
Genetic and Morphological Variation in Hypodontia of Maxillary Lateral Incisors

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Genotyping
2.3. Imaging and Morphometrics

{kind=link}
{kind=link}
{kind=link}
| Type of Measurement | Description |
|---|---|
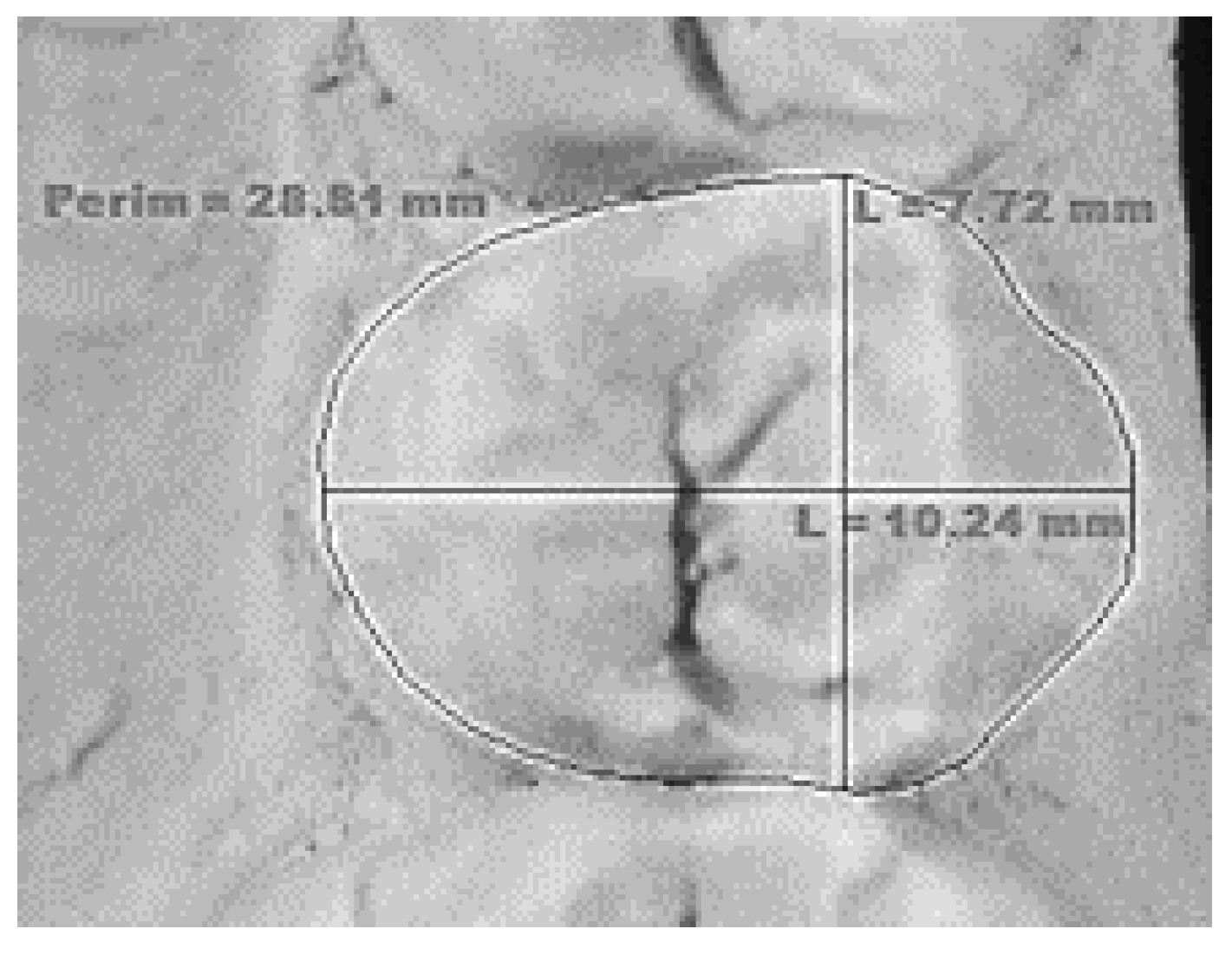
| Mesio-distal dimension | The greatest distance between the mesial and distal contact points (Figure 1). |
| Bucco-lingual dimension | The greatest distance of the tooth between the buccal and the lingual surface (Figure 1). |
| Occlusal area | Area of the occlusal surface defined by the mesial, buccal, distal, and lingual margins (Figure 1). |
| Intercuspal distance on upper first and second premolars | Distance between the tips of the buccal and lingual cusps. |
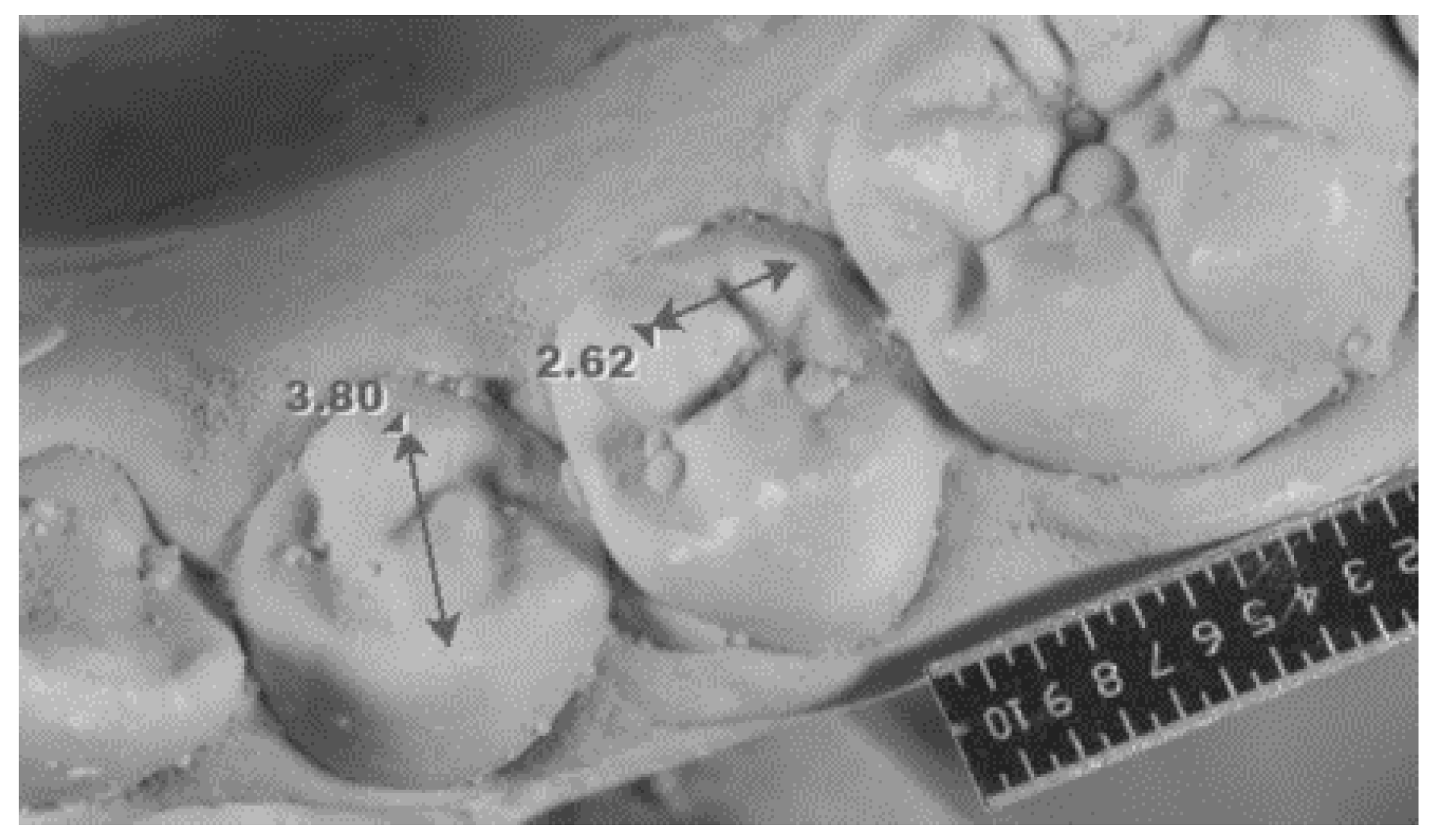
| Intercuspal distance on lower first premolars | Distance between the tips of the buccal and lingual cusps (Figure 2). |
| Intercuspal distance on lower second premolars | Distance between the tips of the two lingual cusps measured only in the case of the three-cusp version of the tooth (Figure 2). |
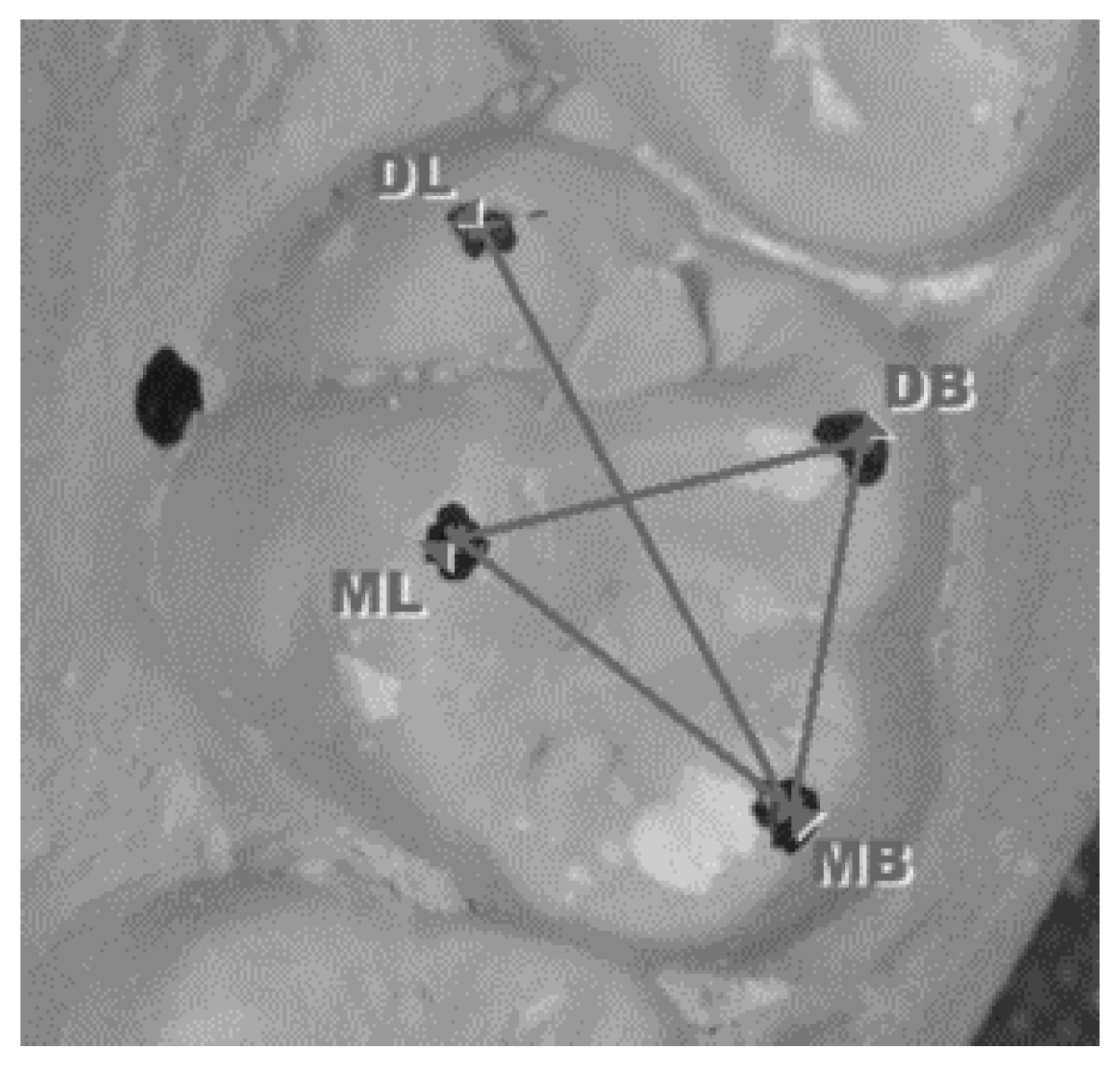
| Intercuspal distances on upper first molars | Distance between the tips of the cusps: mesiobuccal to distobuccal (MB-DB); distobuccal to mesiolingual (DB-ML); mesiolingual to mesiobuccal (ML-MB); and mesiobuccal to distolingual (MB-DL) (Figure 3). |
| Size of the cusp Carabelli on the upper first permanent molars | Diagnosed using the Arizona State University Dental Anthropology System [39] |
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
- (1)
- The genetic factor MSX1 rs8670 variant was associated with variable phenotypic outcomes in multiple morphological parameters of the formed teeth.
- (2)
- The new findings concerning compensatory interactions in the maxillary incisor region indicate interactions of epigenetic and environmental factors with the genetic variant.
- (3)
- A multiple model approach increases understanding of the aetiology of tooth agenesis. The addition of a single level, directional, complex interactive approach incorporates the findings of this study.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matalova, E.; Fleischmannova, J.; Sharpe, P.T.; Tucker, A.S. Tooth agenesis: From molecular genetics to molecular dentistry. J. Dent. Res. 2008, 87, 617–623. [Google Scholar] [CrossRef]
- Cobourne, M.T.; Sharpe, P.T. Diseases of the tooth: The genetic and molecular basis of inherited anomalies affecting the dentition. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Polder, B.J.; Van’t Hof, M.A.; Van Der Linden, F.P.G.M.; Kuijpers-Jagtman, A.M. A meta-analysis of the prevalence of dental agenesis of permanent teeth. Community Dent. Oral Epidemiol. 2004, 32, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Mattheeuws, N.; Dermaut, L.; Martens, G. Has hypodontia increased in Caucasians during the 20th century? A metaanalysis. Eur. J. Orthod. 2004, 26, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, A.H. A unifying aetiological explanation for anomalies of human tooth number and size. Arch. Oral. Biol. 1984, 29, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Brook, A.H.; Winter, G.B. Tooth Abnormalities. In Clinical Dentistry; Rowe, A.H.R., Johns, R.B., Eds.; Blackwell Scientific Publications: Oxford, UK, 1986; Volume 3, pp. 55–103. [Google Scholar]
- Primozic, J.; Farcnik, F.; Ovsenik, M. Places in the dental arch that show a greater variability in tooth number, shape and position. Arch. Oral. Biol. 2012, 57, 744–748. [Google Scholar] [CrossRef]
- Carter, K.; Worthington, S. Morphologic and Demographic Predictors of Third Molar Agenesis: A Systematic Review and Meta-analysis. J. Dent. Res. 2015, 94, 886–894. [Google Scholar] [CrossRef]
- Arte, S. Phenotypic and Genotypic Features of Familial Hypodontia. Ph.D. Thesis, Institute of Dentistry, University of Helsinki, Helsinki, Finland, 2001. [Google Scholar]
- Khalaf, K.; Miskelly, J.; Voge, E.; Macfarlane, T.V. Prevalence of hypodontia and associated factors: A systematic review and meta-analysis. J. Orthod. 2014, 41, 299–316. [Google Scholar] [CrossRef] [Green Version]
- Bassiouny, D.S.; Afify, A.R.; Baeshen, H.A.; Birkhed, D.; Zawawi, K.H. Prevalence of maxillary lateral incisor agenesis and associated skeletal characteristics in an orthodontic patient population. Acta Odontol. Scand. 2016, 74, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, T.; Abdullah, N.; Rsheadat, Y.; Hassan, M.I. Prevalence of isolated maxillary lateral incisor agenesis in Syrian adolescents. J. Orofac. Orthop. 2017, 78, 62–69. [Google Scholar] [CrossRef]
- Arandi, N.Z.; Mustafa, S. Maxillary lateral incisor agenesis; a retrospective cross-sectional study. Saudi Dent. J. 2018, 30, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Celikoglu, M.; Kamak, H.; Yildirim, H.; Ceylan, I. Investigation of the maxillary lateral incisor agenesis and associated dental anomalies in an orthodontic patient population. Med. Oral. Patol. Oral. Cir. Bucal. 2012, 17, e1068-73. [Google Scholar] [CrossRef]
- Tadros, M.; Brook, A.H.; Ranjitkar, S.; Townsend, G.C. Compensatory interactions between developing maxillary anterior teeth in a sample of twins. Arch. Oral. Biol. 2019, 97, 198–207. [Google Scholar] [CrossRef]
- Brook, A.H.; Elcock, C.; Al-Sharood, M.H.; McKeown, H.F.; Khalaf, K.; Smith, R.N. Further studies of a model for the etiology of anomalies of tooth number and size in humans. Connect Tissue Res. 2002, 43, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Hobkirk, J.A.; Gill, D.S.; Jones, S.P.; Hemmings, K.W.; Bassi, G.S.; O’Donnell, A.L.; Goodman, J.R. Hypodontia: A Team Approach to Management, 1st ed.; Wiley-Blackwell: Chichester, UK, 2011. [Google Scholar]
- Al-Ani, A.H. Genetic and Environmental Factors Associated with Hypodontia. Ph.D. Thesis, Clinical Dentistry, University of Otago, Dunedin, NewZealand, 2016. [Google Scholar]
- Pinho, T.; Maciel, P.; Lemos, C.; Sousa, A. Familial aggregation of maxillary lateral incisor agenesis. J. Dent. Res. 2010, 89, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Brook, A.H.; O’Donnell, M.B. The Dentition: A Complex System Demonstrating Self-Principles. In Proceedings of the Fifth IEEE International Conference on Self-Adaptive and Self-Organizing Systems (SASO), Ann Arbor, MI, USA, 3–7 October 2011; pp. 208–209. [Google Scholar]
- Al-Ani, A.H.; Antoun, J.S.; Thomson, W.M.; Merriman, T.R.; Farella, M. Hypodontia: An Update on Its Etiology, Classification, and Clinical Management. BioMed Res. Int. 2017, 2017, 9378325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, T.; Bockmann, M.; Mihalilidis, S.; Bennett, C.; Harris, A.; Seow, W.K.; Lekkas, D.; Ranjitkar, S.; Rupinskas, L.; Pinkerton, S.; et al. Genetic, epigenetic, and environmental influences on dentofacial structures and oral health: Ongoing studies of Australian twins and their families. Twin Res. Hum. Genet. 2013, 16, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.; Sassani, S.; Farella, M.; Ranjitkar, S.; Yong, R.; Swindells, S.; Brook, A. Variations in dental arch morphology are outcomes of the complex adaptive system associated with the developmental variation of hypodontia. Int. J. Des. Nat. Ecodyn. 2018, 13, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Ahmed, N.; Neela, P.K.; Unnisa, N. The Human Genetics of Dental Anomalies. Glob. Med. Genet. 2022, 9, 76–81. [Google Scholar] [CrossRef]
- Bianconi, G. Multilayer Networks; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Caldarelli, G.; Cantanzaro, M. Networks; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Barabasi, A.L. Network Science; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Galluccio, G.; Pilotto, A. Genetics of dental agenesis: Anterior and posterior area of the arch. Eur. Arch. Paediatr. Dent. 2008, 9, 41–45. [Google Scholar] [CrossRef]
- Alves-Ferreira, M.; Pinho, T.; Sousa, A.; Sequeiros, J.; Lemos, C.; Alonso, I. Identification of Genetic Risk Factors for Maxillary Lateral Incisor Agenesis. J. Dent. Res. 2014, 93, 452–458. [Google Scholar] [CrossRef]
- Park, K.; Kim, K.; Rho, S.B.; Choi, K.; Kim, D.; Oh, S.H.; Park, J.; Lee, S.H.; Lee, J.H. Homeobox MSX1 interacts with p53 tumor suppressor and inhibits tumor growth by inducing apoptosis. Cancer Res. 2005, 65, 749–757. [Google Scholar] [CrossRef]
- Boeira Junior, B.R.; Echeverrigaray, S. Polymorphism in the MSX1 gene in a family with upper lateral incisor agenesis. Arch. Oral. Biol. 2012, 57, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Brook, A.H.; Jernvall, J.; Smith, R.N.; Hughes, T.; Townsend, G.C. The dentition: The outcomes of morphogenesis leading to variations of tooth number, size and shape. Aust. Dent. J. 2014, 59, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Bosio, J.A.; Chou, J.C.; Jiang, S.S. Maxillary lateral incisor agenesis and its relationship to overall tooth size. J. Prosthet Dent. 2016, 115, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kerekes-Mathe, B.; Brook, A.H.; Martha, K.; Szekely, M.; Smith, R.N. Mild hypodontia is associated with smaller tooth dimensions and cusp numbers than in controls. Arch. Oral. Biol. 2015, 60, 1442–1449. [Google Scholar] [CrossRef]
- Kerekes-Máthé, B.; Mártha, K.; Brook O’Donnell, M.; Brook, A.H. Dental Arch Dimensions in a Matched Pairs Study of Hypodontia Patients and Controls. Biomed. J. Sci. Tech. Res. 2022, 41, 32426–32433. [Google Scholar]
- Sofaer, J.A.; Chung, C.S.; Niswander, J.D.; Runck, D.W. Developmental interaction, size and agenesis among permanent maxillary incisors. Hum. Biol. 1971, 43, 36–45. [Google Scholar] [PubMed]
- Kondo, S.; Hanamura, H. Does a maxillary lateral incisor reduce to compensate for a large central incisor? Aichi Gakuin J. Dent. Sci. 2010, 48, 215–227. [Google Scholar]
- Brook, A.H.; Smith, R.N.; Elcock, C.; Al-Sharood, M.H.; Shah, A.A.; Khalaf, K.; Robinson, D.L.; Lath, D.L.; Karmo, M. The Measurement of tooth morphology: Validation of an image analysis system. In Current Trends in Dental Morphology Research; Zadzinska, E., Ed.; University of Lodz Press: Łódź, Poland, 2005; pp. 475–482. [Google Scholar]
- Scott, G.R.; Irish, J.D. Human Tooth Crown and Root Morphology: The Arizona State University Dental Anthropology System; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Trybek, G.; Jaroń, A.; Grzywacz, A. Association of Polymorphic and Haplotype Variants of the MSX1 Gene and the Impacted Teeth Phenomenon. Genes 2021, 12, 577. [Google Scholar] [CrossRef]
- Liang, J.; Von den Hoff, J.; Lange, J.; Ren, Y.; Bian, Z.; Carels, C.E. MSX1 mutations and associated disease phenotypes: Genotype-phenotype relations. Eur. J. Hum. Genet. 2016, 24, 1663–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, C. What Are 3′ UTRs Doing? Cold Spring Harb. Perspect Biol. 2019, 11, a034728. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Letra, A. The changing landscape in the genetic aetiology of human tooth agenesis. Genes 2018, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wong, S.W.; Han, D.; Cai, T. Genetic analysis: Wnt and other pathways in non-syndromic tooth agenesis. Oral. Dis. 2019, 25, 646–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mártha, K.; Kerekes Máthé, B.; Moldovan, V.G.; Bănescu, C. Study of rs12532, rs8670 Polymorphism of Msh Homeobox 1 (MSX1), rs61754301, rs4904155 Polymorphism of Paired Box Gene 9 (PAX9), and rs2240308 Polymorphism of Axis Inhibitor Protein 2 (AXIN2) Genes in Nonsyndromic Hypodontia. BioMed Res. Int. 2019, 2019, 2183720. [Google Scholar] [CrossRef]
- Kieser, J.A.; Groeneveld, H.T.; Preston, C.B. On the non-existence of compensatory tooth size interaction in a contemporary human population. J. Dent. Res. 1986, 65, 1105–1107. [Google Scholar] [CrossRef]
- Dhamo, B.; Nguee, A.M.; Ongkosuwito, E.M.; Jaddoe, V.W.V.; Wolvius, E.B.; Kragt, L. The role of accelerated dental development on the occurrence of aberrant dental traits that indicate malocclusion. Eur. J. Orthod 2019, 41, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, A.A. The changing dentition of man. J. Am. Dent. Assoc. 1945, 32, 676–690. [Google Scholar] [CrossRef]
- Townsend, G.; Harris, E.F.; Lesot, H.; Clauss, F.; Brook, A.H. Morphogenetic fields within the human dentition: A new, clinically relevant synthesis of an old concept. Arch. Oral. Biol. 2009, 54, S34–S44. [Google Scholar] [CrossRef]
- Brook, A.H.; Elcock, C.; Aggarwal, M.; Lath, D.L.; Russell, J.M.; Patel, P.I.; Smith, R.N. Tooth dimensions in hypodontia with a known PAX9 mutation. Arch. Oral. Biol. 2009, 54, S57–S62. [Google Scholar] [CrossRef] [Green Version]
- Sato, A.; Arai, K. Sexual dimorphism of tooth agenesis patterns in non-syndromic oligodontia in Japanese patients. Arch. Oral. Biol. 2021, 124, 105038. [Google Scholar] [CrossRef] [PubMed]
- Alamoudi, R.; Ghamri, M.; Mistakidis, I.; Gkantidis, N. Sexual Dimorphism in Third Molar Agenesis in Humans with and without Agenesis of Other Teeth. Biology 2022, 11, 1725. [Google Scholar] [CrossRef]
- Heuberer, S.; Ulm, C.; Zechner, W.; Laky, B.; Watzak, G. Patterns of congenitally missing teeth of non-syndromic and syndromic patients treated at a single-center over the past thirty years. Arch. Oral. Biol. 2019, 98, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Butler, P.M. Studies of the mammalian dentition–differentiation of the post-canine dentition. Proc. Zool. Soc. Lond. Ser. B 1939, 109, 1–36. [Google Scholar] [CrossRef]
- Mitsiadis, T.A.; Smith, M.M. How do teeth make genes to order through development? J. Exp. Biol. Part B Mol. Dev. Evol. 2006, 306, 177–182. [Google Scholar] [CrossRef]
- Townsend, G.; Hughes, T.; Luciano, M.; Bockmann, M.; Brook, A.H. Genetic and environmental influences on human dental variation. Arch. Oral. Biol. 2009, 54S, 45–51. [Google Scholar] [CrossRef]
- Sharpe, P.J. Homeobox genes and orofacial development. Connect. Tissue Res. 1995, 32, 17–25. [Google Scholar] [CrossRef]
- Osborn, J.W. Morphogenetic gradients: Fields versus clones. In Development, Function and Evolution of Teeth; Butler, P.M., Joysey, K.A., Eds.; Academic Press: London, UK, 1978; pp. 171–201. [Google Scholar]
- Jernvall, J. A gene network model accounting for development and evolution of mammalian teeth. Proc. Natl. Acad. Sci. USA 2002, 99, 8116–8120. [Google Scholar]
- Salazar-Ciudad, I.; Jernvall, J. A computational model of teeth and the developmental origins of morphological variation. Nature 2010, 464, 583–586. [Google Scholar] [CrossRef]



| MSX1 rs8670 C/T | Upper Lateral Incisor Agenesis (N = 26) n | Matched Controls (N = 26) n | Statistical Analysis for Genotypes | Statistical Analysis for Alleles |
|---|---|---|---|---|
| C/C | 12 | 22 | C/T, T/T vs. C/C p = 0.007 OR = 6.41 | T vs. C p = 0.0003 OR = 6.90 |
| C/T | 9 | 4 | ||
| T/T | 5 | 0 | ||
| C | 33 | 48 | ||
| T | 19 | 4 |
| Teeth | MD | BL | OA | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hypodontia | Control | p | Hypodontia | Control | p | Hypodontia | Control | p | |
| 1.1; 2.1 | 8.63 ± 0.53 | 8.87 ± 0.6 | 0.06 | 6.65 ± 0.74 | 7.45 ± 0.62 | 0.01 | 44.63 ± 6.37 | 51.51 ± 8.37 | 0.03 |
| 1.3; 2.3 | 7.78 ± 0.59 | 7.82 ± 0.47 | 0.81 | 7.97 ± 0.63 | 8.13 ± 0.26 | 0.43 | 44.76 ± 5.35 | 46.99 ± 4.12 | 0.33 |
| 1.4; 2.4 | 6.99 ± 0.54 | 7.27 ± 0.48 | 0.009 | 9.07 ± 0.39 | 9.61 ± 0.68 | 0.01 | 50.56 ± 3.98 | 55.88 ± 7.83 | 0.03 |
| 1.5; 2.5 | 6.70 ± 0.37 | 6.82 ± 0.48 | 0.42 | 9.04 ± 0.62 | 9.74 ± 0.60 | 0.01 | 47.53 ± 5.89 | 53.17 ± 8.18 | 0.10 |
| 1.6; 2.6 | 10.36 ± 0.56 | 10.48 ± 0.54 | 0.42 | 11.00 ± 0.27 | 11.97 ± 0.61 | 0.81 | 99.49 ± 10.26 | 106.20 ± 11.86 | 0.15 |
| 3.1; 4.1 | 5.40 ± 0.4 | 5.67 ± 0.44 | 0.03 | 5.77 ± 0.40 | 6.27 ± 0.41 | 0.01 | 22.01 ± 1.49 | 26.00 ± 3.80 | 0.004 |
| 3.2; 4.2 | 5.83 ± 0.45 | 6.12 ± 0.39 | 0.02 | 6.10 ± 0.57 | 6.52 ± 0.43 | 0.08 | 24.94 ± 2.13 | 28.90 ± 3.62 | 0.005 |
| 3.3; 4.3 | 6.77 ± 0.48 | 6.88 ± 0.46 | 0.43 | 6.89 ± 0.90 | 7.62 ± 0.36 | 0.03 | 33.99 ± 4.67 | 38.10 ± 3.68 | 0.03 |
| 3.4; 4.4 | 7.17 ± 0.34 | 7.10 ± 0.47 | 0.78 | 8.02 ± 0.57 | 8.05 ± 0.53 | 0.86 | 42.57 ± 3.31 | 45.37 ± 5.68 | 0.11 |
| 3.5; 4.5 | 7.03 ± 0.45 | 7.31 ± 0.45 | 0.04 | 8.36 ± 0.54 | 8.75 ± 0.57 | 0.04 | 45.55 ± 5.13 | 50.46 ± 5.87 | 0.03 |
| 3.6; 4.6 | 11.05 ± 0.75 | 11.20 ± 0.67 | 0.45 | 10.50 ± 0.43 | 11.13 ± 0.65 | 0.01 | 96.72 ± 8.42 | 105.05 ± 10.60 | 0.06 |
| MD | ||||||
| Teeth | 1.1; 2.1 | 1.2 or 2.2 | ||||
| Hypodontia | Control | p | Hypodontia | Control | p | |
| A | 8.93 ± 0.65 | 8.86 ± 0.57 | 0.69 | 6.27 ± 0.69 | 6.96 ± 0.58 | 0.02 |
| B | 8.79 ± 0.45 | 0.68 | 4.82 ± 0.54 | <0.0001 | ||
| C | 8.86 ± 0.53 | 0.98 | 5.59 ± 0.96 | 0.0001 | ||
| D | 8.35 ± 0.31 | 0.01 | NA | NA | ||
| BL | ||||||
| Teeth | 1.1; 2.1 | 1.2 or 2.2 | ||||
| Hypodontia | Control | p | Hypodontia | Control | p | |
| A | 6.84 ± 0.53 | 7.45 ± 0.62 | 0.11 | 6.37 ± 0.87 | 6.47 ± 0.68 | 0.82 |
| B | 6.50 ± 0.87 | 0.04 | 4.72 ± 0.49 | <0.001 | ||
| C | 6.69 ± 0.67 | 0.02 | 5.60 ± 1.10 | 0.08 | ||
| D | 6.58 ± 0.94 | 0.03 | NA | NA | ||
| OA | ||||||
| Teeth | 1.1; 2.1 | 1.2 or 2.2 | ||||
| Hypodontia | Control | p | Hypodontia | Control | p | |
| A | 46.98 ± 7.67 | 51.51 ± 8.37 | 0.42 | 32.02 ± 7.58 | 36.02 ± 2.62 | 0.41 |
| B | 43.48 ± 5.51 | 0.09 | 18.17 ± 2.82 | 0.001 | ||
| C | 45.42 ± 6.63 | 0.12 | 25.56 ± 9.13 | 0.01 | ||
| D | 43.20 ± 6.33 | 0.12 | NA | NA | ||
| Intercuspal Distance | Upper Lateral Incisor Agenesis Group | Matched Controls | p |
|---|---|---|---|
| Upper first premolars | 5.39 ± 0.59 | 5.88 ± 0.46 | 0.03 |
| Upper second premolars | 5.60 ± 0.39 | 6.04 ± 0.41 | 0.0001 |
| Lower first premolars | 3.63 ± 0.61 | 4.02 ± 0.53 | 0.01 |
| Lower second premolars (distance between lingual cusps) | 2.78 ± 0.39 | 3.54 ± 0.31 | <0.00001 |
| Upper first molars | |||
| MB-DB | 5.08 ± 0.49 | 5.04 ± 0.44 | 0.38 |
| DB-ML | 6.92 ± 0.77 | 6.73 ± 0.42 | 0.14 |
| ML-MB | 6.49 ± 0.53 | 6.57 ± 0.56 | 0.31 |
| MB-DL | 8.96 ± 0.47 | 9.24 ± 0.74 | 0.07 |
| Teeth | 1.2; 2.1 | 1.4; 2.4 | 3.2; 4.2 | 3.5; 4.5 | 3.6; 4.6 |
|---|---|---|---|---|---|
| Unilateral agenesis | 8.86 ± 0.53 | 7.20 ± 0.58 | 6.09 ± 0.39 | 7.21 ± 0.51 | 11.36 ± 0.78 |
| Bilateral agenesis | 8.35 ± 0.31 | 6.74 ± 0.32 | 5.51 ± 0.25 | 6.83 ± 0.27 | 10.68 ± 0.48 |
| p value | 0.01 | 0.01 | 0.0003 | 0.03 | 0.01 |
| Outcome Variables | Effect of the Gene Factor (T Allele of the rs8670 in the MSX1 Gene) (N = 14) | Effect of the Hypodontia Factor (Unilateral/Bilateral Hypodontia/Control) | ||
|---|---|---|---|---|
| p value | F ratio | p value | F ratio | |
| Mesio-distal diameter of upper central incisors | 0.04 | 7.39 | 0.29 | 1.57 |
| Bucco-lingual diameter of upper central incisors | 0.05 | 6.61 | 0.34 | 1.31 |
| Occlusal area of upper central incisors | 0.01 | 13.03 | 0.93 | 0.03 |
| Intercuspal distance of lower second premolars | 0.67 | 0.20 | 0.01 | 11.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerekes-Máthé, B.; Mártha, K.; Bănescu, C.; O’Donnell, M.B.; Brook, A.H. Genetic and Morphological Variation in Hypodontia of Maxillary Lateral Incisors. Genes 2023, 14, 231. https://doi.org/10.3390/genes14010231
Kerekes-Máthé B, Mártha K, Bănescu C, O’Donnell MB, Brook AH. Genetic and Morphological Variation in Hypodontia of Maxillary Lateral Incisors. Genes. 2023; 14(1):231. https://doi.org/10.3390/genes14010231
Chicago/Turabian StyleKerekes-Máthé, Bernadette, Krisztina Mártha, Claudia Bănescu, Matthew Brook O’Donnell, and Alan H. Brook. 2023. "Genetic and Morphological Variation in Hypodontia of Maxillary Lateral Incisors" Genes 14, no. 1: 231. https://doi.org/10.3390/genes14010231
APA StyleKerekes-Máthé, B., Mártha, K., Bănescu, C., O’Donnell, M. B., & Brook, A. H. (2023). Genetic and Morphological Variation in Hypodontia of Maxillary Lateral Incisors. Genes, 14(1), 231. https://doi.org/10.3390/genes14010231