Selection of the Reference Gene for Expression Normalization in Salsola ferganica under Abiotic Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA Extraction and cDNA Synthesis
2.3. PCR Primer Design
2.4. Detection of Amplification Efficiency and Selection of Reference Genes
2.5. qRT-PCR Analysis
2.6. Gene Expression Stability Analysis
2.7. Stability Evaluation of Candidate Genes
3. Results
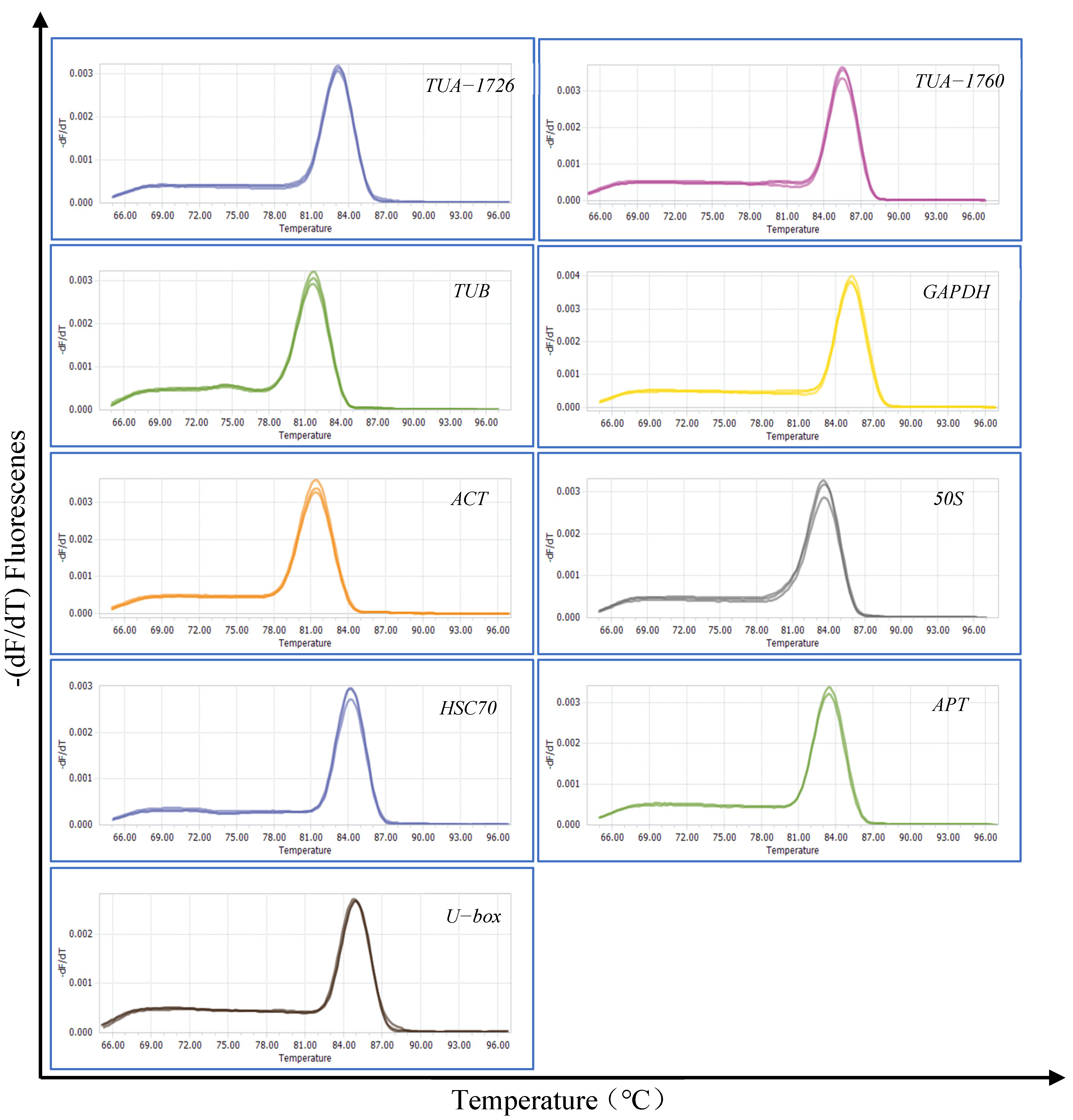
3.1. Primer Specificity and Amplification Efficiency of Candidate Reference Genes
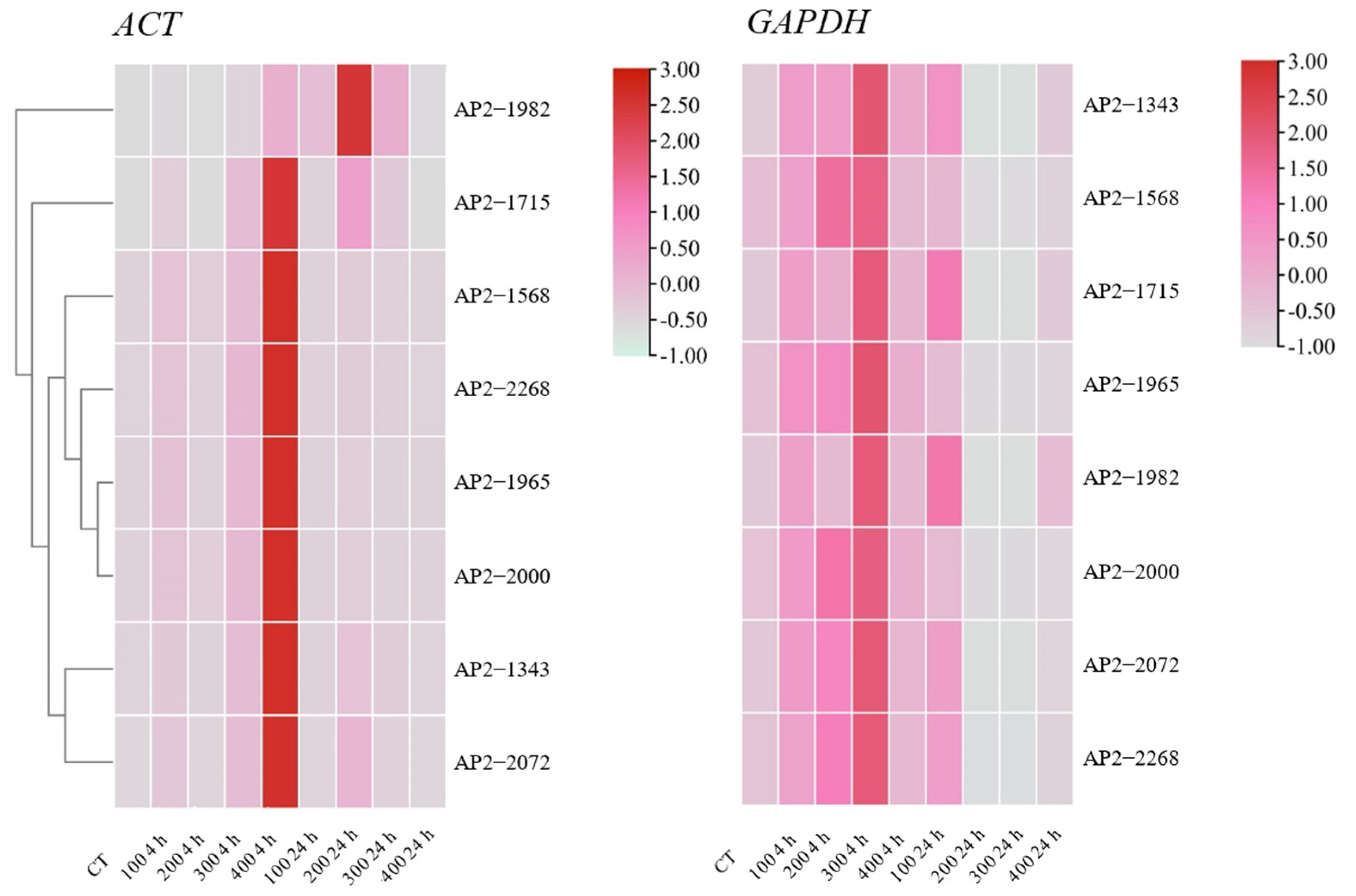
3.2. Relative Expression of Candidate Genes in Different Treatments
3.3. Stability Evaluation of Candidate Genes
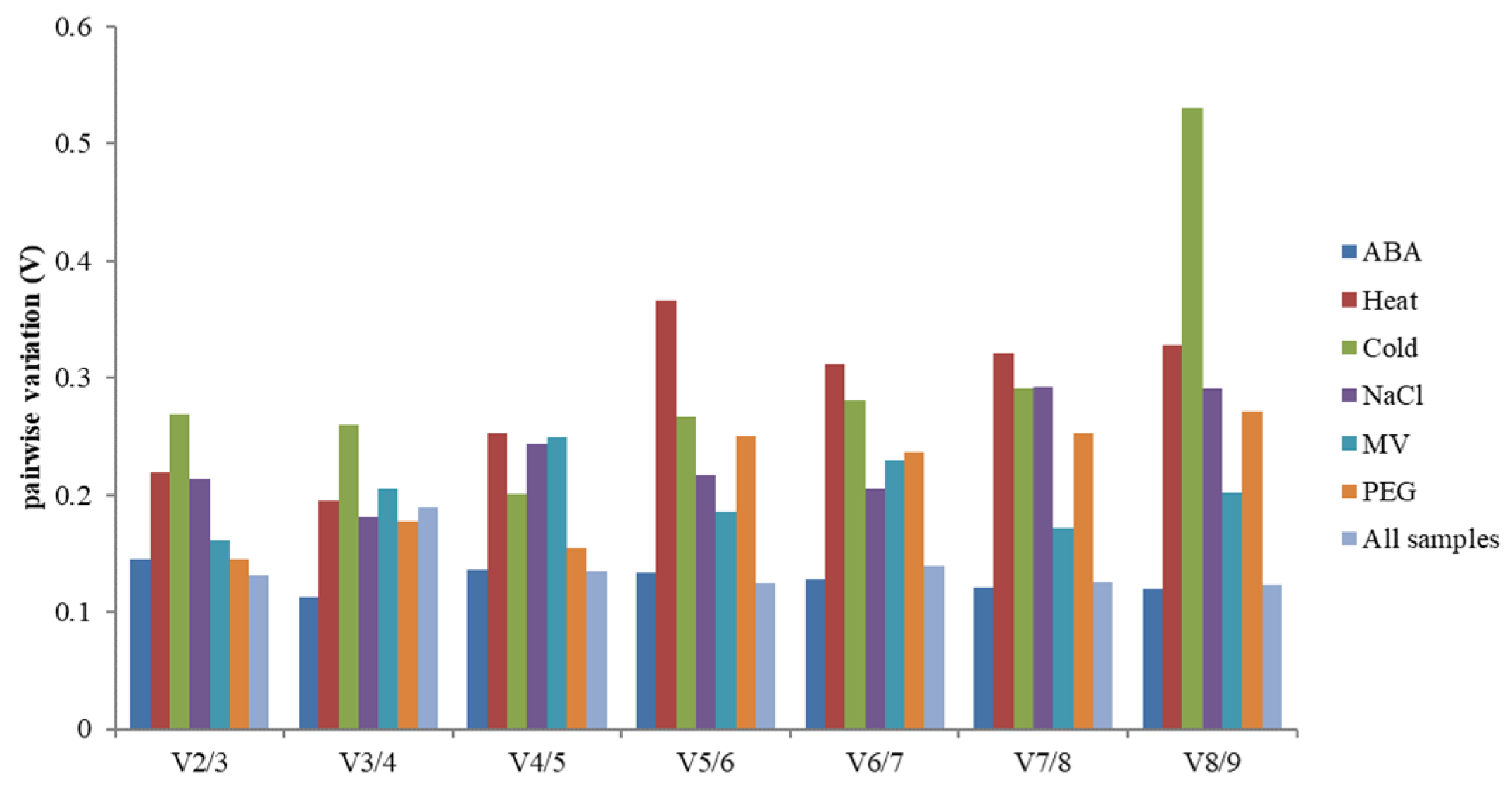
3.3.1. geNorm Analysis
3.3.2. NormFinder Analysis
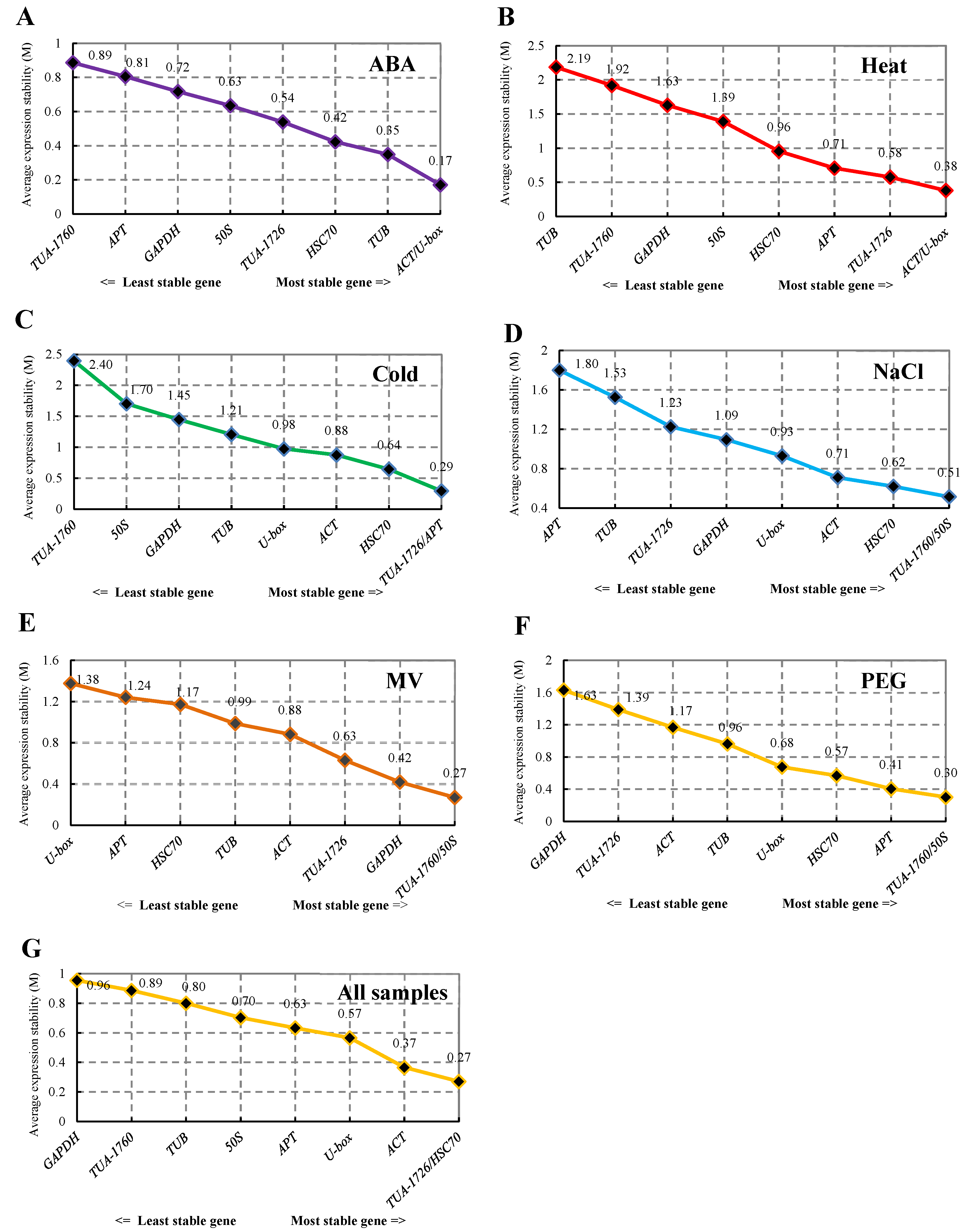
3.3.3. BestKeeper Analysis
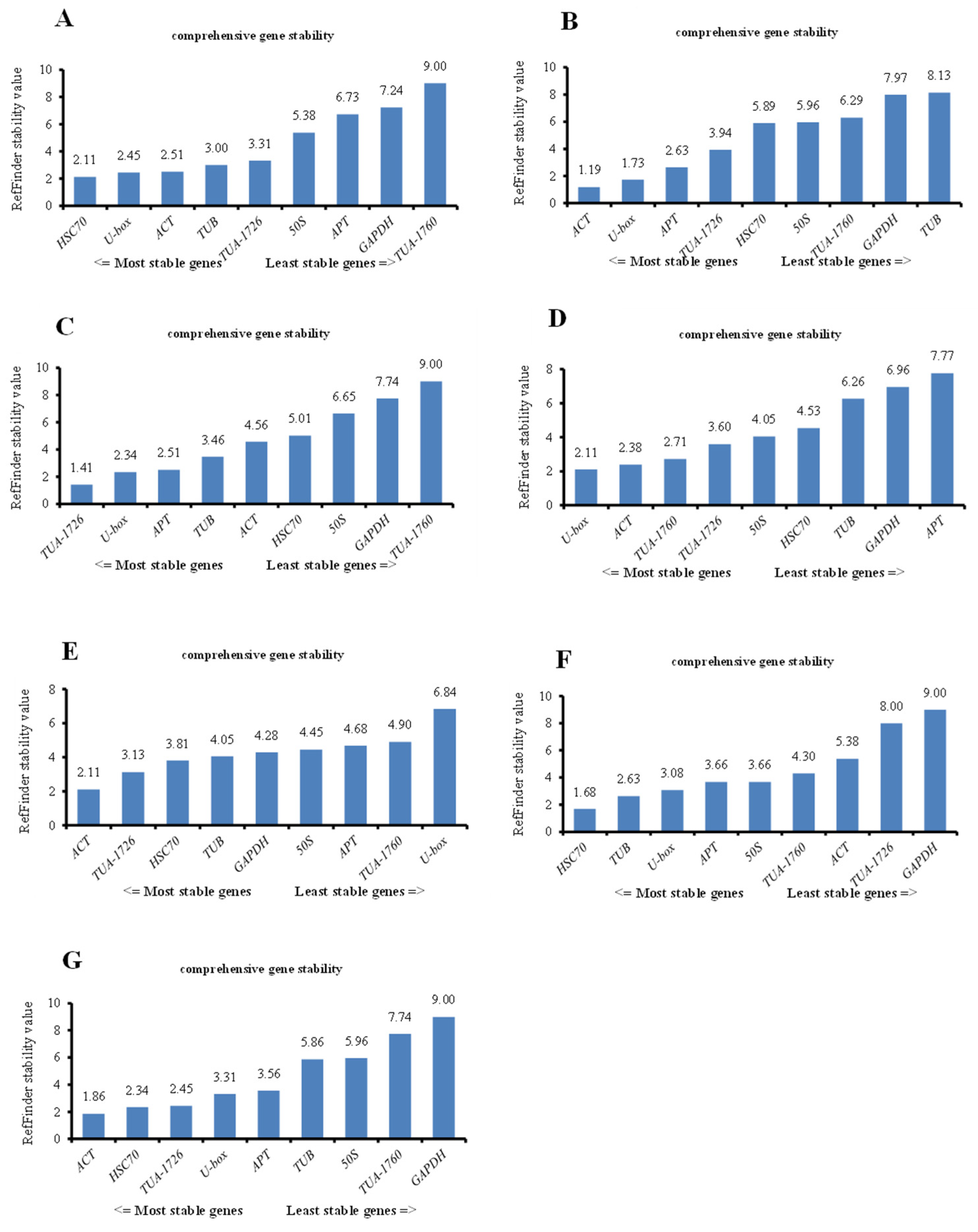
3.3.4. RefFinder Analysis
3.4. Validation of the Best- and Worst-Ranked Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, H.B.; Zhang, F.C. Advances in research on the molecular mechanism of plant salinity tolerance and morphological characters of Chenopodiaceae. Biotechnol. Bull. 2008, 4, 22–26. [Google Scholar]
- Du, H.; Zhou, L.G.; Li, C.; Sui, P. Recent studies on the chemistry and bioactivity of Chenopodiaceous plants. Nat. Prod. Res. Dev. 2007, 19, 884–889. [Google Scholar]
- Ma, Y.; Zhang, J.; Li, X.; Zhang, S.; Lan, H. Effects of environmental stress on seed germination and seedling growth of Salsola ferganica (Chenopodiaceae). Acta Ecol. Sin. 2016, 36, 456–463. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, G.Q.; Han, Y.N.; Liu, M.M. Effects of salt, alkali and salt–alkali mixed stresses on seed germination of the halophyte Salsola ferganica (Chenopodiaceae). Acta Ecol. Sin. 2013, 33, 354–360. [Google Scholar] [CrossRef]
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar] [CrossRef]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Eamens, A.L.; Grof, C.P.L. Reference gene identification for reliable normalisation of quantitative RT-PCR data in Setaria viridis. Plant. Methods 2018, 14, 24. [Google Scholar] [CrossRef]
- Wang, B.; Du, H.; Yao, Z.; Ren, C.; Ma, L.; Wang, J.; Zhang, H.; Ma, H. Validation of reference genes for accurate normalization of gene expression with quantitative real-time PCR in Haloxylon ammodendron under different abiotic stresses. Physiol. Mol. Biol. Plants 2018, 24, 455–463. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, C.; Zhang, J.; Chen, F.; Gong, Y.; Li, Y.; Su, Y.; Wei, Y.; Zhao, Y. Selection of the reference gene for expression normalization in Papaver somniferum L. under abiotic stress and hormone treatment. Genes 2020, 11, 124. [Google Scholar]
- Zhou, Y.; Xia, H.; Liu, X.; Lin, Z.; Guo, Y.; Deng, H.; Wang, J.; Lin, L.; Deng, Q.; Lv, X.; et al. Identification of Suitable Reference Genes for qRT-PCR Normalization in Kiwifruit. Horticulturae 2022, 8, 170. [Google Scholar] [CrossRef]
- Sudhakar Reddy, P.; Srinivas Reddy, D.; Sivasakthi, K.; Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Evaluation of Sorghum [Sorghum bicolor (L.)] reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef]
- Yang, R.W. Molecular Biology; Nanjing University Press: Nanjing, China, 2007. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.G.; Ye, X.Y.; Xing, S.Q.; Hou, H.Y. Study on real time fluorescence quantitative PCR detection technology of potato leaf roll virus. Hunan Agric. Sci. 2018, 9, 9–12. [Google Scholar]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, S.; Jiang, W.; Huang, Y.; Bie, Z. Identification of suitable reference genes for gene expression normalization in qRT-PCR analysis in watermelon. PLoS ONE 2014, 9, e90612. [Google Scholar]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Front. Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.-Y.; Chen, J.-W.; Li, C.-Q.; Chen, L.; Wu, J.-Y.; Chen, J.-Y.; Lu, W.-J.; Li, J.-G. Selection of reliable reference genes for expression studies by reverse transcription quantitative real-time PCR in litchi under different experimental conditions. Plant Cell Rep. 2011, 30, 641–653. [Google Scholar] [CrossRef]
- Lv, Y.; Li, Y.; Liu, X.; Xu, K. Identification of ginger (Zingiber officinale Roscoe) reference genes for gene expression analysis. Front. Genet. 2020, 11, 586098. [Google Scholar] [CrossRef]
- Li, C.; Xu, J.; Deng, Y.; Sun, H.; Li, Y. Selection of reference genes for normalization of cranberry (Vaccinium macrocarpon Ait.) gene expression under different experimental conditions. PLoS ONE 2019, 14, e0224798. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-G.; Wang, X.-P.; Li, X.-R.; Zhang, Z.-S.; Wang, G.; Wang, S.; Ma, F.-Y. Advances and prospect of researches on desert plant life history strategies. J. Desert Res. 2005, 25, 306–314. [Google Scholar]
- Tian, J.; Wang, Y.; Yan, S.; Sun, S.; Jia, J.; Hu, X. Metabolomics technology and its applications in agricultural animal and plant research. Hereditas 2020, 42, 452–465. [Google Scholar] [PubMed]
- Bouton, J. The economic benefits of forage improvement in the United States. Euphytica 2007, 154, 263–270. [Google Scholar] [CrossRef]
- Xiao, F.; Zheng, Y.; Chen, J.; Zhao, C.; Chen, H.; Wang, L.; Liu, S. Selection and validation of reference genes in all-red Amaranth (Amaranthus tricolor L.) seedlings under different culture conditions. J. Hortic. Sci. Biotechnol. 2021, 96, 604–613. [Google Scholar] [CrossRef]
- Cheng, T.; Zhu, F.; Sheng, J.; Zhao, L.; Zhou, F.; Hu, Z.; Diao, Y.; Jin, S. Selection of suitable reference genes for quantitive real-time PCR normalization in Miscanthus lutarioriparia. Mol. Biol. Rep. 2019, 46, 4545–4553. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, L.; Wang, X.; Liu, H.; Tian, H.; Wang, J. Selection of reliable reference genes for gene expression analysis in seeds at different developmental stages and across various tissues in Paeonia ostii. Mol. Biol. Rep. 2019, 46, 6003–6011. [Google Scholar] [CrossRef]
- Jaiswal, P.; Kaur, N.; Randhawa, G. Identification of reference genes for real-time PCR gene expression studies during seed development and under abiotic stresses in Cyamopsis tetragonoloba (L.) Taub. bioRxiv 2018, 313437. [Google Scholar]
- Wen, Z.B.; Zhang, M.L. Reference gene selection for real-time quantitative PCR in Salsola laricifolia under soil drought stress. Plant Physiol. J. 2015, 51, 2031–2038. [Google Scholar]
- Yeap, W.C.; Loo, J.M.; Wong, Y.C.; Kulaveerasingam, H. Evaluation of suitable reference genes for qRT-PCR gene expression normalization in reproductive, vegetative tissues and during fruit development in oil palm. Plant Cell Tissue Organ Cult. PCTOC 2014, 116, 55–66. [Google Scholar] [CrossRef]
- Chen, M.-D.; Wang, B.; Li, Y.-P.; Zeng, M.-J.; Liu, J.-T.; Ye, X.-R.; Zhu, H.-S.; Wen, Q.-F. Reference gene selection for qRT-PCR analyses of luffa (Luffa cylindrica) plants under abiotic stress conditions. Sci. Rep. 2021, 11, 3161. [Google Scholar] [CrossRef]
- Guénin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhang, R.; Zhou, Z. Identification and Validation of Reference Genes for Gene Expression Analysis in Schima superba. Genes 2021, 12, 732. [Google Scholar] [CrossRef]
- Chandna, R.; Augustine, R.; Bisht, N.C. Evaluation of candidate reference genes for gene expression normalization in Brassica juncea using real time quantitative RT-PCR. PLoS ONE 2012, 7, e36918. [Google Scholar] [CrossRef]
- Xu, Z.; Dai, J.; Su, W.; Wu, H.; Shah, K.; Xing, L.; Ma, J.; Zhang, D.; Zhao, C. Selection and Validation of Reliable Reference Genes for Gene Expression Studies in Different Genotypes and TRV-Infected Fruits of Peach (Prunus persica L. Batsch) during Ripening. Genes 2022, 13, 160. [Google Scholar] [CrossRef]
- Moazzam Jazi, M.; Ghadirzadeh Khorzoghi, E.; Botanga, C.; Seyedi, S.M. Identification of reference genes for quantitative gene expression studies in a non-model tree Pistachio (Pistacia vera L.). PLoS ONE 2016, 11, e0157467. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhou, H.; Nie, Z.; Wang, X.; Luo, B.; Yi, Z.; Li, X.; Hu, X.; Yang, T. Appropriate reference genes for RT-qPCR normalization in various organs of Anemone flaccida Fr. Schmidt at different growing stages. Genes 2021, 12, 459. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, C.-S.; Huo, Y.-B.; Zhang, B.; Ma, Z.-Q.; Feng, J.-T.; Zhang, X. Validation of suitable reference genes for quantitative gene expression analysis in Tripterygium wilfordii. Mol. Biol. Rep. 2019, 46, 4161–4174. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, C.; Yang, H.; Lyu, L.; Li, W.; Wu, W. Selection and Validation of Candidate Reference Genes for Gene Expression Analysis by RT-qPCR in Rubus. Int. J. Mol. Sci. 2021, 22, 10533. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Z.; Huang, H.; Wei, S.; Liu, Y.; Jiang, C.; Zhang, J.; Zhang, C. Selection of relatively exact reference genes for gene expression studies in goosegrass (Eleusine indica) under herbicide stress. Sci. Rep. 2017, 7, 46494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraji, S.; Filiz, E.; Kazemitabar, S.K.; Vannozzi, A.; Palumbo, F.; Barcaccia, G.; Heidari, P. The AP2/ERF gene family in Triticum durum: Genome-wide identification and expression analysis under drought and salinity stresses. Genes 2020, 11, 1464. [Google Scholar] [CrossRef]
- Do, T.H.; Pongthai, P.; Ariyarathne, M.; Teh, O.-K.; Fujita, T. AP2/ERF transcription factors regulate salt-induced chloroplast division in the moss Physcomitrella patens. J. Plant Res. 2020, 133, 537–548. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Ngatia, J.N.; Wang, Y.; Khoso, M.A.; Farooq, U.; Chen, S. AP2/ERF, an important cold stress-related transcription factor family in plants: A review. Physiol. Mol. Biol. Plants 2021, 27, 1953–1968. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Primer Sequences (5’→3’) | Amplification Length/bp | Amplification Efficiency/% | R2 |
|---|---|---|---|---|---|
| TUA-1726 | α-tubulin | GTGGCACTGGTTCTGGACTTG | 108 | 98.35 | 0.9977 |
| TTGAAACTTGAGGAGACGGGTAA | |||||
| TUA-1760 | TCCGCAAGCTCGCTGATA | 161 | 105.82 | 0.9992 | |
| GGGAGATGGGTAGATGGTGAA | |||||
| TUB | β-tubulin | TTACACTGAGGGTGCCGAAC | 92 | 90.94 | 0.9995 |
| AAACCTGGAATCCTTGAAGACA | |||||
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | CCATCCTCGGCACATTCAAC | 146 | 102.21 | 0.9934 |
| TCCTTCAATCACCAAGTCTACGC | |||||
| ACT | Actin | TTCATCGGAGACGAAGCAGTAG | 107 | 97.89 | 0.9996 |
| AACCTTTCCATAGCATCCCAGT | |||||
| 50S | 50S ribosomal protein | TTGCTAAGCCTGGTTGCATC | 138 | 95.11 | 0.9998 |
| TGTCAGGACCAAACTTCTCAAAT | |||||
| HSC 70 | Heat shock protein 70 | CCAATGACAAGGGTAGGCTCT | 141 | 101.78 | 0.9991 |
| TCCTCATGTTGTAGGCGTAGTTC | |||||
| APT | Adenine phosphoribosyl transferase-like protein | AAGGCTGAAGTGGCTGAATGT | 127 | 105.85 | 0.9908 |
| TCCTTAAACGGCAGTCTTCTAACT | |||||
| U-box | U-box domain-containing protein | AACACTTGATTCACGCACCCA | 143 | 95.76 | 0.9921 |
| TTGCTTCCATGCTGCCTTTC |
| Rank | ABA | Heat | Cold | NaCl | MV | PEG | All Samples | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | HSC70 | 0.126 | ACT | 0.190 | ACT | 0.248 | TUB | 0.374 | ACT | 0.264 | TUA-1726 | 0.390 | U-box | 0.246 |
| 2 | TUA-1726 | 0.173 | APT | 0.325 | TUA-1726 | 0.326 | HSC70 | 0.590 | TUA-1726 | 0.403 | U-box | 0.629 | ACT | 0.295 |
| 3 | TUB | 0.512 | U-box | 0.385 | TUB | 0.349 | U-box | 0.717 | HSC70 | 0.437 | TUB | 0.671 | TUA-1760 | 0.767 |
| 4 | 50S | 0.595 | TUA-1726 | 1.029 | GAPDH | 1.050 | ACT | 0.893 | APT | 0.586 | APT | 0.743 | TUA-1726 | 1.158 |
| 5 | ACT | 0.617 | 50S | 1.930 | APT | 1.058 | APT | 1.200 | 50S | 0.596 | 50S | 1.534 | HSC70 | 1.197 |
| 6 | U-box | 0.733 | HSC70 | 1.939 | HSC70 | 1.068 | 50S | 1.279 | U-box | 0.642 | ACT | 1.576 | 50S | 1.255 |
| 7 | GAPDH | 0.776 | TUA-1760 | 2.092 | 50S | 1.386 | TUA-1760 | 1.610 | TUA-1760 | 0.984 | HSC70 | 1.604 | GAPDH | 1.619 |
| 8 | APT | 0.956 | GAPDH | 2.301 | TUA-1760 | 1.529 | TUA-1726 | 1.629 | TUB | 1.018 | GAPDH | 2.249 | TUB | 1.955 |
| 9 | TUA-1760 | 1.061 | TUB | 2.903 | U-box | 1.785 | GAPDH | 2.415 | GAPDH | 1.089 | TUA-1760 | 4.746 | APT | 2.586 |
| Rank | ABA | Heat | Cold | NaCl | MV | PEG | All Samples | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | |
| 1 | U-box | 0.507 | 1.434 | U-box | 0.646 | 1.842 | U-box | 0.530 | 1.484 | TUA-1726 | 0.720 | 2.263 | HSC70 | 0.212 | 0.807 | HSC70 | 0.520 | 1.924 | U-box | 0.206 | 0.588 |
| 2 | ACT | 0.513 | 1.639 | ACT | 0.766 | 2.463 | TUB | 0.719 | 2.195 | U-box | 0.755 | 2.089 | APT | 0.227 | 0.700 | U-box | 0.527 | 1.534 | APT | 0.411 | 1.242 |
| 3 | TUB | 0.574 | 1.740 | APT | 0.858 | 2.631 | ACT | 1.068 | 3.401 | TUB | 0.793 | 2.505 | U-box | 0.507 | 1.509 | APT | 0.705 | 2.154 | TUB | 0.428 | 1.333 |
| 4 | APT | 0.738 | 2.214 | TUA-1760 | 1.113 | 3.785 | TUA-1726 | 1.302 | 3.928 | ACT | 1.300 | 4.056 | ACT | 0.540 | 1.726 | TUB | 0.711 | 2.195 | ACT | 0.574 | 1.827 |
| 5 | HSC70 | 0.785 | 2.809 | TUA-1726 | 1.198 | 3.717 | APT | 1.331 | 3.846 | APT | 1.583 | 4.826 | TUB | 0.562 | 1.713 | 50S | 0.958 | 3.127 | HSC70 | 0.821 | 2.926 |
| 6 | TUA-1726 | 1.047 | 3.187 | TUB | 1.473 | 4.914 | HSC70 | 1.608 | 5.487 | TUA-1760 | 1.705 | 5.061 | TUA-1726 | 0.964 | 2.950 | ACT | 1.011 | 3.244 | TUA-1726 | 0.862 | 2.656 |
| 7 | 50S | 1.185 | 3.624 | 50S | 1.760 | 5.731 | 50S | 1.778 | 5.516 | HSC70 | 1.709 | 5.719 | GAPDH | 1.368 | 4.250 | TUA-1760 | 1.087 | 3.545 | 50S | 1.002 | 3.099 |
| 8 | GAPDH | 1.389 | 4.311 | HSC70 | 1.862 | 6.693 | GAPDH | 2.303 | 6.765 | GAPDH | 1.796 | 5.440 | 50S | 1.608 | 4.793 | TUA-1726 | 1.483 | 4.615 | TUA-1760 | 1.090 | 3.402 |
| 9 | TUA-1760 | 1.515 | 4.550 | GAPDH | 1.887 | 6.377 | TUA-1760 | 3.473 | 11.036 | 50S | 1.941 | 5.672 | TUA-1760 | 1.708 | 5.070 | GAPDH | 1.987 | 6.425 | GAPDH | 1.276 | 3.989 |
| Gene | ACT | U-Box | HSC70 | TUA-1726 | TUB | APT | 50S | TUA-1760 | GAPDH |
|---|---|---|---|---|---|---|---|---|---|
| ABA | 3 | 2 | 1 | 5 | 4 | 7 | 6 | 9 | 8 |
| Heat | 1 | 2 | 5 | 4 | 9 | 3 | 6 | 7 | 8 |
| Cold | 5 | 2 | 6 | 1 | 4 | 3 | 7 | 9 | 8 |
| NaCl | 2 | 1 | 7 | 4 | 6 | 9 | 5 | 3 | 8 |
| MV | 1 | 9 | 3 | 2 | 4 | 7 | 6 | 8 | 5 |
| PEG | 7 | 3 | 1 | 8 | 2 | 4 | 5 | 6 | 9 |
| All samples | 1 | 4 | 2 | 3 | 6 | 5 | 7 | 8 | 9 |
| Mean | 2.857 | 3.286 | 3.571 | 3.857 | 5.000 | 5.429 | 6.000 | 7.143 | 7.857 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhang, S. Selection of the Reference Gene for Expression Normalization in Salsola ferganica under Abiotic Stress. Genes 2022, 13, 571. https://doi.org/10.3390/genes13040571
Wang S, Zhang S. Selection of the Reference Gene for Expression Normalization in Salsola ferganica under Abiotic Stress. Genes. 2022; 13(4):571. https://doi.org/10.3390/genes13040571
Chicago/Turabian StyleWang, Shuran, and Sheng Zhang. 2022. "Selection of the Reference Gene for Expression Normalization in Salsola ferganica under Abiotic Stress" Genes 13, no. 4: 571. https://doi.org/10.3390/genes13040571
APA StyleWang, S., & Zhang, S. (2022). Selection of the Reference Gene for Expression Normalization in Salsola ferganica under Abiotic Stress. Genes, 13(4), 571. https://doi.org/10.3390/genes13040571