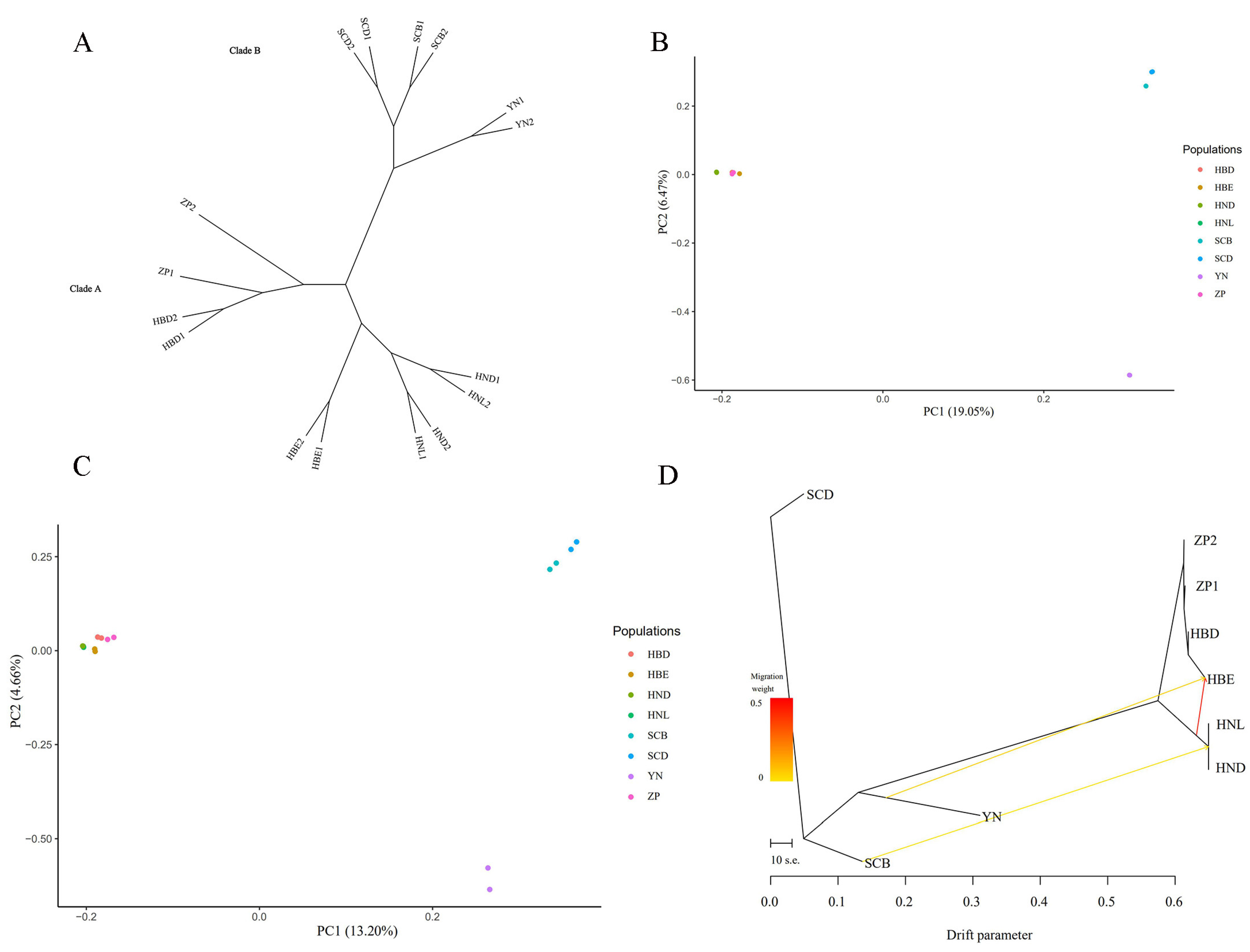
3.6. Population Structure Analysis
3.7. Phylogenetic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shang, H.; Liu, B.; Kang, M.; Yan, Y. One or more species? GBS sequencing and morphological traits evidence reveal species diversification of Sphaeropteris brunoniana in China. Biodivers. Sci. 2019, 27, 1196–1204. [Google Scholar] [CrossRef]
- Murphy, D.J.; Brown, G.K.; Miller, J.T.; Ladiges, P.Y. Molecular phylogeny of Acacia Mill. (Mimosoideae: Leguminosae): Evidence for major clades and informal classification. Taxon 2010, 59, 7–19. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Liu, Y.; Yuan, Q.; Sun, J.; Guo, L. Chloroplast genome variation and phylogenetic relationships of Atractylodes species. BMC Genom. 2021, 22, 103. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Yang, J.; Landis, J.B.; Wang, S.; Yang, Z.; Zhang, Y. Deciphering the Taxonomic Delimitation of Ottelia acuminata (Hydrocharitaceae) Using Complete Plastomes as Super-Barcodes. Front. Plant Sci. 2021, 12, 1356. [Google Scholar] [CrossRef] [PubMed]
- Boutte, J.; Maillet, L.; Chaussepied, T.; Letort, S.; Aury, J.M.; Belser, C.; Boideau, F.; Brunet, A.; Coriton, O.; Deniot, G.; et al. Genome Size Variation and Comparative Genomics Reveal Intraspecific Diversity in Brassica rapa. Front. Plant Sci. 2020, 11, 1579. [Google Scholar] [CrossRef]
- Park, J.; Xi, H.; Kim, Y. The Complete Chloroplast Genome of Arabidopsis thaliana Isolated in Korea (Brassicaceae): An Investigation of Intraspecific Variations of the Chloroplast Genome of Korean A. thaliana. Int. J. Genom. 2020, 2020, 3236461. [Google Scholar] [CrossRef]
- Muraguri, S.; Xu, W.; Chapman, M.; Muchugi, A.; Oluwaniyi, A.; Oyebanji, O.; Liu, A. Intraspecific variation within Castor bean (Ricinus communis L.) based on chloroplast genomes. Ind. Crops Prod. 2020, 155, 112779. [Google Scholar] [CrossRef]
- Smith, D.R. Mutation rates in plastid genomes: They are lower than you might think. Genome Biol. Evol. 2015, 7, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Kelso, S. Flora of China; Science Press: Beijing, China, 1996; pp. 99–108. [Google Scholar]
- Mast, A.R.; Kelso, S.; Richards, A.J.; Lang, D.J.; Feller, D.M.S.; Conti, E.; Journal, I.; Mast, A.R.; Kelso, S.; Richards, A.J.; et al. Phylogenetic relationships in Primula L. and related genera (Primulaceae) based on noncoding chloroplast DNA. Int. J. Plant Sci 2001, 162, 1381–1400. [Google Scholar] [CrossRef] [Green Version]
- De Vos, J.M.; Hughes, C.E.; Schneeweiss, G.M.; Moore, B.R.; Conti, E. Heterostyly accelerates diversification via reduced extinction in primroses. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140075. [Google Scholar] [CrossRef] [Green Version]
- Nuraliev, M.S.; Kuznetsov, A.N.; Kuznetsova, S.P.; Hu, C.M. Primula gracilituba (Primulaceae), a new species from northern Vietnam. Nord. J. Bot. 2020, 38, 1–4. [Google Scholar] [CrossRef]
- Ma, X.D.; Wang, W.G.; Shi, J.P.; Shen, J.Y. Primula longistyla (Primulaceae), a new species from Yunnan, China. Nord. J. Bot. 2021, 39, 1–5. [Google Scholar] [CrossRef]
- Ju, W.B.; Huang, Q.; Sun, Z.Y.; Huang, W.J.; Li, H.C.; Gao, X.F. Primula luteoflora (Primulaceae), a new species from Sichuan, China. Phytotaxa 2018, 367, 297–300. [Google Scholar] [CrossRef]
- Zhang, L.B.; Comes, H.P.; Kadereit, J.W. The temporal course of quaternary diversification in the European high mountain endemic Primula sect. Auricula (Primulaceae). Int. J. Plant Sci. 2004, 165, 191–207. [Google Scholar] [CrossRef]
- Yan, H.F.; Liu, Y.J.; Xie, X.F.; Zhang, C.Y.; Hu, C.M.; Hao, G.; Ge, X.J. DNA barcoding evaluation and its taxonomic implications in the species-rich genus Primula L. in China. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Li, J.; Li, M.; Zhu, W.; Zhang, X. The reproductive characteristics of Primula obconica. J. Hunan Agric. Univ. 2020, 46, 558–564. [Google Scholar] [CrossRef]
- Yuan, S.; Barrett, S.C.H.; Duan, T.; Qian, X.; Shi, M.; Zhang, D. Ecological correlates and genetic consequences of evolutionary transitions from distyly to homostyly. Ann. Bot. 2017, 120, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; You, Y.; Zhang, X. Advances in the research of heterostyly. Acta Prataculturae Sin. 2010, 19, 226–239. [Google Scholar]
- Tendal, K.; Ørgaard, M.; Larsen, B.; Pedersen, C. Recurrent hybridisation events between Primula vulgaris, P. veris and P. elatior (Primulaceae, Ericales) challenge the species boundaries: Using molecular markers to re-evaluate morphological identifications. Nord. J. Bot. 2018, 36, e01778. [Google Scholar] [CrossRef]
- Cianchi, R.; Arduino, P.; Mosco, M.C.; Bullini, L. Evidence of hybrid speciation in the North American primroses Primula suffrutescens P. parryi P. rusbyi and P. angustifolia (Primulaceae). Plant Biosyst. 2015, 149, 229–234. [Google Scholar] [CrossRef]
- He, G.-S.; Hu, C.-M. A new subspecies of Primula obconica Hance from Eastern China. Acta Phytotaxon. Sin. 2002, 40, 551–552. [Google Scholar]
- Richards, J. Primula (New Edition); BT Batsford Ltd.: London, UK, 2002. [Google Scholar]
- Smith, W.; Forrest, G. The sections of the genus Primula. In Notes from the Royal Botanic Garden; Royal Society: London, UK, 1928; Volume 16, pp. 1–50. [Google Scholar]
- Yan, H.; Wang, X.L.; Hu, Q.; Hao, G. Phylogeography of Primula obconica Hance (Primulaceae). J. Trop. Subtrop. Bot. 2005, 13, 526–532. [Google Scholar]
- Nan, P.; Peng, S.; Zhang, Y.; Zhong, Y. Composition of volatile oil of Primula obconica in central China. Nat. Prod. Lett. 2002, 16, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Mowad, C.M. Routine testing for Primula obconica: Is it useful in the United States? Am. J. Contact Dermat. 1998, 9, 231–233. [Google Scholar] [CrossRef]
- Yan, H.F.; Ge, X.J.; Hu, C.M.; Hao, G. Isolation and characterization of microsatellite loci for the ornamental plant Primula obconica Hance (Primulaceae). HortScience 2010, 45, 314–315. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–501. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lischer, H.E.L.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Pickrell, J.K.; Pritchard, J.K. Inference of Population Splits and Mixtures from Genome-Wide Allele Frequency Data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Teshome, G.E.; Mekbib, Y.; Hu, G.; Li, Z.Z.; Chen, J. Comparative analyses of 32 complete plastomes of Tef (Eragrostis tef) accessions from Ethiopia: Phylogenetic relationships and mutational hotspots. PeerJ 2020, 2020, 1–20. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.; Lee, G.H.; Park, C.H. The complete chloroplast genome of Selaginella tamariscina (Beauv.) Spring (Selaginellaceae) isolated in Korea. Mitochondrial DNA Part B Resour. 2020, 5, 1654–1656. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.H.; Suh, H.J.; Park, J.; Kim, Y.; Kim, S. The complete chloroplast genome sequence of Goodyera schlechtendaliana in Korea (Orchidaceae). Mitochondrial DNA Part B Resour. 2019, 4, 2692–2693. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.Y.; Liu, J.; Gao, C.W.; Wu, H.B.; Li, G.H.; Gao, L.Z. Higher Genomic Variation in Wild Than Cultivated Rubber Trees, Hevea brasiliensis, Revealed by Comparative Analyses of Chloroplast Genomes. Front. Ecol. Evol. 2020, 8, 237. [Google Scholar] [CrossRef]
- Hilu, K.; Alice, L.; Liang, H. Phylogeny of Poaceae inferred from matK sequences. Ann. Mo. Bot. Gard. 1999, 86, 835–851. [Google Scholar] [CrossRef]
- Lehnebach, C.A.; Cano, A.; Monsalve, C.; McLenachan, P.; Hörandl, E.; Lockhart, P. Phylogenetic relationships of the monotypic Peruvian genus Laccopetalum (Ranunculaceae). Plant Syst. Evol. 2007, 264, 109–116. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Chen, Q.; Luo, Y.; Zhang, Y.; Tang, H.; Wang, X. Phylogenetic Utility of Chinese Rubus (Rosaceae) Based on ndhF Sequence. Acta Hortic. Sin. 2015, 42, 19–30. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Xia, B.; Li, X. The complete chloroplast genome sequences of five pinnate-leaved Primula species and phylogenetic analyses. Sci. Rep. 2020, 10, 20782. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, A.; Ravinet, M.; Sætre, G.P.; Eroukhmanoff, F. Intraspecific genomic variation and local adaptation in a young hybrid species. Mol. Ecol. 2021, 30, 791–809. [Google Scholar] [CrossRef]
- Fu, Y.-B. Characterizing chloroplast genomes and inferring maternal divergence of the Triticum–Aegilops complex. Sci. Rep. 2021, 11, 15363. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Taniguchi, F.; Kimura, K.; Saba, T.; Ogino, A.; Yamaguchi, S.; Tanaka, J. Worldwide core collections of tea (Camellia sinensis) based on SSR markers. Tree Genet. Genomes 2014, 10, 1555–1565. [Google Scholar] [CrossRef]
- Liu, H.; Hu, H.; Zhang, S.; Jin, J.; Liang, X.; Huang, B.; Wang, L. The complete chloroplast genome of the rare species Epimedium tianmenshanensis and comparative analysis with related species. Physiol. Mol. Biol. Plants 2020, 26, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Tang, D.; Wei, K.; Qin, F.; Li, L.; Lin, Y.; Zhu, Y.; Khan, A.; Kashif, M.H.; Miao, J. The complete chloroplast genome sequence of the medicinal plant Sophora tonkinensis. Sci. Rep. 2020, 10, 12473. [Google Scholar] [CrossRef] [PubMed]
- Raman, G.; Park, S.J. The complete chloroplast genome sequence of the speirantha gardenii: Comparative and adaptive evolutionary analysis. Agronomy 2020, 10, 1405. [Google Scholar] [CrossRef]
- YANG, C.H.; LIU, X.; CUI, Y.X.; NIE, L.P.; LIN, Y.L.; WEI, X.P.; WANG, Y.; YAO, H. Molecular structure and phylogenetic analyses of the complete chloroplast genomes of three original species of Pyrrosiae Folium. Chin. J. Nat. Med. 2020, 18, 573–581. [Google Scholar] [CrossRef]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.-Y.; Gao, L.-Z. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. BMC Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.L.H.; Park, H.S.; Lau, T.W.D.; Lin, Z.; Yang, T.J.; Shaw, P.C. Comparative analysis and phylogenetic investigation of Hong Kong Ilex chloroplast genomes. Sci. Rep. 2021, 11, 5153. [Google Scholar] [CrossRef]
- Zheng, G.; Wei, L.; Ma, L.; Wu, Z.; Gu, C.; Chen, K. Comparative analyses of chloroplast genomes from 13 Lagerstroemia (Lythraceae) species: Identification of highly divergent regions and inference of phylogenetic relationships. Plant Mol. Biol. 2020, 102, 659–676. [Google Scholar] [CrossRef]
- Hu, Q. On the geographical distribution of the Primulaceae. J. Trop. Subtrop. Bot. 1994, 2, 1–14. [Google Scholar]
- Hao, C.-Y.; Tan, L.-H.; Fan, R.; Yu, H.; Yang, J.-F.; Wu, H.-S. Floristic Geography of Piper (Piperaceae) in China. Plant Divers. Resour. 2012, 34, 421. [Google Scholar] [CrossRef]
- Cui, H.-X.; Jiang, G.-M.; Zhang, S.-Y. The distribution, origin and evolution of Syringa. Bull. Bot. Res. 2004, 24, 141–145. [Google Scholar]
- Yan, H.F.; Peng, C.I.; Hu, C.M.; Hao, G. Phylogeographic structure of Primula obconica (Primulaceae) inferred from chloroplast microsatellites (cpSSRs) markers. Acta Phytotaxon. Sin. 2007, 45, 488–496. [Google Scholar] [CrossRef]
- Yan, H.F.; Zhang, C.Y.; Wang, F.Y.; Hu, C.M.; Ge, X.J.; Hao, G. Population Expanding with the Phalanx Model and Lineages Split by Environmental Heterogeneity: A Case Study of Primula obconica in Subtropical China. PLoS ONE 2012, 7, e41315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.C.; Boufford, D.E.; Ree, R.H.; Sun, H. The 29° N latitudinal line: An important division in the Hengduan Mountains, a biodiversity hotspot in southwest China. Nord. J. Bot. 2009, 27, 405–412. [Google Scholar] [CrossRef]
- Zhong, L.; Barrett, S.C.H.; Wang, X.J.; Wu, Z.K.; Sun, H.Y.; Li, D.Z.; Wang, H.; Zhou, W. Phylogenomic analysis reveals multiple evolutionary origins of selfing from outcrossing in a lineage of heterostylous plants. New Phytol. 2019, 224, 1290–1303. [Google Scholar] [CrossRef]
- Syring, J.; Farrell, K.; Businský, R.; Cronn, R.; Liston, A. Widespread genealogical nonmonophyly in species of Pinus subgenus Strobus. Syst. Biol. 2007, 56, 163–181. [Google Scholar] [CrossRef] [Green Version]
- McKay, B.D.; Zink, R.M. The causes of mitochondrial DNA gene tree paraphyly in birds. Mol. Phylogenet. Evol. 2010, 54, 647–650. [Google Scholar] [CrossRef]
- Schmidt-Lebuhn, A.N.; de Vos, J.M.; Keller, B.; Conti, E. Phylogenetic analysis of Primula section Primula reveals rampant non-monophyly among morphologically distinct species. Mol. Phylogenet. Evol. 2012, 65, 23–34. [Google Scholar] [CrossRef]
- Mast, A.R.; Feller, D.M.S.; Kelso, S.; Conti, E. Buzz-pollinated Dodecatheon originated from within the heterostylous Primula subgenus Auriculastrum (Primulaceae): A seven-region cpDNA phylogeny and its implications for floral evolution. Am. J. Bot. 2004, 91, 926–942. [Google Scholar] [CrossRef]
- Song, L.; Li, Y.; Zhang, W.; Shao, J. Highly differentiated phylogeographic structure of Primula ranunculoides. Plant Sci. J. 2017, 35, 503–512. [Google Scholar]
- Wang, F.Y.; Ge, X.J.; Gong, X.; Hu, C.M.; Hao, G. Strong genetic differentiation of Primula sikkimensis in the East Himalaya-Hengduan Mountains. Biochem. Genet. 2008, 46, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Tigano, A.; Friesen, V.L. Genomics of local adaptation with gene flow. Mol. Ecol. 2016, 25, 2144–2164. [Google Scholar] [CrossRef]
- Petit, R.J.; Excoffier, L. Gene flow and species delimitation. Trends Ecol. Evol. 2009, 24, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Volis, S.; Ormanbekova, D.; Shulgina, I. Fine-scale spatial genetic structure in predominantly selfing plants with limited seed dispersal: A rule or exception? Plant Divers. 2016, 38, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Ge, X.; Hao, G.; Zhang, C. High genetic diversity in a rare, narrowly endemic primrose species: Primula interjacens by ISSR. Acta Bot. Sin. 2004, 46, 1163–1169. [Google Scholar]
- Van Geert, A.; Van Rossum, F.; Triest, L. Genetic diversity in adult and seedling populations of Primula vulgaris in a fragmented agricultural landscape. Conserv. Genet. 2008, 9, 845–853. [Google Scholar] [CrossRef]
- Kato, J.; Ohashi, H.; Ikeda, M.; Fujii, N.; Ishikawa, R.; Horaguchi, H.; Amano, J.; Hayashi, M.; Mii, M. Unreduced gametes are the major causal factor for the production of polyploid interspecific hybrids in Primula. Plant Biotechnol. 2008, 25, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.P.; Tian, X.L.; Zhang, J.L.; Wu, Z.K.; Sun, W.B. Evidence for natural hybridization between Primula beesiana and P. bulleyana, two heterostylous primroses in NW Yunnan, China. J. Syst. Evol. 2014, 52, 500–507. [Google Scholar] [CrossRef]
- Qian, C.; Luo, J.; Mu, J.; Li, C.; Zhou, Z.; Ou, M.; Lan, S. Studies on Pollination Insect Species and Their Visiting Behavior of Primula lithophila of Endemic Plants in Guizhou. Mol. Plant Breed. 2017, 15, 3838–3844. [Google Scholar] [CrossRef]
- Abou-El-Enain, M.M. Chromosomal variability in the genus Primula (Primulaceae). Bot. J. Linn. Soc. 2006, 150, 211–219. [Google Scholar] [CrossRef]
- Kong, H.; Zhang, Y.; Hong, Y.; Barker, M.S. Multilocus phylogenetic reconstruction informing polyploid relationships of Aconitum subgenus Lycoctonum (Ranunculaceae) in China. Plant Syst. Evol. 2017, 303, 727–744. [Google Scholar] [CrossRef]
- Kim, Y.; Yi, J.S.; Min, J.; Xi, H.; Kim, D.Y.; Son, J.; Park, J.; Jeon, J.I. The complete chloroplast genome of Aconitum coreanum (H. Lév.) Rapaics (Ranunculaceae). Mitochondrial DNA Part B Resour. 2019, 4, 3404–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Kim, W.J.; Yang, S.; Yeo, S.M.; Li, H.; Moon, B.C. The complete chloroplast genome sequence of Aconitum coreanum and Aconitum carmichaelii and comparative analysis with other Aconitum species. PLoS ONE 2017, 12, e0184257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guggisberg, A.; Mansion, G.; Kelso, S.; Conti, E. Evolution of biogeographic patterns, ploidy levels, and breeding systems in a diploid-polyploid species complex of Primula. New Phytol. 2006, 171, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.-Z.; Liu, J.-Q. Karyomorphology of the genus Pomatosace Maxin. (Primulaceae). Acta Phytotaxon. Sin. 1999, 37, 445–450. [Google Scholar]
- Zhu, H.-F.; Yang, J.-B.; Zhang, C.-Q. Systematic position of Primula secundiflora (Primulaceae) inferred from nuclear ribosomal DNA ITS sequence data. Acta Phytotaxon. Sin. 2002, 40, 133–138. [Google Scholar]
- Xu, Y.; Choudhary, R.K.; Hao, G.; Hu, C.M. Primula subansirica G.D. Pal is not a Primula (Primulaceae), but rather belongs to Gesneriaceae. Nord. J. Bot. 2020, 38, 1–4. [Google Scholar] [CrossRef]
- Yi, T.S.; Jin, G.H.; Wen, J. Chloroplast capture and intra- and inter-continental biogeographic diversification in the Asian—New World disjunct plant genus Osmorhiza (Apiaceae). Mol. Phylogenet. Evol. 2015, 85, 10–21. [Google Scholar] [CrossRef]
- Casazza, G.; Granato, L.; Minuto, L.; Conti, E. Polyploid evolution and Pleistocene glacial cycles: A case study from the alpine primrose Primula marginata (Primulaceae). BMC Evol. Biol. 2012, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Cristina Acosta, M.; Premoli, A.C. Evidence of chloroplast capture in South American Nothofagus (subgenus Nothofagus, Nothofagaceae). Mol. Phylogenet. Evol. 2010, 54, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.F.; He, C.H.; Peng, C.I.; Hu, C.M.; Hao, G. Circumscription of Primula subgenus Auganthus (Primulaceae) based on chloroplast DNA sequences. J. Syst. Evol. 2010, 48, 123–132. [Google Scholar] [CrossRef]
- Liu, Y.J.; Liu, J.; Hu, C.M.; Hao, G. Non-monophyly of Primula subgenera Auganthus and Carolinella (Primlaceae) as confirmed by the nuclear DNA sequence variation. Plant Syst. Evol. 2015, 301, 2057–2071. [Google Scholar] [CrossRef]
- Ren, T.; Yang, Y.; Zhou, T.; Liu, Z.L. Comparative plastid genomes of Primula species: Sequence divergence and phylogenetic relationships. Int. J. Mol. Sci. 2018, 19, 1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trift, I.; Källersjö, M.; Anderberg, A.A. The monophyly of Primula (Primulaceae) evaluated by analysis of sequences from the chloroplast gene rbcL. Syst. Bot. 2002, 27, 396–407. [Google Scholar] [CrossRef]
- Mast, A.R.; Kelso, S.; Conti, E. Are any primroses (Primula) primitively monomorphic? New Phytol. 2006, 171, 605–616. [Google Scholar] [CrossRef]
- Ren, G.; Conti, E.; Salamin, N. Phylogeny and biogeography of Primula sect. Armerina: Implications for plant evolution under climate change and the uplift of the Qinghai-Tibet Plateau. BMC Evol. Biol. 2015, 15, 161. [Google Scholar] [CrossRef] [Green Version]
- Conti, E.; Suring, E.; Boyd, D.; Jorgensen, J.; Grant, J.; Kelso, S. Phylogenetic relationships and character evolution in primula L.: The usefulness of ITS sequence data. Plant Biosyst. 2000, 134, 385–392. [Google Scholar] [CrossRef]
- Hao, G.; Hu, C.M.; Lee, N.S. Circumscriptions and phylogenetic relationships of Primula sects. Auganthus and Ranunculoides: Evidence from nrDNA ITS sequences. Acta Bot. Sin. 2002, 44, 72–75. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q. The Complete Chloroplast Genomes of Primula obconica Provide Insight That Neither Species nor Natural Section Represent Monophyletic Taxa in Primula (Primulaceae). Genes 2022, 13, 567. https://doi.org/10.3390/genes13040567
Li Q. The Complete Chloroplast Genomes of Primula obconica Provide Insight That Neither Species nor Natural Section Represent Monophyletic Taxa in Primula (Primulaceae). Genes. 2022; 13(4):567. https://doi.org/10.3390/genes13040567
Chicago/Turabian StyleLi, Qiang. 2022. "The Complete Chloroplast Genomes of Primula obconica Provide Insight That Neither Species nor Natural Section Represent Monophyletic Taxa in Primula (Primulaceae)" Genes 13, no. 4: 567. https://doi.org/10.3390/genes13040567
APA StyleLi, Q. (2022). The Complete Chloroplast Genomes of Primula obconica Provide Insight That Neither Species nor Natural Section Represent Monophyletic Taxa in Primula (Primulaceae). Genes, 13(4), 567. https://doi.org/10.3390/genes13040567