
Genomic Survey and Resources for the Boring Giant Clam Tridacna crocea




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Sampling and DNA Extraction
2.2. Library Preparation and Sequencing
2.3. Genome Size Estimation in Tridacna crocea
2.4. Repetitive Elements in the Nuclear Genome of Tridacna crocea
2.5. Nuclear Ribosomal Operon in Tridacna crocea
2.6. Mitochondrial Genome Assembly from RNA-seq in Tridacna crocea
3. Results and Discussion
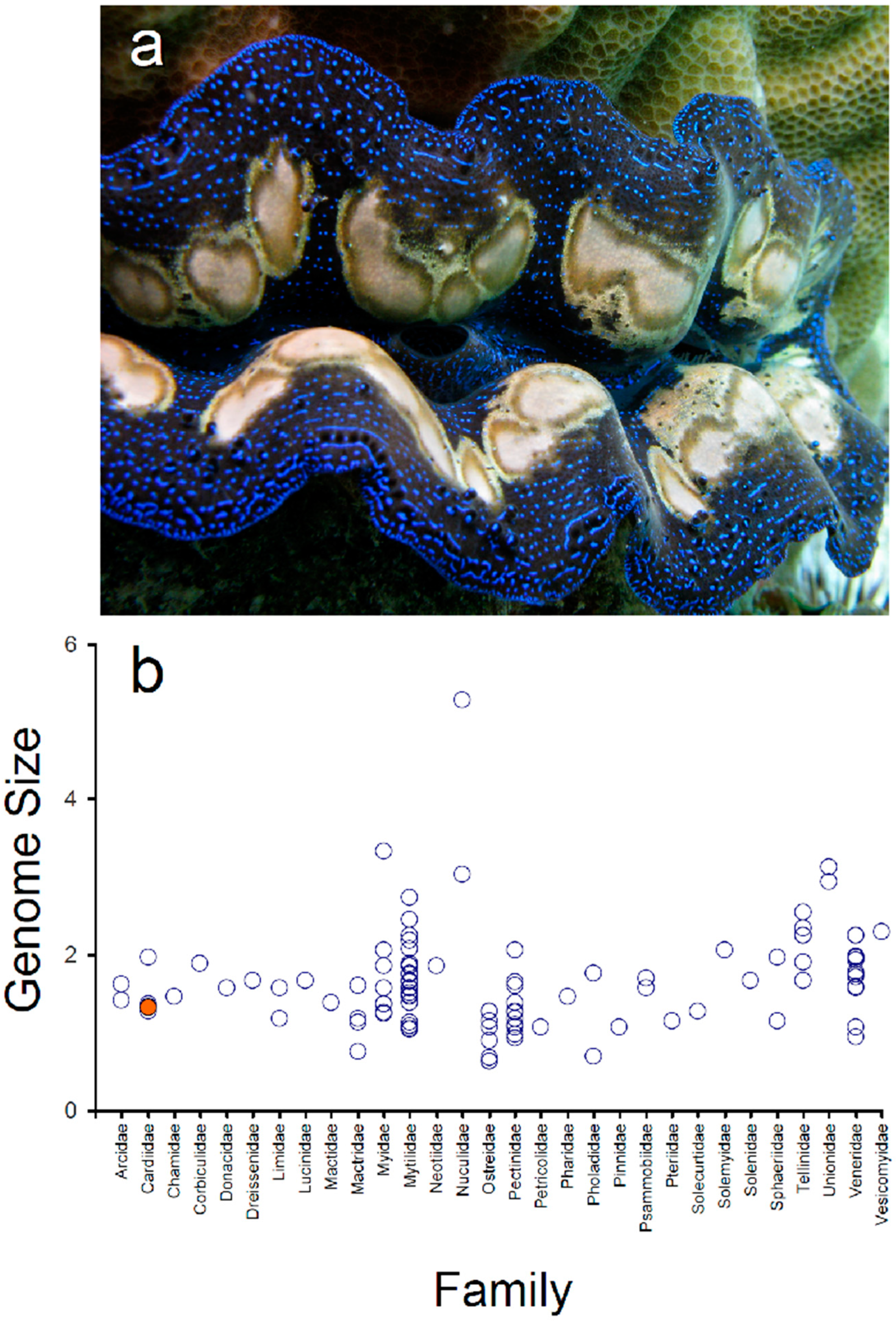
3.1. Genome Size Estimation in Tridacna crocea
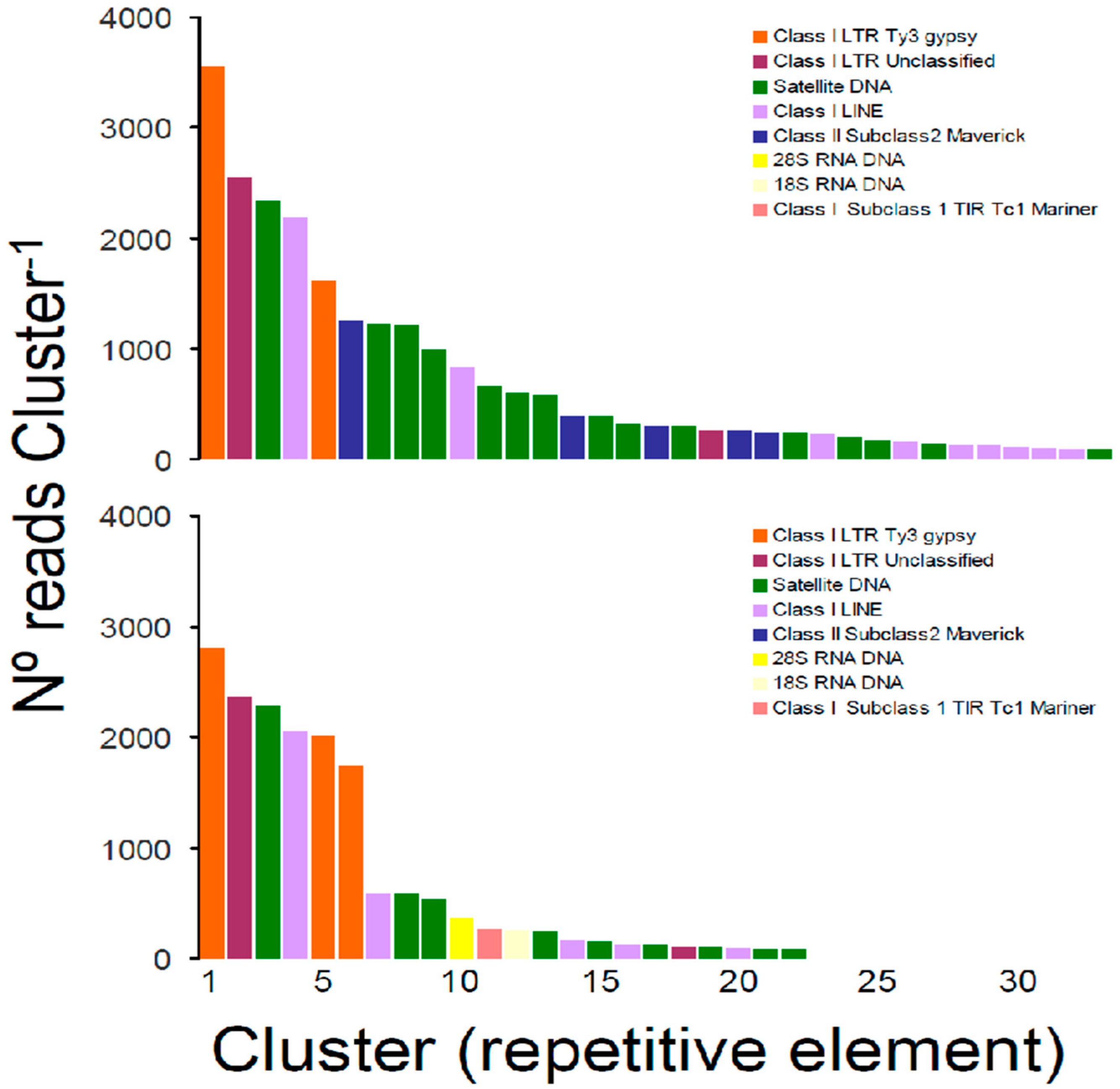
3.2. Repetitive Elements in the Nuclear Genome of Tridacna crocea
3.3. Nuclear Ribosomal Operon in Tridacna crocea
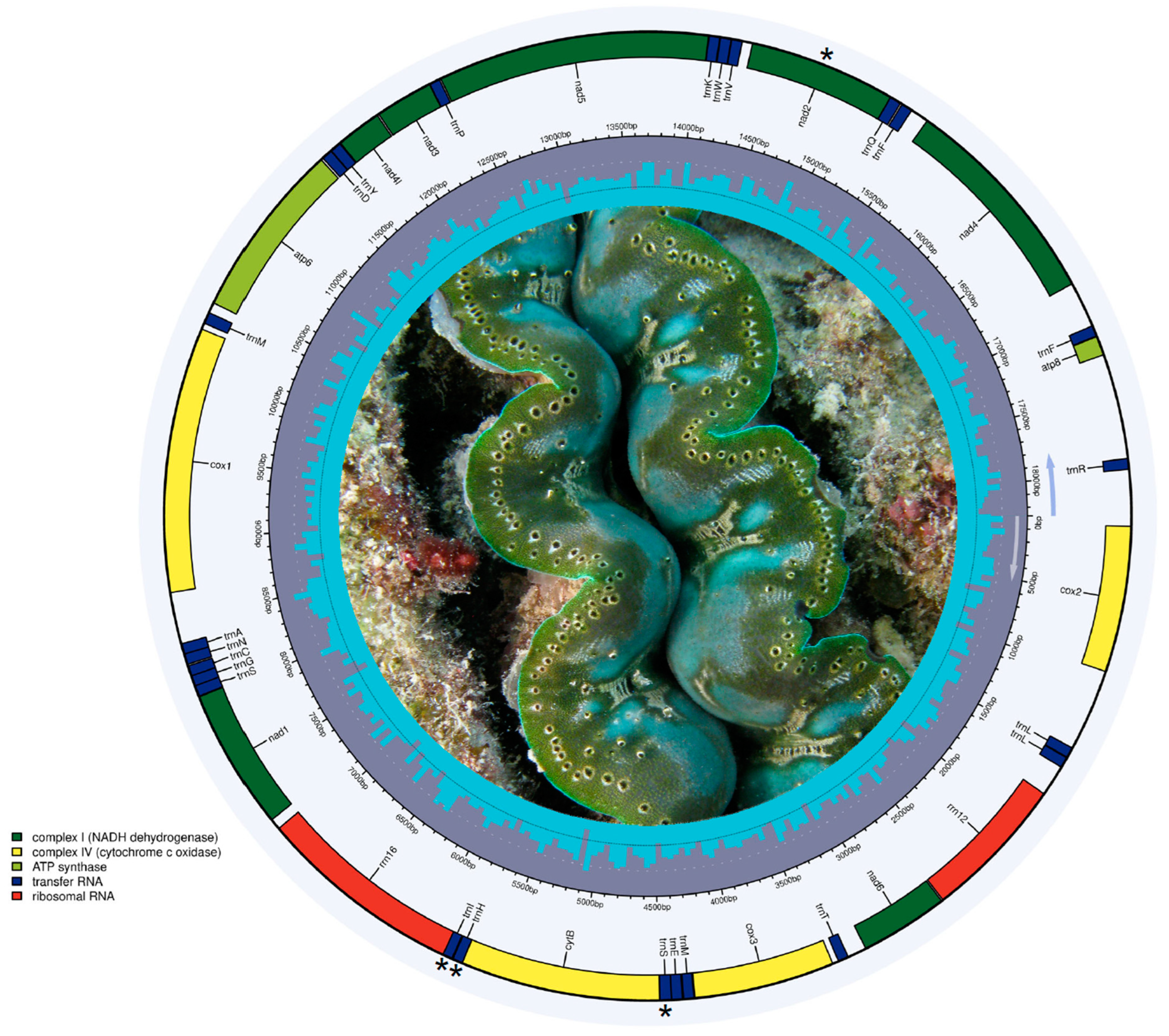
3.4. Mitochondrial Genome Assembly from RNA-seq in Tridacna crocea
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponder, W.F.; Lindberg, D.R. (Eds.) Phylogeny and Evolution of the Mollusca; University of California Press: Berkeley, CA, USA, 2008; p. 488. [Google Scholar]
- Yonge, C.M. Mode of life, feeding, digestion and symbiosis with zooxanthellae in the Tridacnidae. Sci. Rep. Gr. Barrier Reef Exped. 1936, 1, 283–321. [Google Scholar]
- Hawkins, A.J.S.; Klumpp, D.W. Nutrition of the giant clam Tridacna gigas L. II. Relative contributions of filter-feeding and the ammonium-nitrogen acquired and recycled by symbiotic alga towards total nitrogen requirements for tissue growth and metabolism. J. Exp. Mar. Biol. Ecol. 1995, 190, 263–290. [Google Scholar] [CrossRef]
- Norton, J.H.; Shepherd, M.A.; Long, H.M.; Fitt, W.K. The zooxanthellal tubular system in the giant clam. Biol. Bull. 1992, 183, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Holt, A.L.; Vahidinia, S.; Gagnon, Y.L.; Morse, D.E.; Sweeney, A.M. Photosymbiotic giant clams are transformers of solar flux. J. R. Soc. Interface 2014, 11, 20140678. [Google Scholar] [CrossRef] [Green Version]
- Rossbach, S.; Subedi, R.C.; Ng, T.K.; Ooi, B.S.; Duarte, C.M. Iridocytes mediate photonic cooperation between giant clams (Tridacninae) and their photosynthetic symbionts. Front. Mar. Sci. 2020, 7, 465. [Google Scholar] [CrossRef]
- Neo, M.L. Conservation of giant clams (Bivalvia: Cardiidae). In Encyclopedia of the World’s Biomes; Goldstein, M.I., DellaSala, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 4, pp. 527–538. [Google Scholar]
- Neo, M.L.; Wabnitz, C.C.C.; Braley, R.D.; Heslinga, G.A.; Fauvelot, C.; van Wynsberge, S.; Andréfouët, S.; Waters, C.; Tan, A.S.-H.; Gomez, E.D.; et al. Chapter 4. Giant clams (Bivalvia: Cardiidae: Tridacninae): A comprehensive update of species and their distribution, current threats and conservation status. In Oceanography and Marine Biology: An Annual Review; Hawkins, S.J., Evans, A.J., Dale, A.C., Firth, L.B., Hughes, D.J., Smith, I.P., Eds.; CRC Press: Boca Raton, FL, USA, 2017; Volume 55, pp. 87–388. [Google Scholar]
- Elfwing, T.; Plantman, P.; Tedengren, M.; Wijnbladh, E. Responses to temperature, heavy metal and sediment stress by the giant clam Tridacna squamosa. Mar. Freshw. Behav. Physiol. 2001, 34, 239–248. [Google Scholar] [CrossRef]
- Watson, S.-A.; Neo, M.L. Conserving threatened species during rapid environmental change: Using biological responses to inform management strategies of giant clams. Conserv. Physiol. 2021, 9, coab082. [Google Scholar] [CrossRef]
- Davila, F.; Sloan, T.; Milne, M.; van Kerkhoff, L. Impact assessment of giant clam research in the Indo-Pacific region. In ACIAR Impact Assessment Series Report No. 94; Australian Centre for International Agricultural Research: Canberra, Australia, 2017; p. 79. [Google Scholar]
- Moorhead, A. Giant clam aquaculture in the Pacific region: Perceptions of value and impact. Dev. Pract. 2017, 28, 624–635. [Google Scholar] [CrossRef] [Green Version]
- Mies, M.; Dor, P.; Guth, A.Z.; Sumida, P.Y.G. Production in giant clam aquaculture: Trends and challenges. Rev. Fish Sci. Aquac. 2017, 25, 286–296. [Google Scholar] [CrossRef]
- Rehm, L.; Marino, L.L.; Jonathan, R.; Holt, A.L.; McCourt, R.M.; Sweeney, A.M. Population structure of giant clams (subfamily: Tridacninae) across Palau: Implications for conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 617–632. [Google Scholar] [CrossRef]
- Hill, R.W.; Armstrong, E.J.; Inaba, K.; Morita, M.; Tresguerres, M.; Stillman, J.H.; Roa, J.N.; Kwan, G.T. Acid secretion by the boring organ of the burrowing giant clam, Tridacna crocea. Biol. Lett. 2018, 14, 20180047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamner, W.M.; Jones, M.S. Distribution, burrowing, and growth rates of the clam Tridacna crocea on interior reef flats. Oecologia 1976, 24, 207–227. [Google Scholar] [CrossRef] [PubMed]
- Neo, M.L.; Eckman, W.; Vicentuan, K.; Teo, S.L.-M.; Todd, P.A. The ecological significance of giant clams in coral reef ecosystems. Biol. Conserv. 2015, 181, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.S. Giant clams: Description, distribution and life history. In Giant Clams in Asia and the Pacific; Copland, J.W., Lucas, J.S., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1988; pp. 21–32. [Google Scholar]
- UNEP-WCMC (United Nations Environment Programme-World Conservation Monitoring Center). UNEP-WCMC Species Database: CITES-Listed Species; UNEP: Cambridge, UK, 2007. [Google Scholar]
- DeBoer, T.S.; Subia, M.D.; Ambariyanto Erdmann, M.V.; Kovitvongsa, K.; Barber, P.H. Phylogeography and limited genetic connectivity in the endangered boring giant clam across the Coral Triangle. Conserv. Biol. 2008, 22, 1255–1266. [Google Scholar] [CrossRef]
- Kochzius, M.; Nuryanto, A. Strong genetic population structure in the boring giant clam, Tridacna crocea, across the Indo-Malay Archipelago: Implications related to evolutionary processes and connectivity. Mol. Ecol. 2008, 17, 3775–3787. [Google Scholar] [CrossRef]
- Li, J.; Lemer, S.; Kirkendale, L.; Bieler, R.; Cavanaugh, C.; Giribet, G. Shedding light: A phylotranscriptomic perspective illuminates the origin of photosymbiosis in marine bivalves. BMC Evol. Biol. 2020, 20, 50. [Google Scholar] [CrossRef]
- Tan, E.Y.; Quek, Z.R.; Neo, M.L.; Fauvelot, C.; Huang, D. Genome skimming resolves the giant clam (Bivalvia: Cardiidae: Tridacninae) tree of life. Coral Reefs 2021, 40, 1–14. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Kokot, M.; Długosz, M.; Deorowicz, S. KMC 3: Counting and manipulating k-mer statistics. Bioinformatics 2017, 33, 2759–2761. [Google Scholar] [CrossRef] [Green Version]
- Sarmashghi, S.; Balaban, M.; Rachtman, E.; Touri, B.; Mirarab, S.; Bafna, V. Estimating repeat spectra and genome length from low-coverage genome skims with RESPECT. PLoS Comput. Biol. 2021, 17, e1009449. [Google Scholar] [CrossRef] [PubMed]
- Baeza, J.A. A first genomic portrait of the Florida stone crab Menippe mercenaria: Genome size, mitochondrial chromosome, and repetitive elements. Mar. Genom. 2021, 57, 100821. [Google Scholar] [CrossRef] [PubMed]
- Baeza, J.A.; Molina-Quirós, J.L.; Hernández-Muñoz, S. Genome survey sequencing of an iconic ‘Trophy’ sportfish, the Roosterfish Nematistius pectoralis: Genome size, repetitive elements, nuclear RNA gene operon, and microsatellite discovery. Genes 2021, 12, 1710. [Google Scholar] [CrossRef] [PubMed]
- Novak, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [Green Version]
- Novak, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Pevzner, P.A. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Baeza, J.A.; González, M.T. A first look at the ‘repeatome’of Benedenia humboldti, a major pathogen in yellowtail aquaculture: Repetitive element characterization, nuclear rRNA operon assembly, and microsatellite discovery. Mar. Genom. 2021, 58, 100848. [Google Scholar] [CrossRef]
- Hubley, R.; Finn, R.D.; Clements, J.; Eddy, S.R.; Jones, T.A.; Bao, W.; Smit, A.F.; Wheeler, T.J. The Dfam database of repetitive DNA families. Nucleic Acids Res. 2016, 44, D81–D89. [Google Scholar] [CrossRef] [Green Version]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Computac. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Han, L.; Liang, Z.; Hou, X. Comparative analysis of the ribosomal DNA repeat unit (rDNA) of Perna viridis (Linnaeus, 1758) and Perna canaliculus (Gmelin, 1791). PeerJ 2019, 7, e7644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sanchez-Garcia, M.; Ebersberger, I.; De Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914e919. [Google Scholar] [CrossRef]
- Nachtigall, P.G.; Grazziotin, F.G.; Junqueira-de-Azevedo, I.L. MITGARD: An automated pipeline for mitochondrial genome assembly in eukaryotic species using RNA-seq data. Brief Bioinform. 2021, 22, bbaa429. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Bushmanova, E.; Antipov, D.; Lapidus, A.; Prjibelski, A.D. rnaSPAdes: A de novo transcriptome assembler and its application to RNA-Seq data. GigaScience 2019, 8, giz100. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Zheng, S.; Poczai, P.; Hyvönen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An online program for the versatile plotting of organelle genomes. Front. Genet. 2020, 11, 576124. [Google Scholar] [CrossRef]
- Gregory, T.R. Animal Genome Size Database. 2021. Available online: http://www.genomesize.com (accessed on 15 January 2022).
- Hinegardner, R. Cellular DNA content of the Mollusca. Comp. Biochem. Physiol. 1974, 47, 447–460. [Google Scholar] [CrossRef]
- Rodríguez-Juíz, A.M.; Torrado, M.; Méndez, J. Genome-size variation in bivalve molluscs determined by flow cytometry. Mar. Biol. 1996, 126, 489–497. [Google Scholar] [CrossRef]
- Ip, J.C.H.; Xu, T.; Sun, J.; Li, R.; Chen, C.; Lan, Y.; Han, Z.; Zhang, H.; Wei, J.; Wang, H.; et al. Host–endosymbiont genome integration in a deep-sea chemosymbiotic clam. Mol. Biol. Evol. 2021, 38, 502–518. [Google Scholar] [CrossRef] [PubMed]
- Kenny, N.J.; McCarthy, S.A.; Dudchenko, O.; James, K.; Betteridge, E.; Corton, C.; Dolucan, J.; Mead, D.; Oliver, K.; Omer, A.D.; et al. The gene-rich genome of the scallop Pecten maximus. GigaScience 2020, 9, giaa037. [Google Scholar] [CrossRef]
- Steiner, G.; Hammer, S. Molecular phylogeny of the Bivalvia inferred from 18S rDNA sequences with particular reference to the Pteriomorphia. In The Evolutionary Biology of the Bivalvia; Harper, E.M., Taylor, J.D., Crame, J.A., Eds.; The Geological Society of London: London, UK, 2000; pp. 11–29. [Google Scholar]
- Passamaneck, Y.J.; Schander, C.; Halanych, K.M. Investigation of molluscan phylogeny using large-subunit and small-subunit nuclear rRNA sequences. Mol. Phylogenet. Evol. 2004, 32, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Faircloth, B.C.; Branstetter, M.G.; White, N.D.; Brady, S.G. Target enrichment of ultraconserved elements from arthropods provides a genomic perspective on relationships among Hymenoptera. Mol. Ecol. Res. 2015, 15, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, A.R.; Emme, S.A.; Lemmon, E.M. Anchored hybrid enrichment for massively high-throughput phylogenomics. Syst. Biol. 2012, 61, 727–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, M.; Kraemer, W.E.; Seidel, C.; Nuryanto, A.; Joshi, A.; Kochzius, M. Comparative genetic population structure of three endangered giant clams (Cardiidae: Tridacna species) throughout the Indo-West Pacific: Implications for divergence, connectivity and conservation. J. Molluscan Stud. 2016, 82, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Hui, M.; Nuryanto, A.; Kochzius, M. Concordance of microsatellite and mitochondrial DNA markers in detecting genetic population structure in the boring giant clam Tridacna crocea across the Indo-Malay Archipelago. Mar. Ecol. 2017, 38, e12389. [Google Scholar] [CrossRef] [Green Version]
- Huelsken, T.; Keyse, J.; Liggins, L.; Penny, S.; Treml, E.A.; Riginos, C. A novel widespread cryptic species and phylogeographic patterns within several giant clam species (Cardiidae: Tridacna) from the Indo-Pacific Ocean. PLoS ONE 2013, 8, e80858. [Google Scholar] [CrossRef] [Green Version]
- Keyse, J.; Treml, E.A.; Huelsken, T.; Barber, P.H.; DeBoer, T.; Kochzius, M.; Nuryanto, A.; Gardner, J.P.A.; Liu, L.-L.; Penny, S.; et al. Historical divergences associated with intermittent land bridges overshadow isolation by larval dispersal in co-distributed species of Tridacna giant clams. J. Biogeo. 2018, 45, 848–858. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baeza, J.A.; Neo, M.L.; Huang, D. Genomic Survey and Resources for the Boring Giant Clam Tridacna crocea. Genes 2022, 13, 903. https://doi.org/10.3390/genes13050903
Baeza JA, Neo ML, Huang D. Genomic Survey and Resources for the Boring Giant Clam Tridacna crocea. Genes. 2022; 13(5):903. https://doi.org/10.3390/genes13050903
Chicago/Turabian StyleBaeza, Juan Antonio, Mei Lin Neo, and Danwei Huang. 2022. "Genomic Survey and Resources for the Boring Giant Clam Tridacna crocea" Genes 13, no. 5: 903. https://doi.org/10.3390/genes13050903
APA StyleBaeza, J. A., Neo, M. L., & Huang, D. (2022). Genomic Survey and Resources for the Boring Giant Clam Tridacna crocea. Genes, 13(5), 903. https://doi.org/10.3390/genes13050903