
Selection and Validation of Reference Genes for RT-qPCR Analysis in Tibetan Medicinal Plant Saussurea Laniceps Callus under Abiotic Stresses and Hormone Treatments

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Total RNA Extraction and Complementary DNA Synthesis
2.3. Designing Primer and Testing Performance
2.4. Data Analysis and Evaluation
2.5. Validation of Selected Candidate Reference Genes
2.6. Statistical Analysis
3. Results
3.1. Evaluation of Primer Specificity and Effectiveness
3.2. Expression Distribution of Candidate Reference Genes
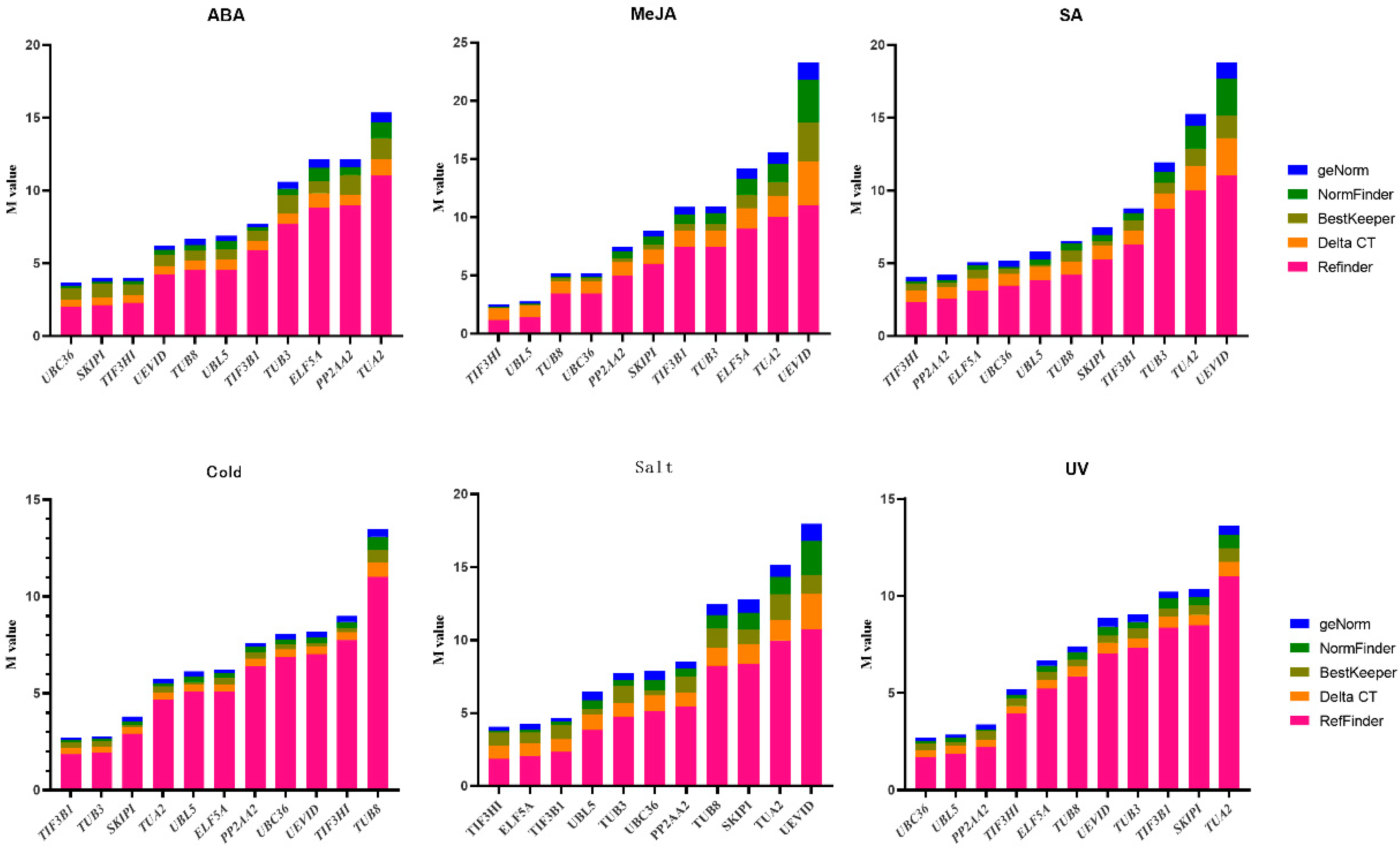
3.3. Candidate Reference Gene Expression Stability
3.4. Validation of Candidate Reference Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chik, W.I.; Zhu, L.; Fan, L.L.; Yi, T.; Zhu, G.Y.; Gou, X.J.; Tang, Y.N.; Xu, J.; Yeung, W.P.; Zhao, Z.Z.; et al. Saussurea involucrata: A review of the botany, phytochemistry and ethnopharmacology of a rare traditional herbal medicine. J. Ethnopharmacol. 2015, 172, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Byambaragchaa, M.; de la Cruz, J.; Yang, S.H.; Hwang, S.G. Anti-metastatic potential of ethanol extract of Saussurea involucrata against hepatic cancer in vitro. Asian Pac. J. Cancer Prev. 2013, 14, 5397–5402. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jin, D.; Lu, C.; Lan, X.; Qiao, P.; Li, H.; Chen, Y. Proteomic responses associated with freezing tolerance in the callus of the Tibetan alpine plant Saussurea laniceps during cold acclimation. Plant Cell Tissue Organ Cult. 2016, 124, 81–95. [Google Scholar] [CrossRef]
- Yu, M.; Chen, Y.; Lan, X.; Jin, D.; Zhang, J.; Lu, C. Effects of cold acclimation on proteome expression patterns related to freezing tolerance in a Tibetan alpine plant Saussurea laniceps. Plant Omics 2014, 7, 221–228. [Google Scholar]
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real time quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef]
- Tong, J.; Hu, M.; Han, B.; Ji, Y.; Wang, B.; Liang, H.; Liu, M.; Wu, Z.; Liu, N. Determination of reliable reference genes for gene expression studies in Chinese chive (Allium tuberosum) based on the transcriptome profiling. Sci. Rep. 2021, 11, 16558. [Google Scholar] [CrossRef]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef]
- Wood, S.H.; Clements, D.N.; McEwan, N.A.; Nuttall, T.; Carter, S.D. Reference genes for canine skin when using quantitative real-time PCR. Vet. Immunol. Immunopathol. 2008, 126, 392–395. [Google Scholar] [CrossRef]
- Gutierrez, L.; Mauriat, M.; Guenin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef]
- Artico, S.; Nardeli, S.M.; Brilhante, O.; Grossi-de-Sa, M.F.; Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutumfor accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol. 2010, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.Q.; Eamens, A.L.; Grof, C.P.L. Reference gene identification for reliable normalisation of quantitative RT-PCR data in Setaria viridis. Plant Methods 2018, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.K.; Mafra, V.; de Souza, W.R.; Ribeiro, A.P.; Vinecky, F.; Basso, M.F.; da Cunha, B.A.; Kobayashi, A.K.; Molinari, H.B. Selection of reliable reference genes for RT-qPCR analysis during developmental stages and abiotic stress in Setaria viridis. Sci. Rep. 2016, 6, 28348. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef]
- Bustin, S. Invited review Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, J.; Eattock, N.; Van Deynze, A.; Blumwald, E. Selection and validation of reference genes for gene expression analysis in switchgrass (Panicum virgatum) using quantitative real-time RT-PCR. PLoS ONE 2014, 9, e91474. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Zhu, B.; Cai, J.; Li, P.; Wang, L.; Dai, H.; Qiu, L.; Yu, H.; Ha, D.; Zhao, H.; et al. Selection of reference genes for gene expression studies in Siberian Apricot (Prunus sibirica L.) Germplasm using quantitative real-time PCR. PLoS ONE 2014, 9, e103900. [Google Scholar] [CrossRef]
- Wang, M.; Lu, S. Validation of suitable reference genes for quantitative gene expression analysis in Panax ginseng. Front. Plant Sci. 2016, 6, 1259. [Google Scholar] [CrossRef] [PubMed]
- Mallona, I.; Lischewski, S.; Weiss, J.; Hause, B.; Egea-Cortines, M. Validation of reference genes for quantitative real-time PCR during leaf and flower development in Petunia hybrida. BMC Plant Biol. 2010, 10, 4. [Google Scholar] [CrossRef]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Leg. Med. 2012, 126, 943–952. [Google Scholar] [CrossRef]
- Molomjamts, M.; Ingolfsland, E. Identification of reference genes for the normalization of retinal gene expression in oxygen induced retinopathy, anemia and erythropoietin administration. Investig. Ophthalmol. Vis. Sci. 2021, 62, 2930. [Google Scholar]
- Zhao, Z.; Zhou, H.; Nie, Z.; Wang, X.; Luo, B.; Yi, Z.; Li, X.; Hu, X.; Yang, T. Appropriate reference genes for RT-qPCR normalization in various organs of Anemone flaccida Fr. Schmidt at different growing stages. Genes 2021, 12, 459. [Google Scholar] [CrossRef]
- Saha, G.C.; Vandemark, G.J. Evaluation of Expression Stability of Candidate References Genes among Green and Yellow Pea Cultivars (Pisum sativum L.) Subjected to Abiotic and Biotic Stress. Am. J. Plant Sci. 2012, 3, 235–242. [Google Scholar] [CrossRef]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of reference genes for quantitative gene expression studies in Platycladus orientalis (Cupressaceae) Using real-time PCR. PLoS ONE 2012, 7, e33278. [Google Scholar] [CrossRef]
- Lovdal, T.; Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem. 2009, 387, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Wan, Y.; Yang, Q.; Zou, B.; Ren, W.; Ding, Y.; Wang, Z.; Wang, R.; Wang, K.; Hou, X. Selection of reference genes for qRT-PCR analysis of gene expression in stipa grandis during environmental stresses. PLoS ONE 2017, 12, e0169465. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, J.; Li, G.; Qi, L.; Yang, F.; Wang, R.; Li, G. Reference gene selection for qRT-PCR in Caragana korshinskii Kom. under different stress conditions. Mol. Biol. Rep. 2014, 41, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Bekker-Jensen, S.; Mailand, N. Ubiquitin-like protein UBL5 promotes the functional integrity of the Fanconi anemia pathway. EMBO J. 2015, 34, 1385–1398. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Roche, J.R.; Donaghy, D.J.; Thrush, A.; Sathish, P. Validation of reference genes for quantitative RT-PCR studies of gene expression in perennial ryegrass (Lolium perenne L.). BMC Mol. Biol. 2010, 11, 8. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, C.; Zhang, J.; Chen, F.; Gong, Y.; Li, Y.; Su, Y.; Wei, Y.; Zhao, Y. Selection of the reference gene for expression normalization in Papaver somniferum L. under abiotic stress and hormone treatment. Genes 2020, 11, 124. [Google Scholar] [CrossRef]
- Wang, B.; Du, H.; Yao, Z.; Ren, C.; Ma, L.; Wang, J.; Zhang, H.; Ma, H. Validation of reference genes for accurate normalization of gene expression with quantitative real-time PCR in Haloxylon ammodendron under different abiotic stresses. Physiol. Mol. Biol. Plants 2018, 24, 455–463. [Google Scholar] [CrossRef]
- Cordoba, E.M.; Die, J.V.; Gonzalez-Verdejo, C.I.; Nadal, S.; Roman, B. Selection of reference genes in Hedysarum coronarium under various stresses and stages of development. Anal. Biochem. 2011, 409, 236–243. [Google Scholar] [CrossRef]
- Li, L.; Li, N.; Fang, H.; Qi, X.; Zhou, Y. Selection and validation of reference genes for normalisation of gene expression in Glehnia littoralis. Sci. Rep. 2020, 10, 7374. [Google Scholar] [CrossRef]
- Lilly, S.; Drummond, R.; Pearson, M.; MacDiarmid, R. Identification and validation of reference genes for normalization of transcripts from virus-infected Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2011, 24, 294–304. [Google Scholar] [CrossRef]
- Wen, R.; Torres-Acosta, J.A.; Pastushok, L.; Lai, X.; Pelzer, L.; Wang, H.; Xiao, W. Arabidopsis UEV1D promotes Lysine-63-linked polyubiquitination and is involved in DNA damage response. Plant Cell 2008, 20, 213–227. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Primer Sequence (5′-3′) | Product (bp) | E (%) | R2 | Arabidopsis Ortholog | |
|---|---|---|---|---|---|---|---|
| Forward/Reverse | |||||||
| TUA2 | Tubulinalp α-2 chain | F-TCAGGCCGGGGTATTCAGGTTG | 146 | 94.5 | 0.998 | AT1G50010 | |
| R-CAGCACCAGTTTCGCTGAAG | |||||||
| TUB3 | Tubulinalp β chain 3 | F-AGATCTGGTGCCTACGGACA | 104 | 109 | 0.999 | AT5G62700 | |
| R-TCAGCGCCTTCGGTATAGTG | |||||||
| TUB8 | Tubulinalp β 8 | F-TGGTACACTGGTGAAGGAATGG | 107 | 94.6 | 0.995 | AT5G23860 | |
| R-TCATCAGCAGTCGCATCTTGA | |||||||
| TIF3B1 | Translation initiation factor 3B | F-AATCCGAGGCCCGATGTTAG | 141 | 108 | 0.998 | AT5G27640 | |
| R-GAATCCCTTCAACCCAGCCA | |||||||
| TIF3H1 | Translation initiation factor 3 subunit H1 | F-TCGGGGAGAGGCTTGAAGAT | 141 | 101 | 0.998 | AT1G10840 | |
| R-CCGTAATCTGTCACGTCAGC | |||||||
| ELF5A | Elongation factor 5A | F-ATGTCGGACGAAGAGCATCA | 101 | 90.4 | 0.993 | AT1G69410 | |
| R-ACGATGTGACCTCCTTTGCG | |||||||
| PP2AA2 | Protein phosphatase 2A | F-TGCCTATCCTCCGCAAGTTC | 123 | 97.4 | 0.998 | AT3G25800 | |
| R-TGAGTTTCCAGATGTTCGCCT | |||||||
| UEV1D | Ubiquitin E2 variant 1D | F-GGCACTATCATTGGCCCTCA | 101 | 93.6 | 0.992 | AT3G52560 | |
| R-CGCACACTAGGAGGCCCTCTC | |||||||
| UBL5 | Ubiquitin-like protein 5 | F-GACGATACCATCGGCGACTT | 141 | 90.5 | 0.998 | AT5G42300 | |
| R-GCCCATGCCATCGTGAATCT | |||||||
| UBC36 | Ubiquitin-conjugating enzyme 36 | F-GACGAGTCCACTCCTTTGCT | 151 | 92.3 | 0.991 | AT1G16890 | |
| R-AGCCCTGCTCTTCAGATTCG | |||||||
| SKIP1 | SKP1 interaction partner 1 | F-GCAGACAAACATGGCACCTC | 106 | 96.2 | 0.997 | AT5G57900 | |
| R-CTCAGGACTCTACTCCGGCT | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Lu, Y.; Wang, X.; Wang, X.; Li, R.; Lu, C.; Lan, X.; Chen, Y. Selection and Validation of Reference Genes for RT-qPCR Analysis in Tibetan Medicinal Plant Saussurea Laniceps Callus under Abiotic Stresses and Hormone Treatments. Genes 2022, 13, 904. https://doi.org/10.3390/genes13050904
Liu H, Lu Y, Wang X, Wang X, Li R, Lu C, Lan X, Chen Y. Selection and Validation of Reference Genes for RT-qPCR Analysis in Tibetan Medicinal Plant Saussurea Laniceps Callus under Abiotic Stresses and Hormone Treatments. Genes. 2022; 13(5):904. https://doi.org/10.3390/genes13050904
Chicago/Turabian StyleLiu, Huan, Yaning Lu, Xiaojing Wang, Xiaowei Wang, Rongchen Li, Cunfu Lu, Xiaozhong Lan, and Yuzhen Chen. 2022. "Selection and Validation of Reference Genes for RT-qPCR Analysis in Tibetan Medicinal Plant Saussurea Laniceps Callus under Abiotic Stresses and Hormone Treatments" Genes 13, no. 5: 904. https://doi.org/10.3390/genes13050904
APA StyleLiu, H., Lu, Y., Wang, X., Wang, X., Li, R., Lu, C., Lan, X., & Chen, Y. (2022). Selection and Validation of Reference Genes for RT-qPCR Analysis in Tibetan Medicinal Plant Saussurea Laniceps Callus under Abiotic Stresses and Hormone Treatments. Genes, 13(5), 904. https://doi.org/10.3390/genes13050904