
Comparative Transcriptome and Interaction Protein Analysis Reveals the Mechanism of IbMPK3-Overexpressing Transgenic Sweet Potato Response to Low-Temperature Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cold Stress Treatment
2.2. RNA Extraction and cDNA Library Construction
2.3. Sequencing and Data Processing
2.4. Identification of Differentially-Expressed Genes (DEGs)
2.5. Functional Annotation and Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis
2.6. qRT-PCR Validation
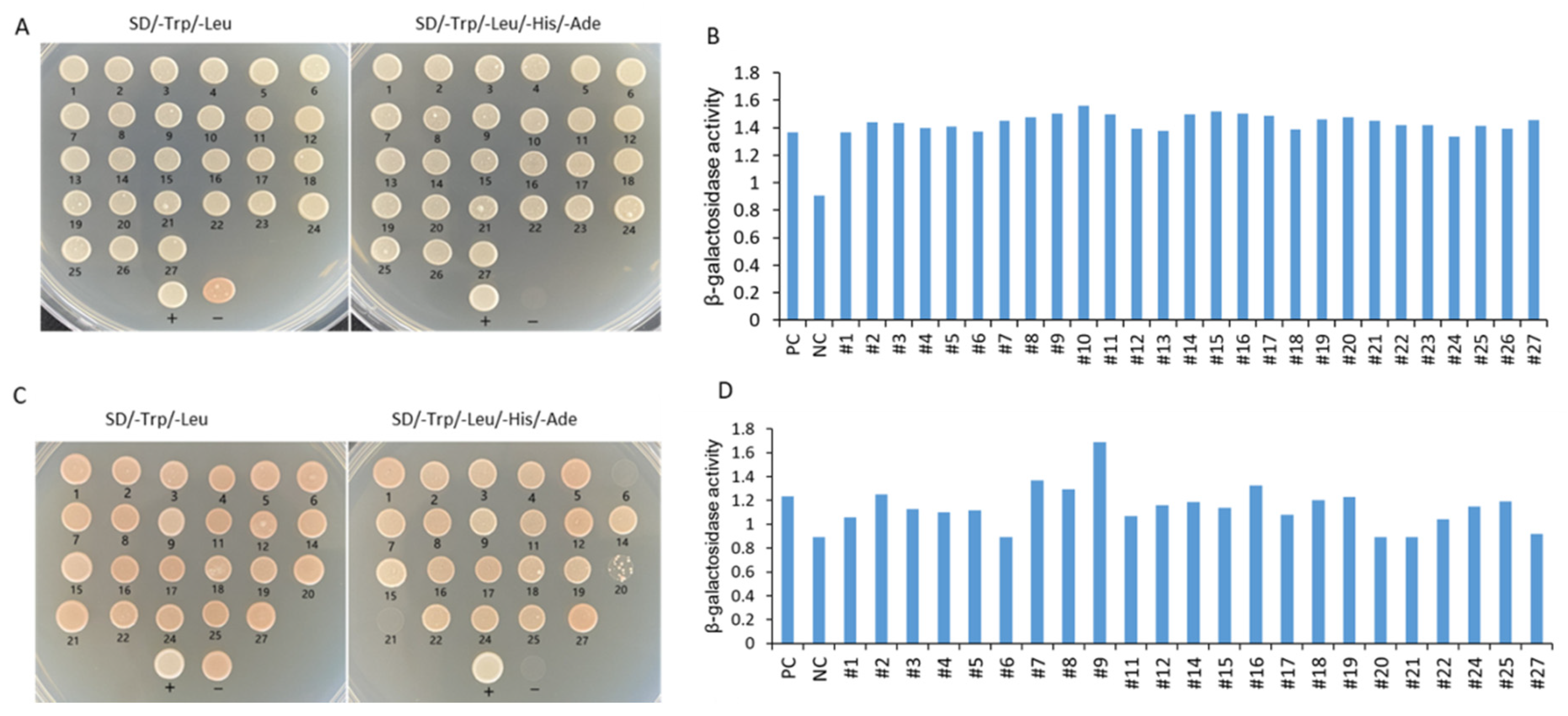
2.7. Yeast Two-Hybrid (Y2H) Analysis
3. Results
3.1. RNA-seq Data Analysis
3.2. RNA Sequencing Validation by qRT-PCR
3.3. Identification, Functional Annotation, and Cluster Analysis of DEGs
3.4. Functional Analysis of Common and Unique DEGs under Low-Temperature Stress
3.5. Analysis of MAPK Pathway in WT and M3 Plants during Short-Term Low-Temperature Stress
3.6. Analysis of Secondary Metabolic Pathway and ABA-Independent Pathway in WT and M3 Plants during Long-Term Low-Temperature Stress Conditions
3.7. IbMPK3 Interact Proteins Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Dong, W.J.O.M.; Longevity, C. ROS and ROS-mediated cellular signaling. Oxidative Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Hirt, H. Nuclear Signaling of Plant MAPKs. Front. Plant Sci. 2018, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Teige, M.; Scheikl, E.; Eulgem, T.; Dóczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Xie, G.; Kato, H.; Imai, R. Biochemical identification of the OsMKK6-OsMPK3 signalling pathway for chilling stress tolerance in rice. Biochem. J. 2012, 443, 95–102. [Google Scholar] [CrossRef]
- Vatanparast, M.; Park, Y. Comparative RNA-Seq Analyses of Solenopsis japonica (Hymenoptera: Formicidae) Reveal Gene in Response to Cold Stress. Genes 2021, 12, 1610. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR Signaling Cascade and Its Regulation in Plants Responding to Cold Stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, W.; Hu, Y.; Liu, X.; He, Y. Cis-regulatory element-based genome-wide identification of DREB1/CBF targets in Arabidopsis. Prog. Nat. Sci. Mater. Int. 2008, 18, 579–583. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, J.; Sun, T.; Wang, X.; Zhu, C.; Ai, Y.; Gu, H. The precise regulation of different COR genes by individual CBF transcription factors in Arabidopsis thaliana. J. Integr. Plant Biol. 2017, 59, 118–183. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.-H.; Hong, X.; Agarwal, M.; Zhu, J.-K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Coustry, F.; Maity, S.N.; de Crombrugghe, B. Studies on transcription activation by the multimeric CCAAT-binding factor CBF. J. Biol. Chem. 1995, 270, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, H.S.; Bahk, S.; An, J.; Yoo, Y.; Kim, J.-Y.; Chung, W.S. Phosphorylation of the transcriptional repressor MYB15 by mitogen-activated protein kinase 6 is required for freezing tolerance in Arabidopsis. Nucleic Acids Res. 2017, 45, 6613–6627. [Google Scholar] [CrossRef] [Green Version]
- Castroverde, C.D.M.; Dina, D. Temperature regulation of plant hormone signaling during stress and development. J. Exp. Bot. 2021, 72, 7436–7458. [Google Scholar] [CrossRef]
- Yu, J.; Cang, J.; Li, Y.; Huang, R.; Lu, Q.; Wang, X.; Liu, L.; Xu, Q.; Zhang, K. Salicylic acid-induced antioxidant protection against low temperature in cold-hardy winter wheat. Acta Physiol. Plant. 2016, 38, 261. [Google Scholar] [CrossRef]
- Sharma, M.; Laxmi, A. Jasmonates: Emerging Players in Controlling Temperature Stress Tolerance. Front. Plant Sci. 2016, 6, 1129. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, H.; Chen, Y.; Zhu, S.; Chen, M.; Lan, X.; Chen, G.; Liao, Z. Cold stress improves the production of artemisinin depending on the increase in endogenous jasmonate. Biotechnol. Appl. Biochem. 2016, 64, 305–314. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Kim, M.D.; Park, S.-C.; Yang, K.-S.; Jeong, J.C.; Lee, H.-S.; Kwak, S.-S. SCOF-1-expressing transgenic sweetpotato plants show enhanced tolerance to low-temperature stress. Plant Physiol. Biochem. 2011, 49, 1436–1441. [Google Scholar] [CrossRef]
- Ji, C.Y.; Jin, R.; Xu, Z.; Kim, H.S.; Lee, C.-J.; Kang, L.; Kim, S.-E.; Lee, H.-U.; Lee, J.S.; Kang, C.H.; et al. Overexpression of Arabidopsis P3B increases heat and low temperature stress tolerance in transgenic sweetpotato. BMC Plant Biol. 2017, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Jin, R.; Kim, H.S.; Yu, T.; Zhang, A.; Yang, Y.; Liu, M.; Yu, W.; Zhao, P.; Zhang, Q.; Cao, Q.; et al. Identification and function analysis of bHLH genes in response to cold stress in sweetpotato. Plant Physiol. Biochem. 2021, 169, 224–235. [Google Scholar] [CrossRef]
- Jin, R.; Kim, B.H.; Ji, C.Y.; Kim, H.S.; Li, H.M.; Ma, D.F.; Kwak, S.-S. Overexpressing IbCBF3 increases low temperature and drought stress tolerance in transgenic sweetpotato. Plant Physiol. Biochem. 2017, 118, 45–54. [Google Scholar] [CrossRef]
- Jin, R.; Kim, H.S.; Yu, T.; Liu, M.; Yu, W.; Zhao, P.; Zhang, A.; Zhang, Q.; Liu, Y.; Cao, Q.; et al. Overexpression of IbMPK3 increases low-temperature tolerance in transgenic sweetpotato. Plant Biotechnol. Rep. 2022, 16, 91–100. [Google Scholar] [CrossRef]
- Ji, C.Y.; Kim, H.S.; Lee, C.-J.; Kim, S.-E.; Lee, H.-U.; Nam, S.-S.; Li, Q.; Ma, D.-F.; Kwak, S.-S. Comparative transcriptome profiling of tuberous roots of two sweetpotato lines with contrasting low temperature tolerance during storage. Gene 2019, 727, 144244. [Google Scholar] [CrossRef]
- Ji, C.Y.; Chung, W.-H.; Kim, H.S.; Jung, W.Y.; Kang, L.; Jeong, J.C.; Kwak, S.-S. Transcriptome profiling of sweetpotato tuberous roots during low temperature storage. Plant Physiol. Biochem. 2016, 112, 97–108. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef]
- Tansey, W.; Koyejo, O.; Poldrack, R.A.; Scott, J.G. False discovery rate smoothing. J. Am. Stat. Assoc. 2018, 113, 1156–1171. [Google Scholar] [CrossRef] [Green Version]
- Soneral, P.; Skorseth, C. Gene Ontology Analysis of and Subtyping of Breast Tumors by RNA-Seq and BiNGO. FASEB J. 2018, 32, 117. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Hiraki, H.; Uemura, M.; Kawamura, Y. Calcium Signaling-LinkedCBF/DREB1Gene Expression was Induced Depending on the Temperature Fluctuation in the Field: Views from the Natural Condition of Cold Acclimation. Plant Cell Physiol. 2018, 60, 303–317. [Google Scholar] [CrossRef]
- Aslam, S.; Gul, N.; Mir, M.A.; Asgher, M.; Al-Sulami, N.; Abulfaraj, A.A.; Qari, S. Role of Jasmonate, Calcium, and Glutathione in plants to combat Abiotic Stresses through precise signaling cascade. Front. Plant Sci. 2021, 12, 668029. [Google Scholar] [CrossRef]
- Zhang, M.; Su, J.; Zhang, Y.; Xu, J.; Zhang, S. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 2018, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, C.; Wang, H.; Li, L.; Wang, C. The Function of MAPK Cascades in Response to Various Stresses in Horticultural Plants. Front. Plant Sci. 2020, 11, 952. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Nakashima, K.; Miyakawa, T.; Kuromori, T.; Tanokura, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular Basis of the Core Regulatory Network in ABA Responses: Sensing, Signaling and Transport. Plant Cell Physiol. 2010, 51, 1821–1839. [Google Scholar] [CrossRef] [PubMed]
- Tavanti, T.R.; Melo, A.A.R.d.; Moreira, L.D.K.; Sanchez, D.E.J.; Silva, R.d.S.; Silva, R.M.d.; Reis, A.R.d. Micronutrient fertilization enhances ROS scavenging system for alleviation of abiotic stresses in plants. Plant Physiol. Biochem. 2021, 160, 386–396. [Google Scholar] [CrossRef]
- Ogbe, A.A.; Finnie, J.F.; Van Staden, J. The role of endophytes in secondary metabolites accumulation in medicinal plants under abiotic stress. South Afr. J. Bot. 2020, 134, 126–134. [Google Scholar] [CrossRef]
- Mishra, B.; Chand, S.; Singh Sangwan, N. ROS management is mediated by ascorbate-glutathione-α-tocopherol triad in co-ordination with secondary metabolic pathway under cadmium stress in Withania somnifera. Plant Physiol. Biochem. 2019, 139, 620–629. [Google Scholar] [CrossRef]
- Bonza, M.C.; De Michelis, M.I. The plant Ca2+-ATPase repertoire: Biochemical features and physiological functions. Plant Biol. 2010, 13, 421–430. [Google Scholar] [CrossRef]
- Jian, L.-C.; Li, J.-H.; Chen, W.-P.; Li, P.H.; Ahlstrand, G.G. Cytochemical Localization of Calcium and Ca2+-ATPase Activity in Plant Cells under Chilling Stress: A Comparative Study between the Chilling-Sensitive Maize and the Chilling-Insensitive Winter Wheat. Plant Cell Physiol. 1999, 40, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Thiel, G.; Schmidt, T.; Rössler, O.G. Ca2+ Microdomains, Calcineurin and the Regulation of Gene Transcription. Cells 2021, 10, 875. [Google Scholar] [CrossRef]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.-P.; Thuleau, P.; Mazars, C. Calcium Sensors as Key Hubs in Plant Responses to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.A.; Fariduddin, Q.; Yusuf, M. Low-temperature stress: Is phytohormones application a remedy? Environ. Sci. Pollut. Res. 2017, 24, 21574–21590. [Google Scholar] [CrossRef]
- Ketehouli, T.; Nguyen Quoc, V.H.; Dong, J.; Do, H.; Li, X.; Wang, F. Overview of the roles of calcium sensors in plants’ response to osmotic stress signalling. Funct. Plant Biol. 2022, 49, 589–599. [Google Scholar] [CrossRef]
- Guo, X.; Liu, D.; Chong, K. Cold signaling in plants: Insights into mechanisms and regulation. J. Integr. Plant Biol. 2018, 60, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.M. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, D.; Pan, J.; Kong, X.; Liu, Y.; Sun, L.; Wang, L.; Li, D. Overexpression of a multiple stress-responsive gene, ZmMPK4, enhances tolerance to low temperature in transgenic tobacco. Plant Physiol. Biochem. 2012, 58, 174–181. [Google Scholar] [CrossRef]
- Taj, G.; Agarwal, P.; Grant, M.; Kumar, A. MAPK machinery in plants. Plant Signal. Behav. 2010, 5, 1370–1378. [Google Scholar] [CrossRef] [Green Version]
- Amorim, L.L.B.; da Fonseca Dos Santos, R.; Neto, J.P.B.; Guida-Santos, M.; Crovella, S.; Benko-Iseppon, A.M. Transcription Factors Involved in Plant Resistance to Pathogens. Curr. Protein Pept. Sci. 2016, 18, 335–351. [Google Scholar] [CrossRef]
- Poulaki, E.G.; Tsolakidou, M.-D.; Gkizi, D.; Pantelides, I.S.; Tjamos, S.E. The Ethylene Biosynthesis Genes ACS2 and ACS6 Modulate Disease Severity of Verticillium dahliae. Plants 2020, 9, 907. [Google Scholar] [CrossRef]
- Rubio, S.; Noriega, X.; Pérez, F.J. Abscisic acid (ABA) and low temperatures synergistically increase the expression of CBF/DREB1 transcription factors and cold-hardiness in grapevine dormant buds. Ann. Bot. 2018, 123, 681–689. [Google Scholar] [CrossRef]
- Duarte, K.E.; de Souza, W.R.; Santiago, T.R.; Sampaio, B.L.; Ribeiro, A.P.; Cotta, M.G.; da Cunha, B.A.D.B.; Marraccini, P.R.R.; Kobayashi, A.K.; Molinari, H.B.C. Identification and characterization of core abscisic acid (ABA) signaling components and their gene expression profile in response to abiotic stresses in Setaria viridis. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Li, H.; Zhang, X.; Xie, Q.; Gong, Z.; Yang, S. OST1 Kinase Modulates Freezing Tolerance by Enhancing ICE1 Stability in Arabidopsis. Dev. Cell 2015, 32, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Jia, Y.; Shi, Y.; Zhang, X.; Song, C.; Gong, Z.; Yang, S. OST1-mediated BTF3L phosphorylation positively regulates CBFs during plant cold responses. EMBO J. 2018, 37, e98228. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, B.; Zhai, J.; Wang, Q.; Song, S. Differential regulation of jasmonate responses in multiple jaz mutants. Plant Signal. Behav. 2021, 17, e1997240. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Wang, F.; Li, S.; Huang, X. Enhancing heterogenous expression and purification of bioactive Arabidopsis COI1 protein in tobacco. Braz. J. Bot. 2016, 39, 651–657. [Google Scholar] [CrossRef]
- Tian, H.; Qi, T.; Li, Y.; Wang, C.; Ren, C.; Song, S.; Huang, H. Regulation of the WD-repeat/bHLH/MYB complex by gibberellin and jasmonate. Plant Signal. Behav. 2016, 11, e1204060. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; An, F.; Feng, Y.; Li, P.; Xue, L.; A, M.; Jiang, Z.; Kim, J.-M.; To, T.K.; Li, W.; et al. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 12539–12544. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Tian, S.; Hou, L.; Huang, X.; Zhang, X.; Guo, H.; Yang, S. Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef] [Green Version]
- Mullins, E.D.; McCollum, T.G.; McDonald, R.E. Ethylene: A regulator of stress-induced ACC synthase activity in nonclimacteric fruit. Physiol. Plant. 1999, 107, 1–7. [Google Scholar] [CrossRef]
- Lavy, M.; Prigge, M.J.; Tao, S.; Shain, S.; Kuo, A.; Kirchsteiger, K.; Estelle, M. Constitutive auxin response inPhyscomitrellareveals complex interactions between Aux/IAA and ARF proteins. eLife 2016, 5, 427. [Google Scholar] [CrossRef]
- Shani, E.; Salehin, M.; Zhang, Y.; Sanchez, S.E.; Doherty, C.; Wang, R.; Mangado, C.C.; Song, L.; Tal, I.; Pisanty, O.; et al. Plant Stress Tolerance Requires Auxin-Sensitive Aux/IAA Transcriptional Repressors. Curr. Biol. 2017, 27, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.; Sugita, K.; Qin, Y.; Rahman, A. Aux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. Int. J. Mol. Sci. 2020, 21, 8441. [Google Scholar] [CrossRef]
- Zhou, M.Q.; Shen, C.; Wu, L.H.; Tang, K.X.; Lin, J. CBF-dependent signaling pathway: A key responder to low temperature stress in plants. Crit. Rev. Biotechnol. 2011, 31, 186–192. [Google Scholar] [CrossRef]
- Debbarma, J.; Sarki, Y.N.; Saikia, B.; Boruah, H.P.D.; Singha, D.L.; Chikkaputtaiah, C. Ethylene Response Factor (ERF) Family Proteins in Abiotic Stresses and CRISPR-Cas9 Genome Editing of ERFs for Multiple Abiotic Stress Tolerance in Crop Plants: A Review. Mol. Biotechnol. 2019, 61, 153–172. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, T.-J.; Liu, J.-M.; Liu, W.-Q.; Liu, Q.; Yan, Y.-B.; Zhou, H.-M. The conserved Ala37 in the ERF/AP2 domain is essential for binding with the DRE element and the GCC box. FEBS Lett. 2006, 580, 1303–1308. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Han, J.; Xu, T.; Li, X.; Yao, C.; Li, T.; Sun, X.; Wang, X.; Yang, G. Overexpression of MbERF12, an ERF gene from (Malus baccata L.) Borkh, increases cold and salt tolerance in Arabidopsis thaliana associated with ROS scavenging through ethylene signal transduction. In Vitro Cell. Dev. Biol. Plant 2021, 57, 760–770. [Google Scholar] [CrossRef]
- Stock, J.; Bräutigam, A.; Melzer, M.; Bienert, G.P.; Bunk, B.; Nagel, M.; Overmann, J.; Keller, E.R.J.; Mock, H.-P. The transcription factor WRKY22 is required during cryo-stress acclimation in Arabidopsis shoot tips. J. Exp. Bot. 2020, 71, 4993–5009. [Google Scholar] [CrossRef]
- Zhu, L.; Li, S.; Ouyang, M.; Yang, L.; Sun, S.; Wang, Y.; Cai, X.; Wu, G.; Li, Y. Overexpression of watermelon ClWRKY20 in transgenic Arabidopsis improves salt and low-temperature tolerance. Sci. Hortic. 2022, 295, 110848. [Google Scholar] [CrossRef]
- Guo, F.; Liu, S.; Zhang, C.; Dong, T.; Meng, X.; Zhu, M. Genome-wide systematic survey and analysis of NAC transcription factor family and their response to abiotic stress in sweetpotato. Sci. Hortic. 2022, 299, 111048. [Google Scholar] [CrossRef]
- Li, X.-D.; Zhuang, K.-Y.; Liu, Z.-M.; Yang, D.-Y.; Ma, N.-N.; Meng, Q.-W. Overexpression of a novel NAC-type tomato transcription factor, SlNAM1, enhances the chilling stress tolerance of transgenic tobacco. J. Plant Physiol. 2016, 204, 54–65. [Google Scholar] [CrossRef]
- Hou, X.-M.; Zhang, H.-F.; Liu, S.-Y.; Wang, X.-K.; Zhang, Y.-M.; Meng, Y.-C.; Luo, D.; Chen, R.-G. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- Arif, M.A.; Hiss, M.; Tomek, M.; Busch, H.; Meyberg, R.; Tintelnot, S.; Reski, R.; Rensing, S.A.; Frank, W. ABA-Induced Vegetative Diaspore Formation in Physcomitrella patens. Front. Plant Sci. 2019, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Manasa, S., L.; Panigrahy, M.; Panigrahi, K.C.S.; Rout, G.R. Overview of Cold Stress Regulation in Plants. Bot. Rev. 2021, 1–29. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, N.; Agarwal, P.; Khurana, P. Heat shock factors in rice (Oryza sativa L.): Genome-wide expression analysis during reproductive development and abiotic stress. Mol. Genet. Genom. MGG 2011, 286, 171–187. [Google Scholar] [CrossRef]
- Deng, X.; Wang, J.; Wang, J.; Tian, W. Two HbHsfA1 and HbHsfB1 genes from the tropical woody plant rubber tree confer cold stress tolerance in Saccharomyces cerevisiae. Braz. J. Bot. 2018, 41, 711–724. [Google Scholar] [CrossRef]
- Gong, C.; Pang, Q.; Li, Z.; Li, Z.; Chen, R.; Sun, G.; Sun, B. Genome-Wide Identification and Characterization of Hsf and Hsp Gene Families and Gene Expression Analysis under Heat Stress in Eggplant (Solanum melongema L.). Horticulturae 2021, 7, 149. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | NCBI Accession | NCBI Description |
|---|---|---|
| 1 | XM_031267007.1 | PREDICTED: Ipomoea triloba calmodulin-7 (LOC116025682), mRNA (84–633) |
| 2 | XM_031270274.1 | PREDICTED: Ipomoea triloba 40S ribosomal protein S16-like (LOC116028540), mRNA (193–663) |
| 3 | XM_031274641.1 | PREDICTED: Ipomoea triloba chlorophyll a-b binding protein of LHCII type 1-like (LOC116032192), mRNA (31–769) |
| 4 | XM_031244518.1 | PREDICTED: Ipomoea triloba photosystem I reaction center subunit IV B, chloroplastic (LOC116004457), mRNA (1–540) |
| 10 | PREDICTED: Ipomoea triloba photosystem I reaction center subunit IV B, chloroplastic (LOC116004457), mRNA (1–586) | |
| 5 | XM_031252462.1 | PREDICTED: Ipomoea triloba ribulose bisphosphate carboxylase small chain SSU11A, chloroplastic-like (LOC116012798), mRNA (274–786) |
| 7 | XM_031272632.1 | PREDICTED: Ipomoea triloba peptidyl-prolyl cis-trans isomerase-like (LOC116030396), mRNA (29–776) |
| 8 | XM_031276471.1 | PREDICTED: Ipomoea triloba 60S ribosomal protein L3 (LOC116033703), mRNA (652–1447) |
| 9 | XM_031272345.1 | PREDICTED: Ipomoea triloba (S)-ureidoglycine aminohydrolase (LOC116030182), mRNA (152–1218) |
| 11 | XM_031239861.1 | PREDICTED: Ipomoea triloba NAC domain-containing protein 2-like (LOC115999908), mRNA (61–803) |
| 26 | PREDICTED: Ipomoea triloba NAC domain-containing protein 2-like (LOC115999908), mRNA (61–1047) | |
| 12 | XM_031258537.1 | PREDICTED: Ipomoea triloba translationally-controlled tumor protein homolog (LOC116017878), mRNA (166–709) |
| 14 | XM_031241907.1 | PREDICTED: Ipomoea triloba mannose/glucose-specific lectin-like (LOC116001939), mRNA (363–672) |
| 15 | XM_019333621.1 | PREDICTED: Ipomoea nil RHOMBOID-like protein 2 (LOC109183524), mRNA ( |
| 16 | XM_031275192.1 | PREDICTED: Ipomoea triloba chlorophyll a-b binding protein CP24 10A, chloroplastic (LOC116032562), mRNA (62–803) |
| 17 | XM_031246527.1 | PREDICTED: Ipomoea triloba 40S ribosomal protein S14 (LOC116006230), mRNA (78–754) |
| 18 | XM_031246505.1 | PREDICTED: Ipomoea triloba protein METHYLENE BLUE SENSITIVITY 1-like (LOC116006203), mRNA (75–680) |
| 19 | XM_031272347.1 | PREDICTED: Ipomoea triloba aquaporin TIP1-1-like (LOC116030185), mRNA (54–1070) |
| 22 | XM_031272599.1 | PREDICTED: Ipomoea triloba chlorophyll a-b binding protein CP29.1, chloroplastic-like (LOC116030369), mRNA (617–1076) |
| 24 | XM_031257562.1 | PREDICTED: Ipomoea triloba glycine-rich protein-like (LOC116017048), mRNA (239–526) |
| 25 | MG001450.1 | I.batatas metallothionein mRNA, complete cds (2–563) |
| 27 | XM_031255147.1 | PREDICTED: Ipomoea triloba photosystem I reaction center subunit psaK, chloroplastic (LOC116015131), mRNA (90–597) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, R.; Yu, T.; Guo, P.; Liu, M.; Pan, J.; Zhao, P.; Zhang, Q.; Zhu, X.; Wang, J.; Zhang, A.; et al. Comparative Transcriptome and Interaction Protein Analysis Reveals the Mechanism of IbMPK3-Overexpressing Transgenic Sweet Potato Response to Low-Temperature Stress. Genes 2022, 13, 1247. https://doi.org/10.3390/genes13071247
Jin R, Yu T, Guo P, Liu M, Pan J, Zhao P, Zhang Q, Zhu X, Wang J, Zhang A, et al. Comparative Transcriptome and Interaction Protein Analysis Reveals the Mechanism of IbMPK3-Overexpressing Transgenic Sweet Potato Response to Low-Temperature Stress. Genes. 2022; 13(7):1247. https://doi.org/10.3390/genes13071247
Chicago/Turabian StyleJin, Rong, Tao Yu, Pengyu Guo, Ming Liu, Jiaquan Pan, Peng Zhao, Qiangqiang Zhang, Xiaoya Zhu, Jing Wang, Aijun Zhang, and et al. 2022. "Comparative Transcriptome and Interaction Protein Analysis Reveals the Mechanism of IbMPK3-Overexpressing Transgenic Sweet Potato Response to Low-Temperature Stress" Genes 13, no. 7: 1247. https://doi.org/10.3390/genes13071247
APA StyleJin, R., Yu, T., Guo, P., Liu, M., Pan, J., Zhao, P., Zhang, Q., Zhu, X., Wang, J., Zhang, A., Cao, Q., & Tang, Z. (2022). Comparative Transcriptome and Interaction Protein Analysis Reveals the Mechanism of IbMPK3-Overexpressing Transgenic Sweet Potato Response to Low-Temperature Stress. Genes, 13(7), 1247. https://doi.org/10.3390/genes13071247