
Genomic Structure, Protein Character, Phylogenic Implication, and Embryonic Expression Pattern of a Zebrafish New Member of Zinc Finger BED-Type Gene Family

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Zebrafish Husbandry and Microscopy
2.3. Sequence Alignments
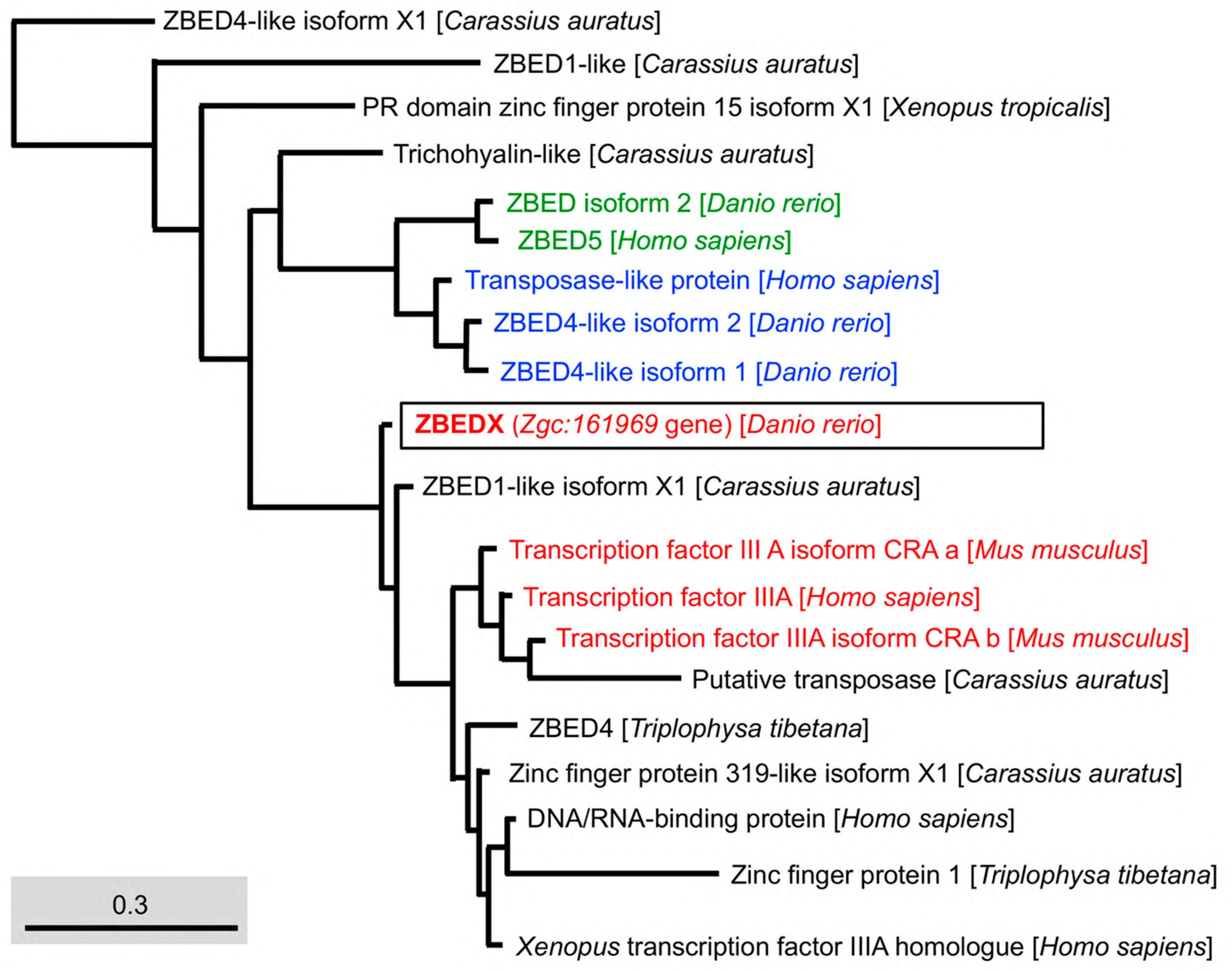
2.4. Phylogenetic Tree and Homology Analyses
- (1)
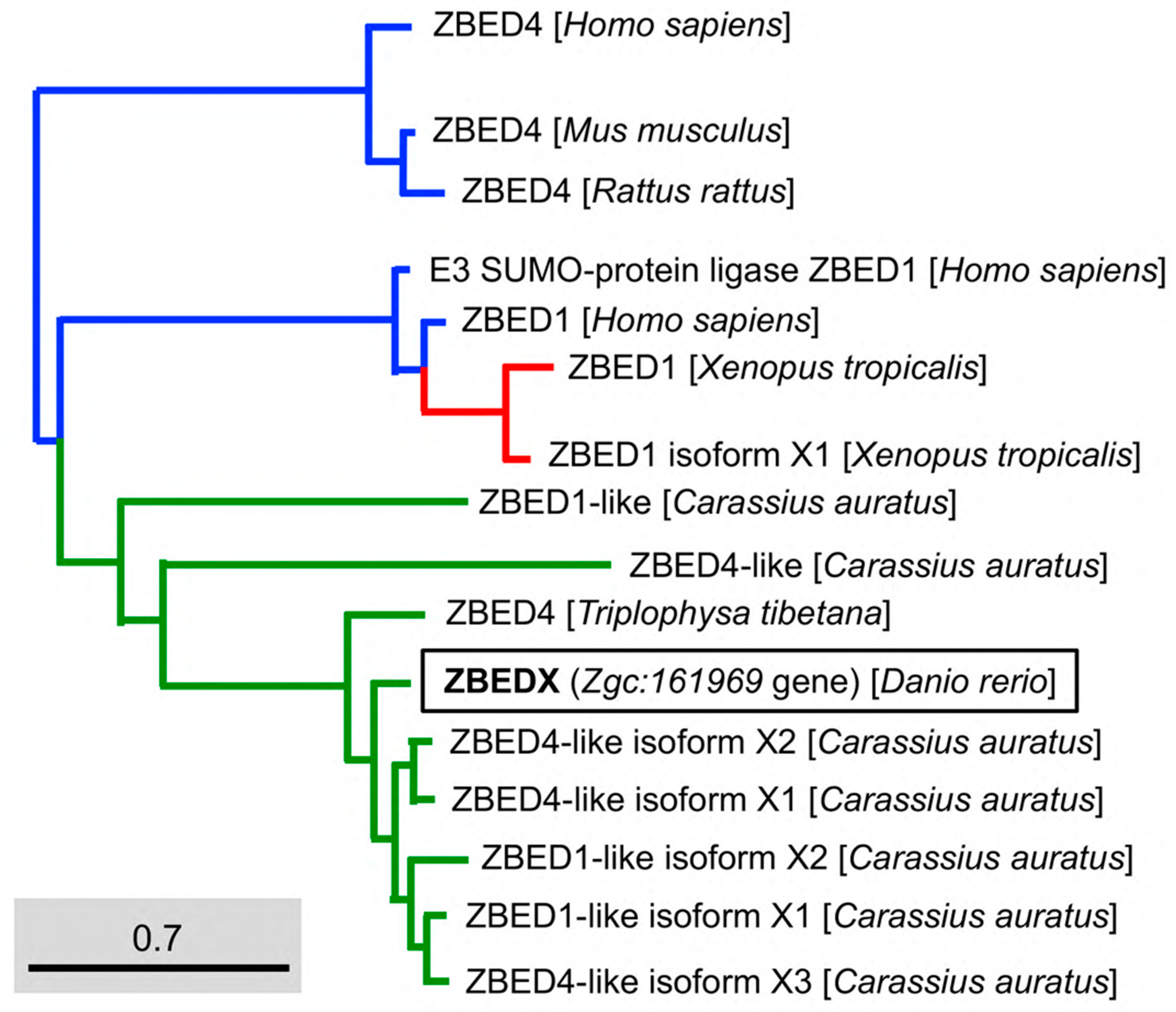
- Based on the full-length amino acid sequence: the alignment between LOC569044 and other members in the ZBED family of known species, including Homo sapiens ZBED4 (AAI67155.1), E3 SUMO-protein ligase ZBED1 (NP_001164606.1) and ZBED1 (AAH15030.1); Rattus norvegicus ZBED4 (XP_032775174.1); Mus musculus ZBED4 (NP_852077.1); Xenopus tropicalis ZBED1 (XP_004911791.1), and ZBED1 isoform X1 (XP_012813067.2); Carassius auratus ZBED1-like (XP_026072939.1), ZBED1-like isoform X1 (XP_026117831.1), ZBED1-like isoform X2 (XP_026117841.1), ZBED4-like (XP_026116228.1), ZBED4-like isoform X1 (XP_026069790.1), ZBED4-like isoform X2 (XP_026069791.1), ZBED4-like isoform X3 (XP_026117848.1), and Triplophysa tibetana ZBED4 (KAA0713175.1).
- (2)
- Based on the BED domain: the alignment between LOC569044 and other known species, including H. sapiens ZBED5 (NP_001137139.1), Transposase-like protein (AAF18454.1), Transcription factor IIIA (NP_002088.2), DNA/RNA-binding protein (AAA75623.1), and Xenopus transcription factor IIIA homologue (BAA06988.1); M. musculus Transcription factor IIIA isoform CRA a (EDL05819.1) and Transcription factor IIIA isoform CRA b (EDL05820.1); X. tropicalis PR domain zinc finger protein 15 isoform X1 (XP_012813515.1); C. auratus ZBED1-like (XP_026072939.1), ZBED4-like isoform X1 (XP_026117831.1), Trichohyalin-like (XP_026072553.1), ZBED1-like isoform X1 (XP_026117831.1), Zinc finger protein 319-like isoform X1 (XP_026084591.1) and Putative transposase (AFC96943.1); T. tibetana ZBED4 (KAA0712498.1), and Zinc finger protein 1 (KAA0710290.1).
2.5. Protein Structure Prediction
2.6. RNA Extraction, cDNA Synthesis, Polymerase Chain Reaction (PCR), and Antisense mRNA Probe Synthesis
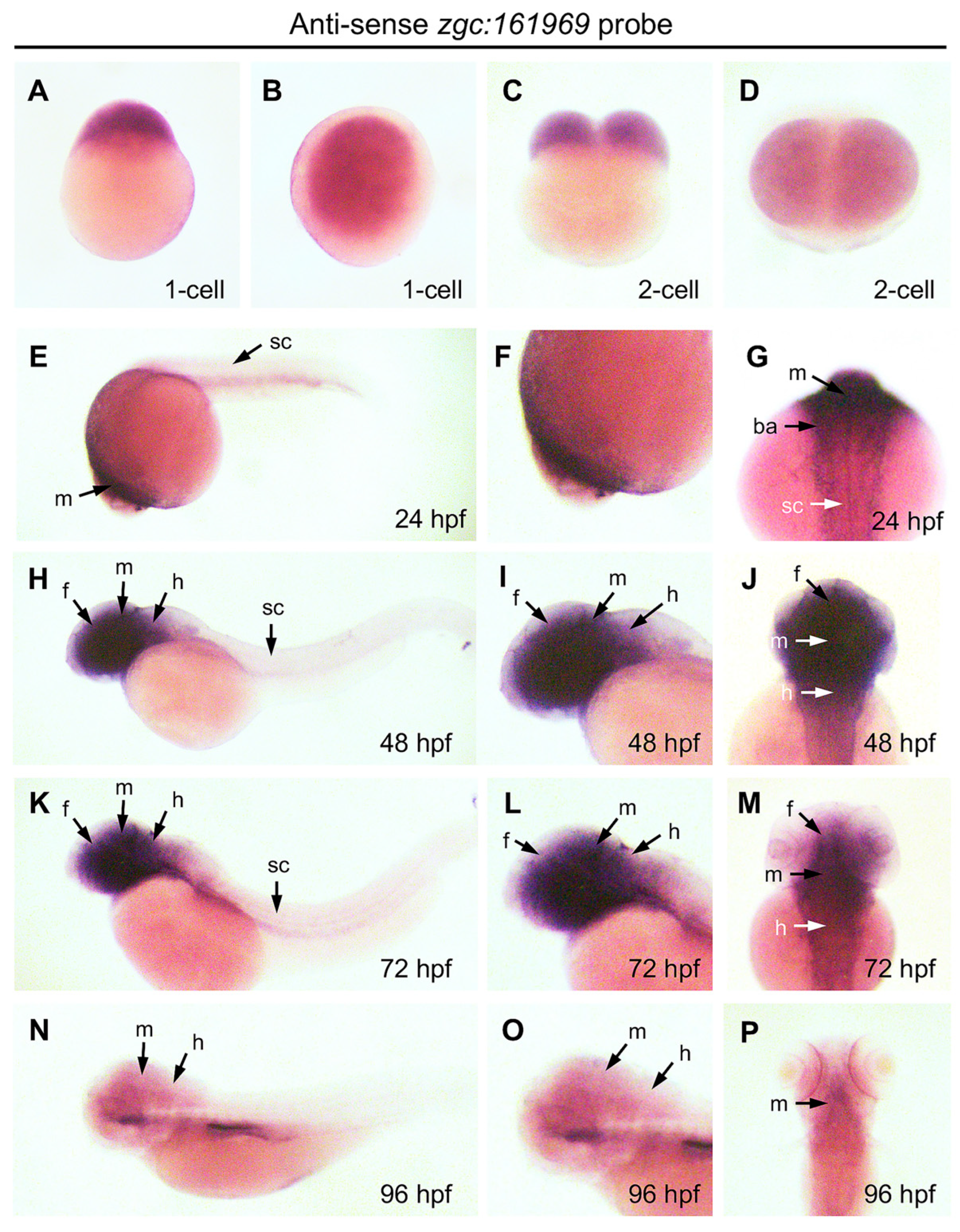
2.7. Whole-Mount In Situ Hybridization (WISH)
3. Results and Discussion
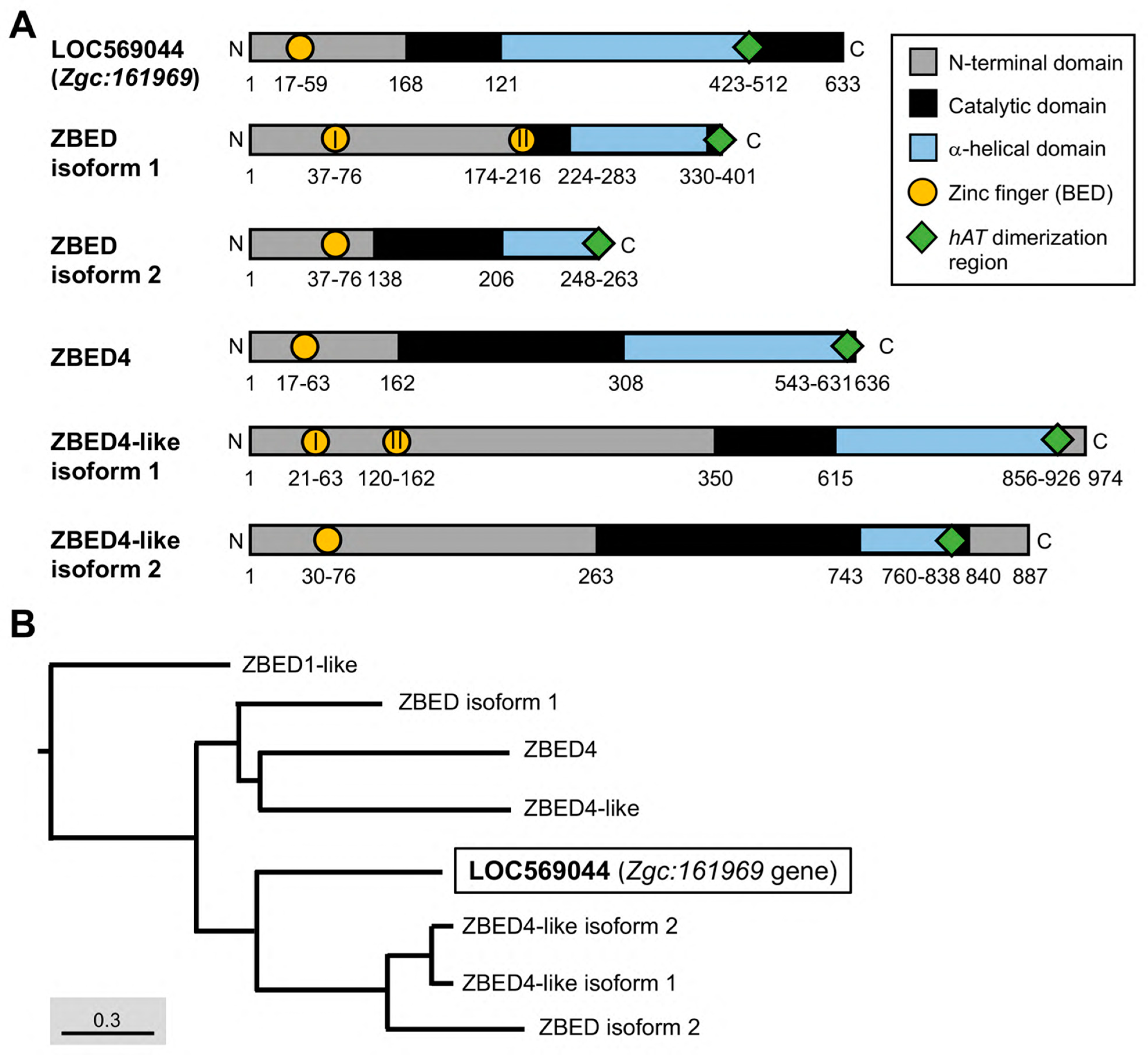
3.1. A New Member of the ZBED Protein Family Encoded by Zebrafish Zgc:161969
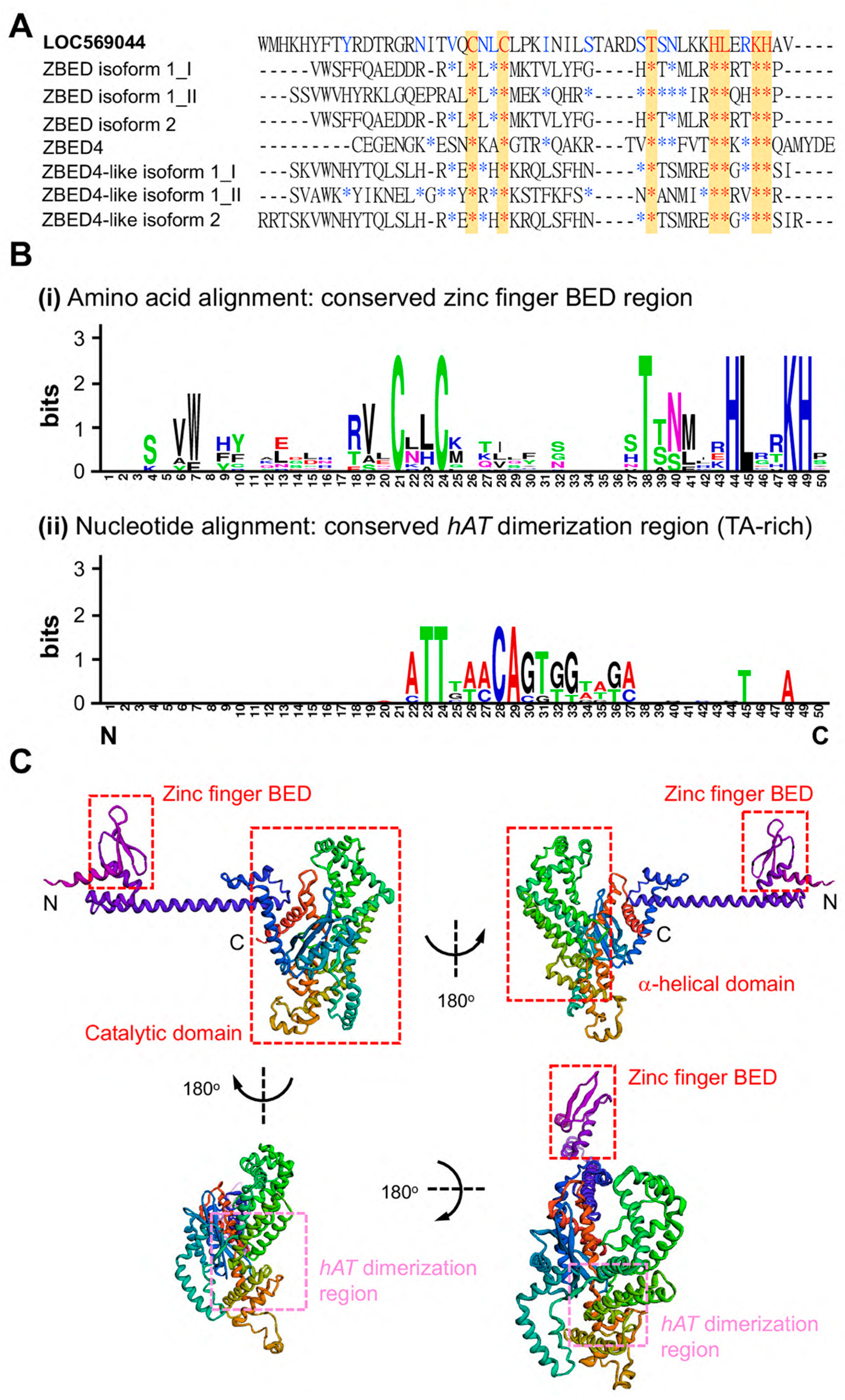
3.2. Prediction of ZBEDX Protein Structure
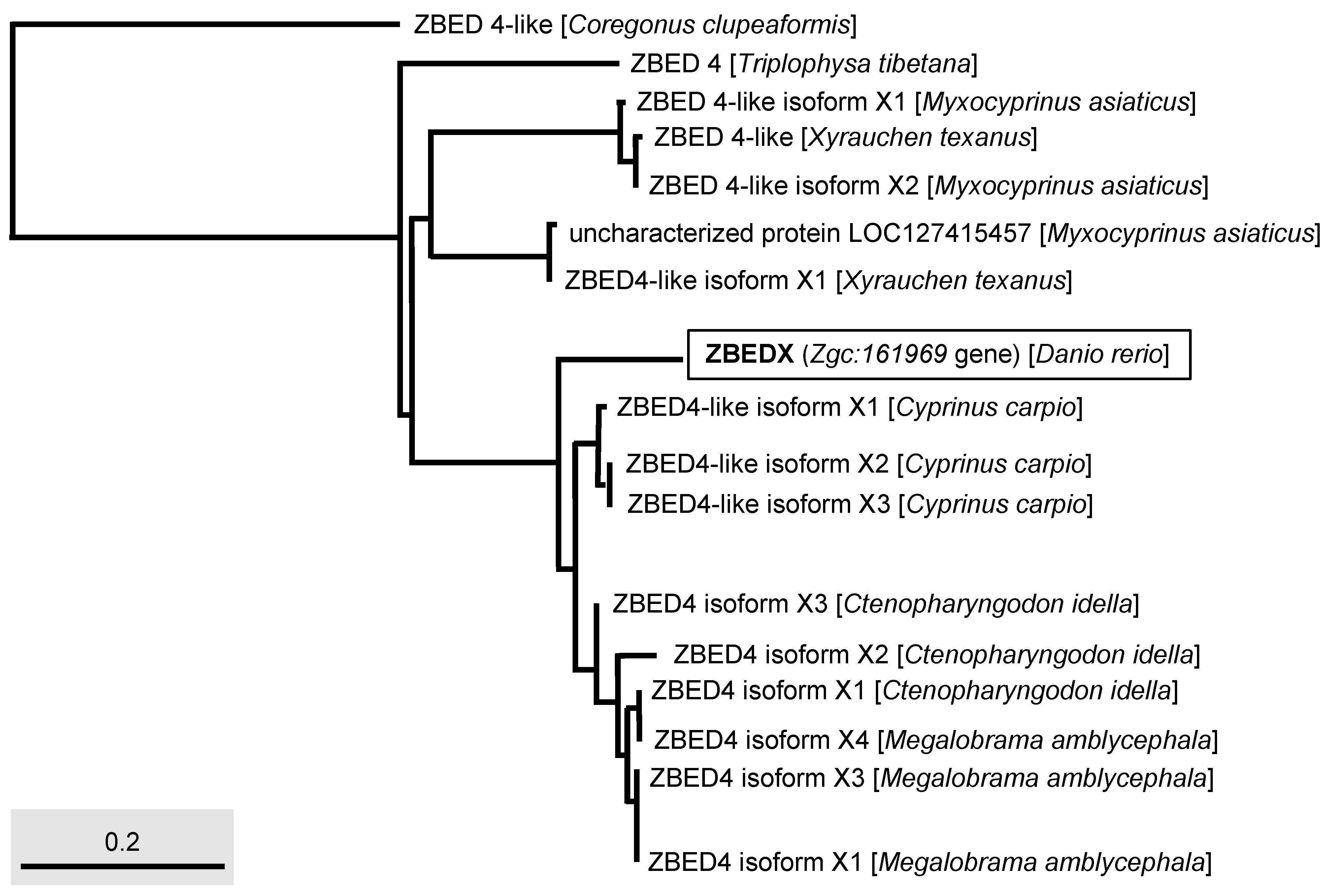
3.3. Phylogenetic Analysis of Zebrafish ZBEDX Protein
3.4. Temporospatial Expression of ZBEDX mRNA in Zebrafish Embryos
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sidik, H.; Talbot, W.S. A Zinc Finger Protein That Regulates Oligodendrocyte Specification, Migration, and Myelination in Zebrafish. Development 2015, 142, 4119–4128. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Wang, X.; Zhu, C.; Dong, X.; Zhang, Q.; Wang, X.; Duan, X.; Qian, F.; Shi, Y.; Gao, Y.; et al. Insm1a Regulates Motor Neuron Development in Zebrafish. Front. Mol. Neurosci. 2017, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- DeLaurier, A.; Eames, B.F.; Blanco-Sánchez, B.; Peng, G.; He, X.; Swartz, M.E.; Ullmann, B.; Westerfield, M.; Kimmel, C.B. Zebrafish Sp7:EGFP: A Transgenic for Studying Otic Vesicle Formation, Skeletogenesis, and Bone Regeneration. Genesis 2010, 48, 505–511. [Google Scholar] [CrossRef]
- Aravind, L. The BED Finger, a Novel DNA-Binding Domain in Chromatin-Boundary-Element-Binding Proteins and Transposases. Trends Biochem. Sci. 2000, 25, 421–423. [Google Scholar] [CrossRef]
- Somerville, T.D.D.; Xu, Y.; Wu, X.S.; Maia-Silva, D.; Hur, S.K.; de Almeida, L.M.N.; Preall, J.B.; Koo, P.K.; Vakoc, C.R. ZBED2 Is an Antagonist of Interferon Regulatory Factor 1 and Modifies Cell Identity in Pancreatic Cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 11471–11482. [Google Scholar] [CrossRef]
- Chen, M.; Philipp, M.; Wang, J.; Premont, R.T.; Garrison, T.R.; Caron, M.G.; Lefkowitz, R.J.; Chen, W. G Protein-Coupled Receptor Kinases Phosphorylate LRP6 in the Wnt Pathway. J. Biol. Chem. 2009, 284, 35040–35048. [Google Scholar] [CrossRef] [Green Version]
- Saghizadeh, M.; Akhmedov, N.B.; Yamashita, C.K.; Gribanova, Y.; Theendakara, V.; Mendoza, E.; Nelson, S.F.; Ljubimov, A.V.; Farber, D.B. ZBED4, a BED-Type Zinc-Finger Protein in the Cones of the Human Retina. Investig. Opthalmology Vis. Sci. 2009, 50, 3580. [Google Scholar] [CrossRef]
- Markljung, E.; Jiang, L.; Jaffe, J.D.; Mikkelsen, T.S.; Wallerman, O.; Larhammar, M.; Zhang, X.; Wang, L.; Saenz-Vash, V.; Gnirke, A.; et al. ZBED6, a Novel Transcription Factor Derived from a Domesticated DNA Transposon Regulates IGF2 Expression and Muscle Growth. PLoS Biol. 2009, 7, e1000256. [Google Scholar] [CrossRef]
- Zeng, C.-W.; Kamei, Y.; Shigenobu, S.; Sheu, J.-C.; Tsai, H.-J. Injury-Induced Cavl-Expressing Cells at Lesion Rostral Side Play Major Roles in Spinal Cord Regeneration. Open Biol. 2021, 11, 200304. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio); Zebrafish International Resource Center, the University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Zeng, C.-W.; Sheu, J.-C.; Tsai, H.-J. The Neuronal Regeneration of Adult Zebrafish after Spinal Cord Injury Is Enhanced by Transplanting Optimized Number of Neural Progenitor Cells. Cell Transplant. 2020, 29, 096368972090367. [Google Scholar] [CrossRef]
- Bateman, A. The Pfam Protein Families Database. Nucleic Acids Res. 2002, 30, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Zeng, C.-W.; Sheu, J.-C.; Tsai, H.-J. A New Member of the Forkhead Box Protein Family in Zebrafish: Domain Composition, Phylogenetic Implication and Embryonic Expression Pattern. Gene Expr. Patterns 2020, 35, 119093. [Google Scholar] [CrossRef]
- Kim, D.E.; Chivian, D.; Baker, D. Protein Structure Prediction and Analysis Using the Robetta Server. Nucleic Acids Res. 2004, 32, W526–W531. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.-W.; Sheu, J.-C.; Tsai, H.-J. Hypoxia-Responsive Subtype Cells Differentiate into Neurons in the Brain of Zebrafish Embryos Exposed to Hypoxic Stress. Cell Transplant. 2022, 31, 096368972210779. [Google Scholar] [CrossRef]
- Lin, C.-Y.; He, J.-Y.; Zeng, C.-W.; Loo, M.-R.; Chang, W.-Y.; Zhang, P.-H.; Tsai, H.-J. MicroRNA-206Modulates an Rtn4a/Cxcr4a/Thbs3a Axis in Newly Forming Somites to Maintain and Stabilize the Somite Boundary Formation of Zebrafish Embryos. Open Biol. 2017, 7, 170009. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.-W.; Kamei, Y.; Wang, C.-T.; Tsai, H.-J. Subtypes of Hypoxia-Responsive Cells Differentiate into Neurons in Spinal Cord of Zebrafish Embryos after Hypoxic Stress. Biol. Cell 2016, 108, 357–377. [Google Scholar] [CrossRef]
- Lee, H.-C.; Fu, C.-Y.; Zeng, C.-W.; Tsai, H.-J. Embryonic Expression Patterns of Eukaryotic EndoU Ribonuclease Family Gene EndouC in Zebrafish. Gene Expr. Patterns 2017, 25–26, 66–70. [Google Scholar] [CrossRef]
- O’brochta, D.A.; Atkinson, P.W. Transposable Elements and Gene Transformation in Non-Drosophilid Insects. Insect Biochem. Mol. Biol. 1996, 26, 739–753. [Google Scholar] [CrossRef]
- Kawakami, K.; Shima, A. Identification of the Tol2 Transposase of the Medaka Fish Oryzias Latipes That Catalyzes Excision of a Nonautonomous Tol2 Element in Zebrafish Danio Rerio. Gene 1999, 240, 239–244. [Google Scholar] [CrossRef]
- Arensburger, P.; Hice, R.H.; Zhou, L.; Smith, R.C.; Tom, A.C.; Wright, J.A.; Knapp, J.; O’Brochta, D.A.; Craig, N.L.; Atkinson, P.W. Phylogenetic and Functional Characterization of ThehATTransposon Superfamily. Genetics 2011, 188, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodard, L.E.; Li, X.; Malani, N.; Kaja, A.; Hice, R.H.; Atkinson, P.W.; Bushman, F.D.; Craig, N.L.; Wilson, M.H. Comparative Analysis of the Recently Discovered HAT Transposon TcBuster in Human Cells. PLoS ONE 2012, 7, e42666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, M.A.; Murray, A.; Samuels, H.H. NRC-Interacting Factor 1 Is a Novel Cotransducer That Interacts with and Regulates the Activity of the Nuclear Hormone Receptor Coactivator NRC. Mol. Cell. Biol. 2002, 22, 6883–6894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klug, A. The Discovery of Zinc Fingers and Their Development for Practical Applications in Gene Regulation and Genome Manipulation. Q. Rev. Biophys. 2010, 43, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc Finger Proteins: New Insights into Structural and Functional Diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Berg, J.M. Zinc Fingers and Other Metal-Binding Domains. Elements for Interactions between Macromolecules. J. Biol. Chem. 1990, 265, 6513–6516. [Google Scholar] [CrossRef]
- Hayward, A.; Ghazal, A.; Andersson, G.; Andersson, L.; Jern, P. ZBED Evolution: Repeated Utilization of DNA Transposons as Regulators of Diverse Host Functions. PLoS ONE 2013, 8, e59940. [Google Scholar] [CrossRef] [Green Version]
- Stünkel, W.; Kober, I.; Kauer, M.; Taimor, G.; Seifart, K.H. Human TFIIIA Alone Is Sufficient to Prevent Nucleosomal Repression of a Homologous 5S Gene. Nucleic Acids Res. 1995, 23, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural Stem Cells and Neurogenesis in the Adult Zebrafish Brain: Origin, Proliferation Dynamics, Migration and Cell Fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Name | Transcript ID | Length (aa) | Total Identity (%) a | BED Domain Identity (%) b |
|---|---|---|---|---|---|
| D. rerio | LOC569044 (Zgc:161969 gene) | BC134839.1 | 630 | 100 | 100 |
| ZBED1-like | XP_021327416.1 | 661 | 19 | 37 | |
| ZBED4 | XP_002666517.2 | 636 | 25 | 67 | |
| ZBED4-like | XP_001340716.5 | 650 | 20 | 58 | |
| ZBED4-like isoform 1 | XP_005165631.1 | 976 | 26 | 50 | |
| ZBED4-like isoform 2 | XP_005165631.1 | 887 | 26 | 50 | |
| ZBED isoform 1 | NP_001373713.1 | 401 | 34 | 52 | |
| ZBED isoform 2 | NP_001373714.1 | 263 | 26 | 54 |
| Species | Name | Transcript ID | Length (aa) | Identity (%) a |
|---|---|---|---|---|
| D. rerio | ZBEDX (Zgc:161969 gene) | NP_001077318.1 | 630 | 100 |
| C. auratus | ZBED1-like | XP_026072939.1 | 628 | 22 |
| ZBED1-like isoform X1 | XP_026117831.1 | 729 | 75 | |
| ZBED1-like isoform X2 | XP_026117841.1 | 667 | 75 | |
| ZBED4-like | XP_026116228.1 | 631 | 20 | |
| ZBED4-like isoform X1 | XP_026069790.1 | 722 | 75 | |
| ZBED4-like isoform X2 | XP_026069791.1 | 635 | 82 | |
| ZBED4-like isoform X3 | XP_026117848.1 | 642 | 81 | |
| H. sapiens | ZBED4 | AAI67155.1 | 1171 | 20 |
| E3 SUMO-protein ligase ZBED1 | NP_001164606.1 | 694 | 24 | |
| ZBED1 | AAH15030.1 | 694 | 21 | |
| M. musculus | ZBED4 | NP_852077.1 | 1168 | 21 |
| R. rattus | ZBED4 | XP_032775174.1 | 1170 | 21 |
| X. tropicalis | ZBED1 | XP_004911791.1 | 639 | 21 |
| ZBED1 isoform X1 | XP_012813067.2 | 693 | 20 |
| Species | Name | Transcript ID | Length (aa) | Identity (%) a |
|---|---|---|---|---|
| D. rerio | ZBEDX (Zgc:161969 gene) | NP_001077318.1 | 630 | 100 |
| Coregonus clupeaformis | ZBED4-like | XP_041752989.2 | 623 | 51 |
| T. tibetana | ZBED4 | KAA0713175.1 | 622 | 68 |
| Myxocyprinus asiaticus | ZBED 4-like isoform X1 | XP_051540434.1 | 637 | 69 |
| ZBED 4-like isoform X2 | XP_051540435.1 | 636 | 69 | |
| uncharacterized protein LOC127415457 | XP_051510176.1 | 627 | 67 | |
| Xyrauchen texanus | ZBED 4-like | XP_051973224.1 | 636 | 69 |
| ZBED4-like isoform X1 | XP_051950732.1 | 627 | 67 | |
| C. idella | ZBED4 isoform X1 | XP_051752038.1 | 749 | 78 |
| ZBED4 isoform X2 | XP_051752039.1 | 662 | 84 | |
| ZBED4 isoform X3 | XP_051752040.1 | 654 | 78 | |
| Megalobrama amblycephala | ZBED4 isoform X1 | XP_048056422.1 | 730 | 78 |
| ZBED4 isoform X3 | XP_048056424.1 | 724 | 78 | |
| ZBED4 isoform X4 | XP_048056425.1 | 716 | 78 | |
| C. carpio | ZBED4-like isoform X1 | XP_042613966.1 | 722 | 77 |
| ZBED4-like isoform X2 | XP_018955884.1 | 635 | 83 | |
| ZBED4-like isoform X3 | XP_018927958.2 | 637 | 83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, C.-W.; Sheu, J.-C.; Tsai, H.-J. Genomic Structure, Protein Character, Phylogenic Implication, and Embryonic Expression Pattern of a Zebrafish New Member of Zinc Finger BED-Type Gene Family. Genes 2023, 14, 179. https://doi.org/10.3390/genes14010179
Zeng C-W, Sheu J-C, Tsai H-J. Genomic Structure, Protein Character, Phylogenic Implication, and Embryonic Expression Pattern of a Zebrafish New Member of Zinc Finger BED-Type Gene Family. Genes. 2023; 14(1):179. https://doi.org/10.3390/genes14010179
Chicago/Turabian StyleZeng, Chih-Wei, Jin-Chuan Sheu, and Huai-Jen Tsai. 2023. "Genomic Structure, Protein Character, Phylogenic Implication, and Embryonic Expression Pattern of a Zebrafish New Member of Zinc Finger BED-Type Gene Family" Genes 14, no. 1: 179. https://doi.org/10.3390/genes14010179
APA StyleZeng, C. -W., Sheu, J. -C., & Tsai, H. -J. (2023). Genomic Structure, Protein Character, Phylogenic Implication, and Embryonic Expression Pattern of a Zebrafish New Member of Zinc Finger BED-Type Gene Family. Genes, 14(1), 179. https://doi.org/10.3390/genes14010179