
Opioid Use Disorder and Alternative mRNA Splicing in Reward Circuitry

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Post-Mortem Brain Samples
2.2. Sample and Data Processing
2.3. Analyses
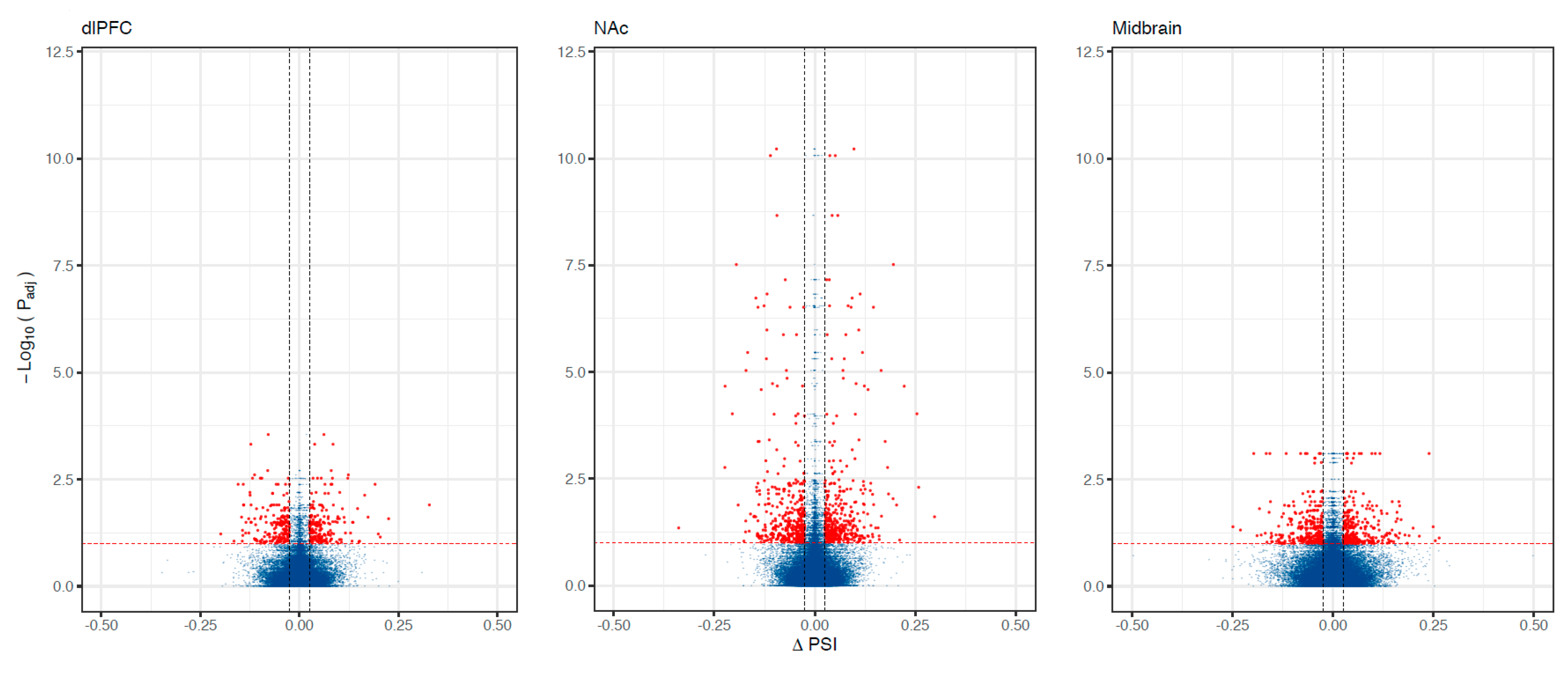
2.3.1. Differential Splicing
2.3.2. Differential Expression
2.3.3. Partitioned Heritability
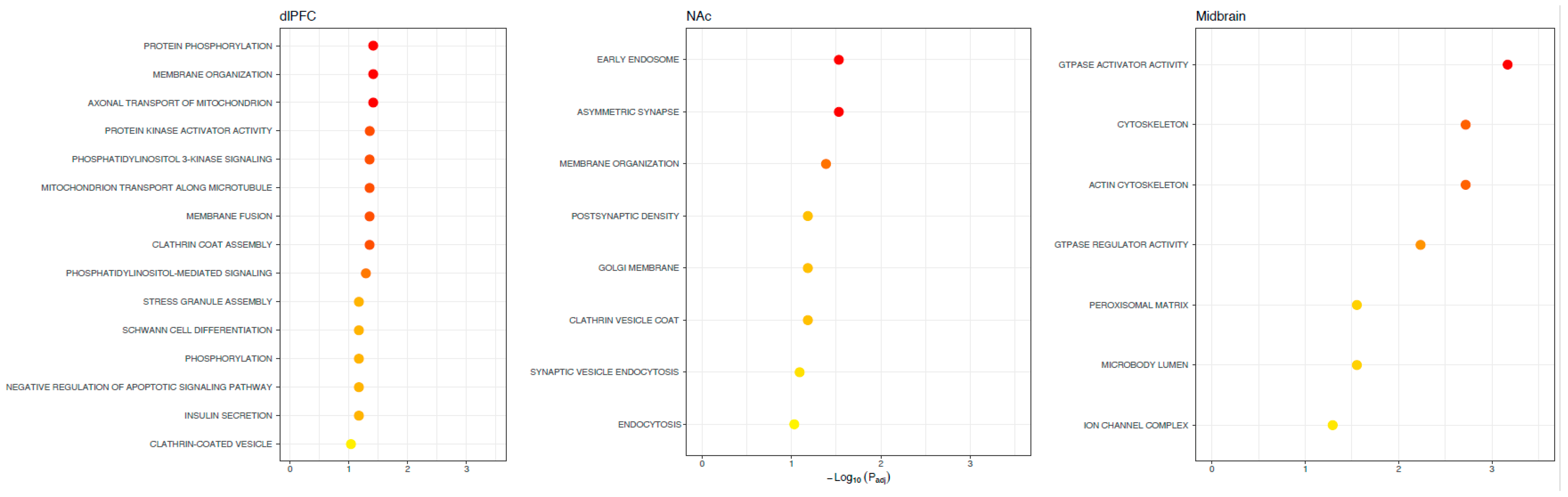
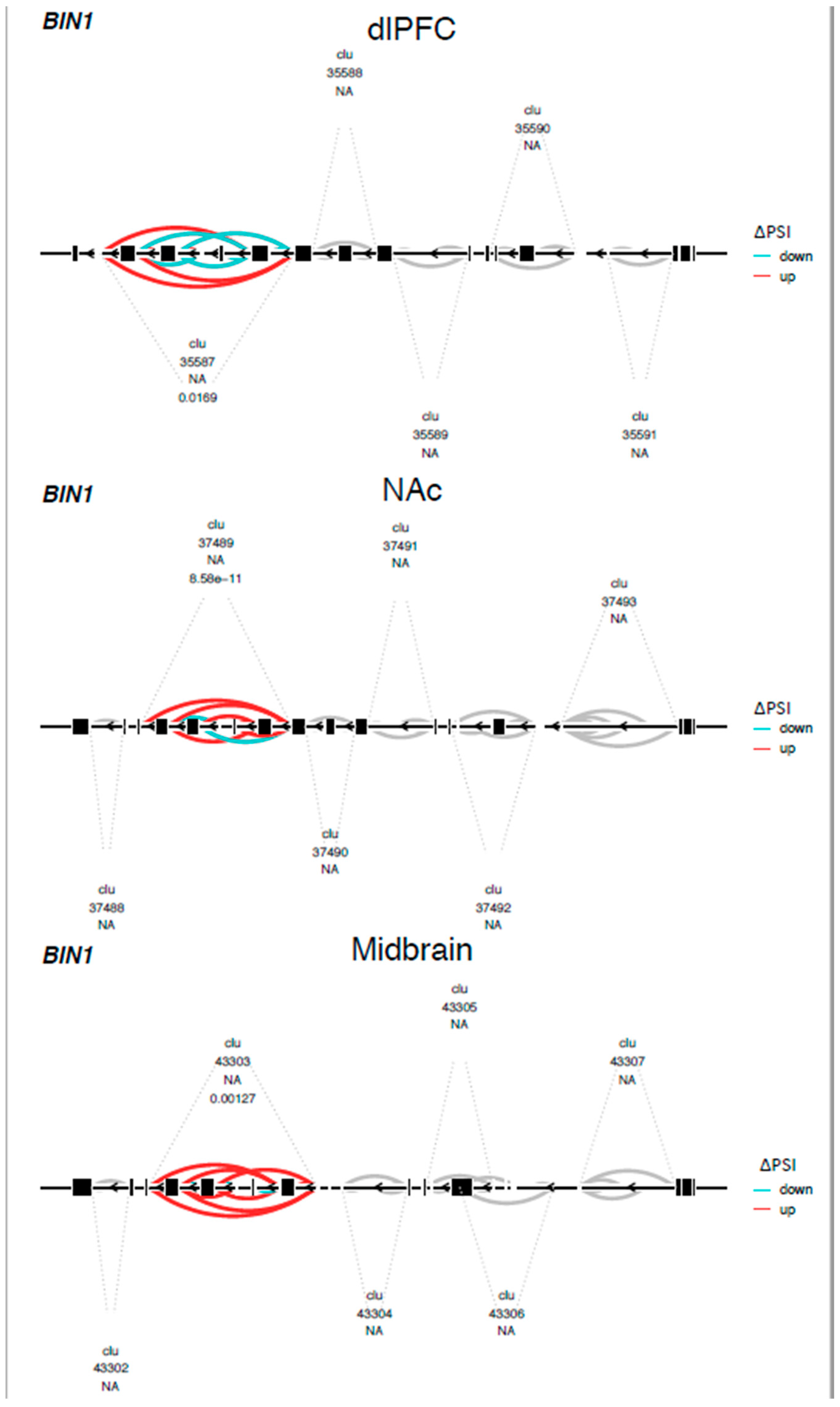
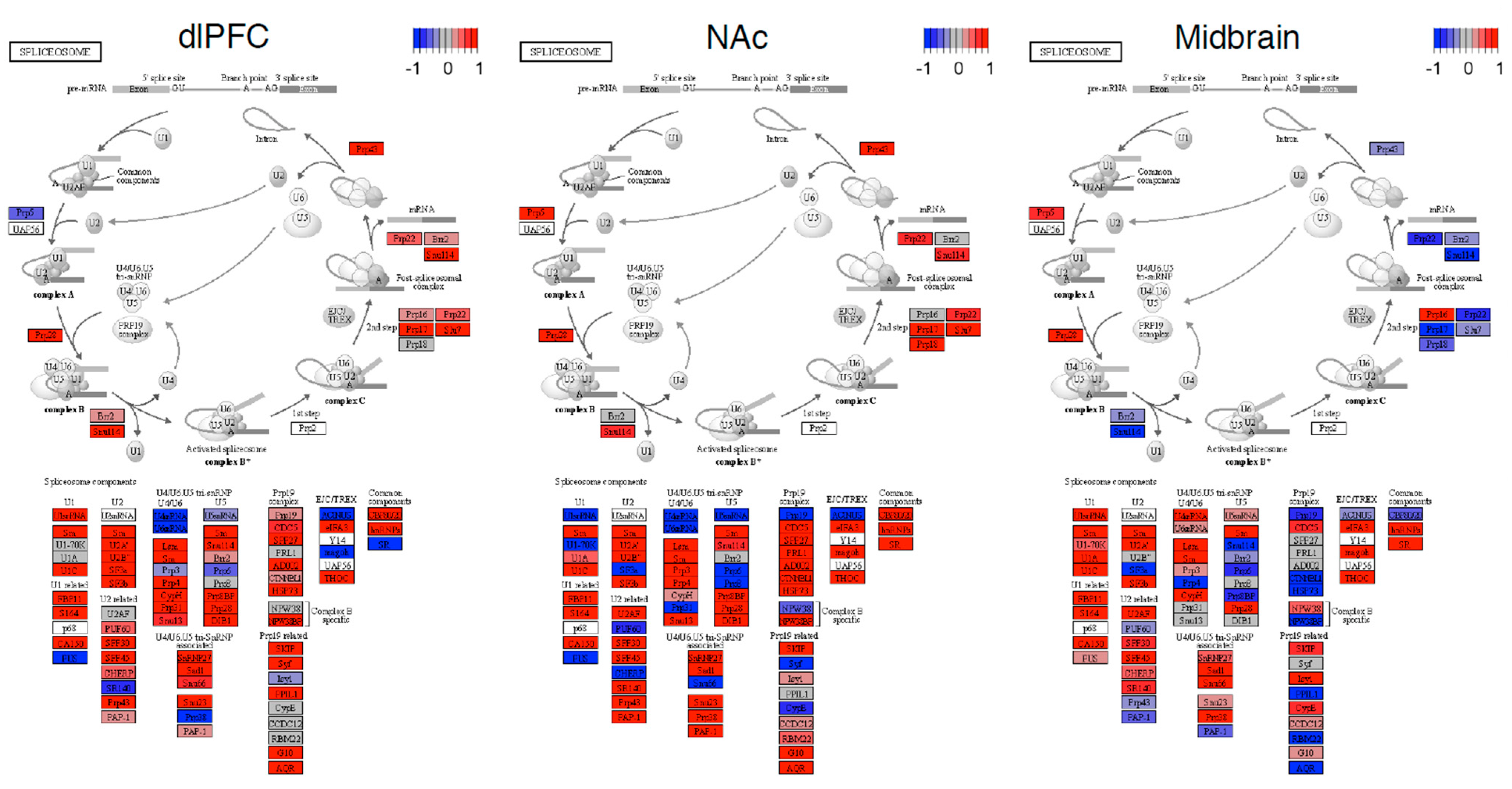
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mattson, C.L.; Tanz, L.J.; Quinn, K.; Kariisa, M.; Patel, P.; Davis, N.L. Trends and Geographic Patterns in Drug and Synthetic Opioid Overdose Deaths—United States, 2013–2019. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Seney, M.L.; Kim, S.M.; Glausier, J.R.; Hildebrand, M.A.; Xue, X.; Zong, W.; Wang, J.; Shelton, M.A.; Phan, B.N.; Srinivasan, C.; et al. Archival Report Transcriptional Alterations in Dorsolateral Prefrontal Cortex and Nucleus Accumbens Implicate Neuroinflammation and Synaptic Remodeling in Opioid Use Disorder. Biol. Psychiatry 2021, 90, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.H.; Rumschlag, M.; Guerra, M.H.; Savonen, C.L.; Jaster, A.M.; Olson, P.D.; Alazizi, A.; Luca, F.; Pique-Regi, R.; Schmidt, C.J.; et al. Differentially expressed gene networks, biomarkers, long noncoding RNAs, and shared responses with cocaine identified in the midbrains of human opioid abusers. Sci. Rep. 2019, 9, 1534. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Huang, B.O.; Xu, Y.M.; Li, J.; Huang, L.F.; Lin, J.; Zhang, J.; Min, Q.H.; Yang, W.M.; et al. Mechanism of alternative splicing and its regulation. Biomed. Rep. 2015, 3, 152–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iijima, T.; Hidaka, C.; Iijima, Y. Spatio-temporal regulations and functions of neuronal alternative RNA splicing in developing and adult brains. Neurosci. Res. 2018, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lipscombe, D.; Andrade, A.; Allen, S.E. Alternative splicing: Functional diversity among voltage-gated calcium channels and behavioral consequences. Biochim. Biophys. Acta 2014, 1828, 1522–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thalhammer, A.; Jaudon, F.; Cingolani, L.A. Emerging Roles of Activity-Dependent Alternative Splicing in Homeostatic Plasticity. Front. Cell. Neurosci. 2020, 14, 104. [Google Scholar] [CrossRef] [PubMed]
- Lopez Soto, E.J.; Gandal, M.J.; Gonatopoulos-Pournatzis, T.; Heller, E.A.; Luo, D.; Zheng, S. Mechanisms of Neuronal Alternative Splicing and Strategies for Therapeutic Interventions. J. Neurosci. 2019, 39, 8193–8199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razanau, A.; Xie, J. Emerging mechanisms and consequences of calcium regulation of alternative splicing in neurons and endocrine cells. Cell. Mol. Life Sci. 2013, 70, 4527–4536. [Google Scholar] [CrossRef] [PubMed]
- Gandal, M.J.; Zhang, P.; Hadjimichael, E.; Walker, R.L.; Chen, C.; Liu, S.; Won, H.; Van Bakel, H.; Varghese, M.; Wang, Y.; et al. Transcriptome-wide isoform-level dysregulation in ASD, schizophrenia. Science 2018, 362, eaat8127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasabe, T.; Ishiura, S. Alcoholism and Alternative Splicing of Candidate Genes. Int. J. Environ. Res. Public Health 2010, 7, 1448–1466. [Google Scholar] [CrossRef] [Green Version]
- Stolf, A.R.; Cupertino, R.B.; Müller, D.; Sanvicente-Vieira, B.; Roman, T.; Vitola, E.S.; Grevet, E.H.; von Diemen, L.; Kessler, F.H.P.; Grassi-Oliveira, R.; et al. Effects of DRD2 splicing-regulatory polymorphism and DRD4 48 bp VNTR on crack cocaine addiction. J. Neural Transm. 2019, 126, 193–199. [Google Scholar] [CrossRef]
- Moyer, R.A.; Wang, D.; Papp, A.C.; Smith, R.M.; Duque, L.; Mash, D.C.; Sadee, W. Intronic Polymorphisms Affecting Alternative Splicing of Human Dopamine D2 Receptor Are Associated with Cocaine Abuse. Neuropsychopharmacology 2010, 36, 753–762. [Google Scholar] [CrossRef]
- Sullivan, D.; Pinsonneault, J.K.; Papp, A.C.; Zhu, H.; Lemeshow, S.; Mash, D.C.; Sadee, W. Dopamine transporter DAT and receptor DRD2 variants affect risk of lethal cocaine abuse: A gene–gene–environment interaction. Transl. Psychiatry 2013, 3, e222. [Google Scholar] [CrossRef] [Green Version]
- Clarke, T.; Weiss, A.M.Y.R.D.; Ferarro, T.N.; Kampman, K.M. The dopamine receptor D2 (DRD2) SNP rs1076560 is associated with opioid addiction. Ann. Hum. Genet. 2014, 78, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, S.; Pinsonneault, J.K.; Sadee, W.; Lee, S.H.; Zafar, M.M.; Raja, M.S.; Raja, G.K. Significant Association of DRD2 Enhancer Variant rs12364283 with Heroin Addiction in a Pakastani Population. Ann. Hum. Genet. 2019, 83, 367–372. [Google Scholar] [CrossRef]
- Neve, K.A.; Ford, C.P.; Buck, D.C.; Grandy, D.K.; Neve, R.L.; Phillips, T.J. Normalizing Dopamine D2 Receptor-Mediated Responses in D2 Null Mutant Mice by Virus-Mediated Receptor Restoration: Comparing D2L and D2S. Neuroscience 2014, 17, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.G.; Xu, J.; Hurd, Y.L.; Pan, Y. Dysregulated expression of the alternatively spliced variant mRNAs of the mu opioid receptor gene, OPRM1, in the medial prefrontal cortex of male human heroin abusers and heroin self-administering male rats. J. Neurosci. Res. 2022, 100, 35–47. [Google Scholar] [CrossRef]
- Van Booven, D.; Li, M.; Sunil Rao, J.; Blokhin, I.O.; Dayne Mayfield, R.; Barbier, E.; Heilig, M.; Wahlestedt, C. Alcohol use disorder causes global changes in splicing in the human brain. Transl. Psychiatry 2021, 11, 2. [Google Scholar] [CrossRef]
- Kular, L.; Kular, S. Epigenetics applied to psychiatry: Clinical opportunities and future challenges. Psychiatry Clin. Neurosci. 2018, 72, 195–211. [Google Scholar] [CrossRef]
- Li, Y.I.; Wong, G.; Humphrey, J. Prioritizing Parkinson’s disease genes using population-scale transcriptomic data. Nat. Commun. 2019, 10, 994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huggett, S.B.; Ikeda, A.S.; Yuan, Q.; Benca-Bachman, C.E.; Palmer, R.H.C. Genome- and Transcriptome-wide Splicing Associations with Problematic Alcohol Use and Alcohol Use Disorder. bioRxiv 2021. [Google Scholar] [CrossRef]
- Huggett, S.B.; Stallings, M.C. Cocaine’omics: Genome-wide and transcriptome-wide analyses provide biological insight into cocaine use and dependence. Addict. Biol. 2019, 25, e12719. [Google Scholar] [CrossRef] [PubMed]
- Rossell, D.; Attolini, C.S.O.; Kroiss, M.; Stöcker, A. Quantifying alternative splicing from paired-end RNA-sequencing data. Ann. Appl. Stat. 2014, 8, 309–330. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Genome analysis Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner Alexander. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. Sequence analysis featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Abraham, G.; Qiu, Y.; Inouye, M. Genetics and population analysis FlashPCA2: Principal component analysis of Biobank-scale genotype datasets. Bioinformatics 2017, 33, 2776–2778. [Google Scholar] [CrossRef]
- Li, Y.I.; Knowles, D.A.; Humphrey, J.; Barbeira, A.N.; Dickinson, S.P.; Im, H.K.; Pritchard, J.K. Annotation-free quantification of RNA splicing using LeafCutter. Nat. Genet. 2018, 50, 151–158. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2021. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Rentsch, C.T.; Cheng, Z.; Kember, R.L.; Nunez, Y.Z.; Sherva, R.M.; Tate, J.P.; Dao, C.; Xu, K.; Polimanti, R.; et al. Association of OPRM1 Functional Coding Variant With Opioid Use Disorder A Genome-Wide Association Study. JAMA Psychiatry 2020, 1072, 1072–1081. [Google Scholar] [CrossRef]
- Gandal, M.J.; Leppa, V.; Won, H.; Parikshak, N.N.; Geschwind, D.H. The road to precision psychiatry: Translating genetics into disease mechanisms. Nat. Neurosci. 2016, 19, 1397–1407. [Google Scholar] [CrossRef] [Green Version]
- Bulik-Sullivan, B.K.; Loh, P.R.; Finucane, H.K.; Ripke, S.; Yang, J.; Patterson, N.; Yang, J.; Schizophrenia Working Group of the Psychiatric Genomics Consortium; Patterson, N.; Daly, M.J.; et al. LD Score Regression Distinguishes Confounding from Polygenicity in Genome-Wide Association Studies. Nat. Genet. 2015, 47, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Prokic, I.; Cowling, B.S.; Laporte, J. Amphiphysin 2 (BIN1) in physiology and diseases. J. Mol. Med. 2014, 92, 453–463. [Google Scholar] [CrossRef]
- Lazo, P.; Nadal, M.; Ferrer, M.; Area, E.; Hernández-Torres, J.; Nabokina, S.; Mollinedo, F.; Estivill, X. Genomic organization, chromosomal localization, alternative splicing, and isoforms of the human synaptosome-associated protein-23 gene implicated in vesicle-membrane fusion processes. Hum. Genet. 2001, 108, 211–215. [Google Scholar] [CrossRef]
- Liu, Y.; Surka, M.C.; Schroeter, T.; Lukiyanchuk, V.; Schmid, S.L. Isoform and Splice-Variant Specific Functions of Dynamin-2 Revealed by Analysis of Conditional Knock-Out Cells. Mol. Biol. Cell 2008, 19, 5347–5359. [Google Scholar] [CrossRef]
- Ellis, J.D.; Barrios-Rodiles, M.; Colak, R.; Irimia, M.; Kim, T.; Calarco, J.A.; Wang, X.; Pan, Q.; O’Hanlon, D.; Kim, P.M.; et al. Tissue-Specific Alternative Splicing Remodels Protein-Protein Interaction Networks. Cell 2012, 46, 884–892. [Google Scholar] [CrossRef] [Green Version]
- Koch, T.; Widera, A.; Bartzsch, K.; Schulz, S.; Brandenburg, L.O.; Wundrack, N.; Beyer, A.; Grecksch, G.; Höllt, V. Receptor Endocytosis Counteracts the Development of Opioid Tolerance Receptor Endocytosis Counteracts the Development of Opioid Tolerance. Mol. Pharmacol. 2005, 67, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Finn, A.K.; Whistler, J.L. Endocytosis of the Mu Opioid Receptor Reduces Tolerance and a Cellular Hallmark. Neuron 2001, 32, 829–839. [Google Scholar] [CrossRef] [Green Version]
- von Zastrow, M.; Svingos, A.; Haberstock-Debic, H.; Evans, C. Regulated endocytosis of opioid receptors: Cellular mechanisms and proposed roles in physiological adaptation to opiate drugs. Curr. Opin. Neurobiol. 2003, 13, 348–353. [Google Scholar] [CrossRef]
- Martini, L.; Whistler, L.J. The role of mu opioid receptor desensitization and endocytosis in morphine tolerance and dependence. Curr. Opin. Neurobiol. 2007, 17, 556–564. [Google Scholar] [CrossRef]
- Van der Feltz, C.; Hoskins, A.A. Structural and Functional Modularity of the U2 snRNP in pre- mRNA Splicing. Crit. Rev. Biochem. Mol. Biol. 2020, 54, 443–465. [Google Scholar] [CrossRef]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptive Information on the RNA-Seq Brain Data | ||||
|---|---|---|---|---|
| Variable | DlPFC and NAc [2] | Midbrain [3] | ||
| OUD | Control | OUD | Control | |
| Sex | 50% Female | 50% Female | 100% Female | 100% Female |
| Age: M (s.d.) | 46.9 (7.3) | 47.3 (9.5) | 49.5 (6.4) | 52.9 (2.0) |
| Race | 5% African-American | 35% African-American | 76.66% African-American | 60% African-American |
| RIN: M (s.d.) | 7.8 (0.7) | 8.0 (0.7) | 7.3 (0.5) | 7.4 (0.5) |
| Brain pH: M (s.d.) | 6.4 (0.2) | 6.6 (0.3) | 6.5 (0.2) | 6.6 (0.13) |
| Partitioned Heritability of OUD | |||
|---|---|---|---|
| Differential | Spliceosome | Differentially | |
| Expressed Genes | Genes | Spliced Genes | |
| Number of Genes | 922 | 127 | 1141 |
| Surrounding Region | 100 kb | 100 kb | 100 kb |
| % of Total SNPs | 6.06% | 0.89% | 5.71% |
| % of OUD h2SNP | 4.20% | 0.27% | 6.48% |
| Enrichment (se) h2SNP | 0.69 (0.47) | 0.32 (1.01) | 1.13 (0.37) |
| p-value h2SNP | 0.506 | 0.488 | 0.715 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huggett, S.B.; Ikeda, A.S.; McGeary, J.E.; Kaun, K.R.; Palmer, R.H.C. Opioid Use Disorder and Alternative mRNA Splicing in Reward Circuitry. Genes 2022, 13, 1045. https://doi.org/10.3390/genes13061045
Huggett SB, Ikeda AS, McGeary JE, Kaun KR, Palmer RHC. Opioid Use Disorder and Alternative mRNA Splicing in Reward Circuitry. Genes. 2022; 13(6):1045. https://doi.org/10.3390/genes13061045
Chicago/Turabian StyleHuggett, Spencer B., Ami S. Ikeda, John E. McGeary, Karla R. Kaun, and Rohan H. C. Palmer. 2022. "Opioid Use Disorder and Alternative mRNA Splicing in Reward Circuitry" Genes 13, no. 6: 1045. https://doi.org/10.3390/genes13061045
APA StyleHuggett, S. B., Ikeda, A. S., McGeary, J. E., Kaun, K. R., & Palmer, R. H. C. (2022). Opioid Use Disorder and Alternative mRNA Splicing in Reward Circuitry. Genes, 13(6), 1045. https://doi.org/10.3390/genes13061045